


Research Progress in Biological Characteristics of Fructilactobacillus sanfranciscensis and Its Influence on Fermented Pasta
-
摘要: 旧金山果实乳杆菌(Fructilactobacillus sanfranciscensis)作为酸面团中常见的菌种,是具有良好应用价值的酸面团乳酸菌之一,该菌种通过自身的生长代谢及其与酸面团中酵母菌间的相互作用对发酵面食的风味、营养及质构等方面产生重要影响。自1971年分离该菌株以来,其分类学地位经历多次变迁,命名从Lactobacillus sanfrancisco变更为Lactobacillus sanfranciscensis,在2020年确定为Fructilactobacillus sanfranciscensis,至今,该菌种相关的研究文献已达500余篇。目前,该菌种已列入欧洲食品安全局的可食用菌种名单和韩国的限制性使用的微生物菌种名单,但尚不在我国可食用菌种名单中。本文系统综述了旧金山果实乳杆菌目前的分类学地位及其在酸面团中的普遍性,菌种的生化特性及基因特性,并阐述了其对发酵面食的影响,探索旧金山果实乳杆菌在酸面团中的应用潜力,以期开发用于酸面团的标准化发酵剂,为传统发酵面食产业化提供理论基础。
-
关键词:
- 酸面团 /
- 旧金山果实乳杆菌 /
- 发酵面食 /
- 酿酒酵母 /
- 矮小哈萨克斯坦酵母菌
Abstract: Fructilactobacillus sanfranciscensis, a common used strain in sourdough, is considered as one of the most valuable lactic acid bacteria (LAB) in the sourdough. F. sanfranciscensis has an important impact on the flavor, nutrition, and texture of fermented pasta through its own growth and metabolism as well as its interactions with yeasts in sourdough. Since this strain was isolated in 1971, its taxonomic status has undergone several changes, with the nomenclature changing from Lactobacillus sanfrancisco to Lactobacillus sanfranciscensis, and in 2020 it was identified as Fructilactobacillus sanfranciscensis, and to date, more than 500 research papers related to this strain have been published. Currently, this strain is included in the list of food cultures of the European Food Safety Authority (EFSA) and the list of microbial strains for restricted use in Korea, but it is not yet in the list of microorganisms used in food in China. This paper provides a systematical overview of the current taxonomic status of F. sanfranciscensis as well as its prevalence in sourdough production. Currently, it explores the biochemical and genetic characteristics of this species and their effects on fermented pasta. This review aims to explore the potential of the application of F. sanfranciscensis in sourdoughs for developing standardized sourdough fermentation agents and providing a theoretical basis for the industrialization of traditional fermented pasta. -
酸面团是制作馒头、面包等发酵面食的传统发酵剂,具有悠久的使用历史。在我国,酸面团发酵制备馒头、包子及发面饼等面食,传统酸面团发酵技术可追溯至2000年前的东汉时期[1];在国外,考古证据表明,酸面团发酵面包可追溯到5000年前的古埃及[2]。如今,酸面团广泛用于工业和手工馒头及面包店的各种产品中,从馒头、包子、馕到面包、饼干、法式长棍面包、煎饼和比萨饼等发酵面食。酸面团是由谷物粉和水的混合物经过天然或发酵剂发酵的产物,属于多菌种发酵体系,目前已在酸面团中检测到80余种乳酸菌,包括旧金山果实乳杆菌(Fructilactobacillus sanfranciscensis,F. sanfranciscensis)、植物乳植杆菌(Lactiplantibacillus plantarum,L. plantarum)和戊糖片球菌(Pediococcus pertosaceus,P. pertosaceus)等,以及80种酵母菌,包括酿酒酵母菌(Saccharomyces cerevisiae,S. cerevisiae)、矮小哈萨克斯坦酵母菌(Kazachstania humilis,K. humilis)和库德里阿兹威毕赤酵母(Pichia kudriavzevii,P. kudriavzevii)等[3−5]。其中,F. sanfranciscensis是酸面团生态系统中最普遍和最关键的乳酸菌之一,被视为用于发酵酸面团的关键微生物,受到国内外的广泛关注,该菌种发酵赋予了面包、馒头等发酵面食独特的口感和风味[4,6−7]。
F. sanfranciscensis作为发酵面食中重要的微生物菌种之一,在食品工业中扮演着关键角色,近年来,其安全性和应用潜力受到了国际社会的广泛关注。欧洲食品安全局(European Food Safety Authority,EFSA)在2016年将F. sanfranciscensis列入可食用菌种名单[8]。在韩国,F. sanfranciscensis可作为食品中限制性使用的微生物菌种,可用于面包制作[9]。目前,我国《可用于食品的菌种名单》明确列出38个可食用菌种,而F. sanfranciscensis未被列入名单中。2022年,中国工业微生物菌种保藏管理中心发表了传统发酵食品用微生物菌种名单,该名单涵盖56个属124种[10],在这些微生物中,仅有3种被明确指出可应用于发酵面食的生产(面包和馒头),包括S. cerevisiae、L. plantarum、干酪乳杆菌,而干酪乳杆菌明确可用于发酵制备面包,而非馒头中。未来,随着对F. sanfranciscensis更深入的研究,积极推动F. sanfranciscensis列入我国食品用微生物菌种名单,为推动F. sanfranciscensis在发酵面食产业中的生产和应用奠定理论基础。
本文从F. sanfranciscensis的分类学地位变迁开始梳理,明确菌种的生态位分布,介绍F. sanfranciscensis的生化代谢特性,阐明F. sanfranciscensis在基因水平上的基本特征,阐述F. sanfranciscensis的菌种特性及其对发酵面食的影响,为进一步应用F. sanfranciscensis微生物资源提供理论参考。
1. 旧金山果实乳杆菌的研究现状
1.1 旧金山果实乳杆菌在酸面团中的普遍性
F. sanfranciscensis在中国、意大利、比利时、法国、德国、美国等国家的酸面团样本中均被鉴定出,该菌种普遍存在于I型酸面团中,是常用于发酵面包和馒头等发酵面食的乳酸菌之一[5,11]。鉴于F. sanfranciscensis独特的生理和基因特性,菌株的来源受到研究者们的关注,早期研究普遍认为该菌种是只从酸面团环境中分离出的自生菌,但随着宏基因组测序技术的进步,已有研究表明在昆虫幼虫体内检测到F. sanfranciscensis[12−13]。
1.2 旧金山果实乳杆菌的分类学地位变迁
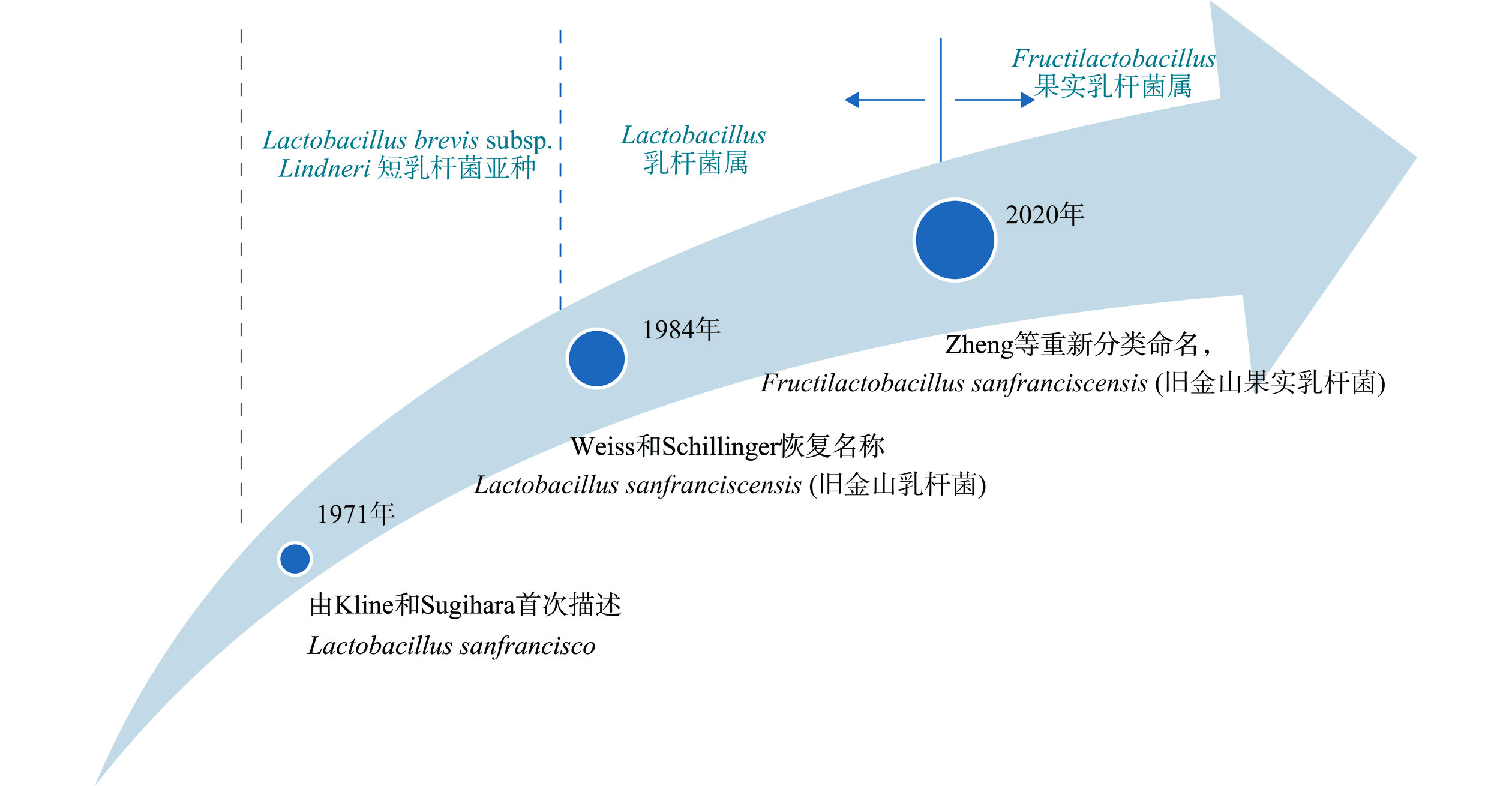
传统的乳酸菌分类主要以形态特征和生理生化特征(包括最佳生长温度、糖的利用和产生的代谢产物)为依据,费时费力且不精确,而正确的微生物系统分类学信息是提出和利用菌种名单的基本前提,分子微生物学技术的进步则为确定F. sanfranciscensis的分类学地位提供了依据,图1展示了F. sanfranciscensis的分类学地位变迁。在1971年,Kline和Sugihara从旧金山酸面团中分离并首次以Lactobacillus sanfrancisco描述了该专性异型发酵乳酸菌[14],Lactobacillus sanfrancisco这个名字是取自酸面团的来源城市:旧金山[15]。因从酸面团中分离的“L. brevis var. lindneri”菌株的DNA与F. sanfranciscensis DSM20451具有97%以上的DNA相似性,以及同类型的Lys-DAla细胞壁蛋白[16],因此,在1978年之前F. sanfranciscensis被指定为Lactobacillus brevis subsp. lindneri短乳杆菌亚种[17]。但在当时,该物种未被列入批准的细菌名称清单,并且在细菌学命名中没有地位,命名问题也可能是分类混乱的原因,直到1984年Weiss和Schillinger恢复该名称[18]。为了遵循《国际原核生物命名法规》(the International Code of Nomenclature of Prokaryotes)同位主位名词的要求,于1997年将该菌株名称由Lactobacillus sanfrancisco改为Lactobacillus sanfranciscensis[19],对应的中文名为“旧金山乳杆菌”。
![]() 图 1 旧金山果实乳杆菌的分类学地位变迁Figure 1. Evolution of the taxonomic status of F. sanfranciscensis
图 1 旧金山果实乳杆菌的分类学地位变迁Figure 1. Evolution of the taxonomic status of F. sanfranciscensis因该乳酸菌不以果糖作为碳源,而是将果糖作为电子受体的生长偏好性,在2020年4月,Zheng等[20]在《国际系统与进化微生物学杂志》(International Journal of Systematic and Evolutionary Microbiology, IJSEM)发表了乳酸菌的重要分类学变迁研究,对乳杆菌属(Lactobacillus)的261个菌种的全基因组和生理生化数据进行分析,将乳杆菌属重新分类为25个,IJSEM期刊发表的菌种新名称是被广泛认可的,也是国际权威机构或组织更新相应菌种名单的重要依据,其中,Lactobacillus sanfranciscensis更名为Fructilactobacillus sanfranciscensis,分属乳杆菌科Lactobacillaceae,从乳杆菌属Lactobacillus变到果实乳杆菌属Fructobacillus,需注意的是在2020年Zheng等研究乳酸菌分属问题之前,果乳杆菌属是属于明串珠菌科Leuconostocaceae。同年,为规范微生物学家对微生物名称的使用,中国工业微生物菌种保藏管理中心(China Center of Industrial Culture Collection,CICC)汇编了乳酸菌中分类学地位发生变化的新物种的中文名称和描述,Fructilactobacillus sanfranciscensis的中文名定为:旧金山果实乳杆菌[21]。
1.3 旧金山果实乳杆菌的研究概况
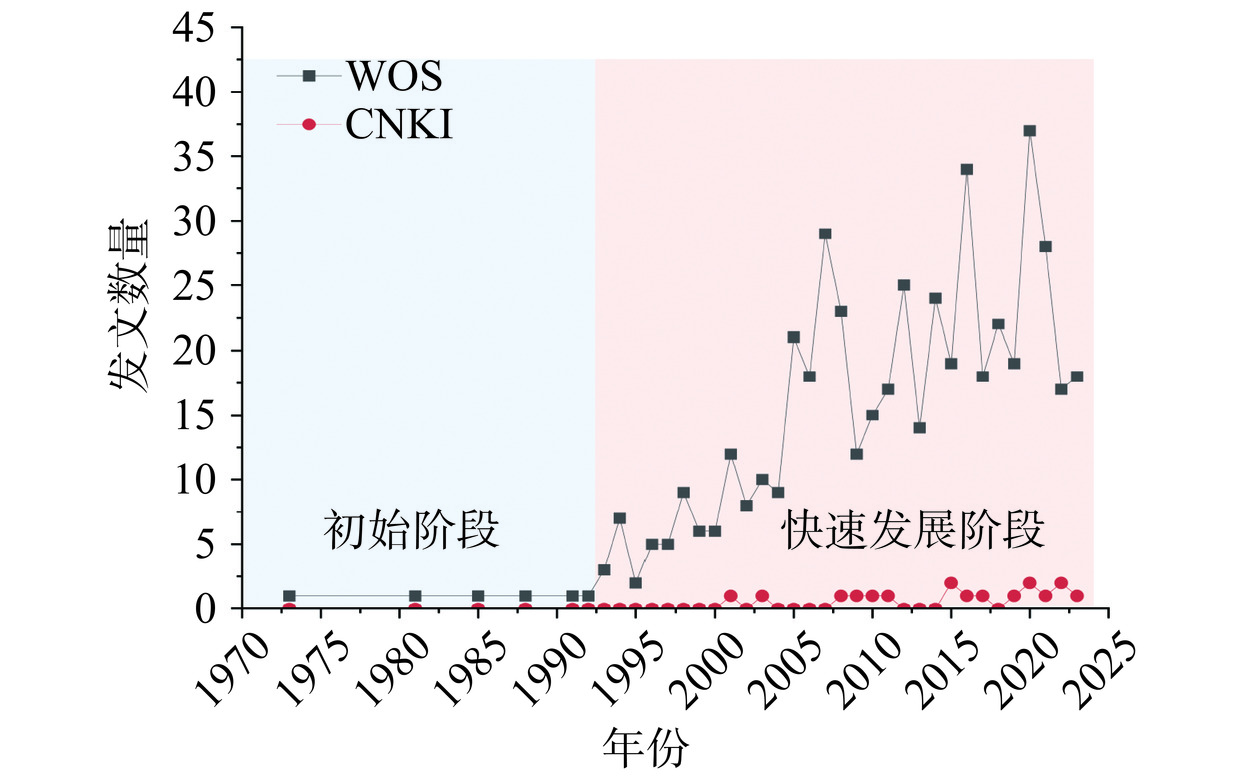
从1971年开始至今,在Web of Science(WOS)数据库和中国知网(CNKI)数据库有关F. sanfranciscensis的学术论文发表数量如图2所示。
![]() 图 2 1971-2024年旧金山果实乳杆菌学术论文发表数量Figure 2. Number of academic literature on F. sanfranciscensis from 1971 to 2024
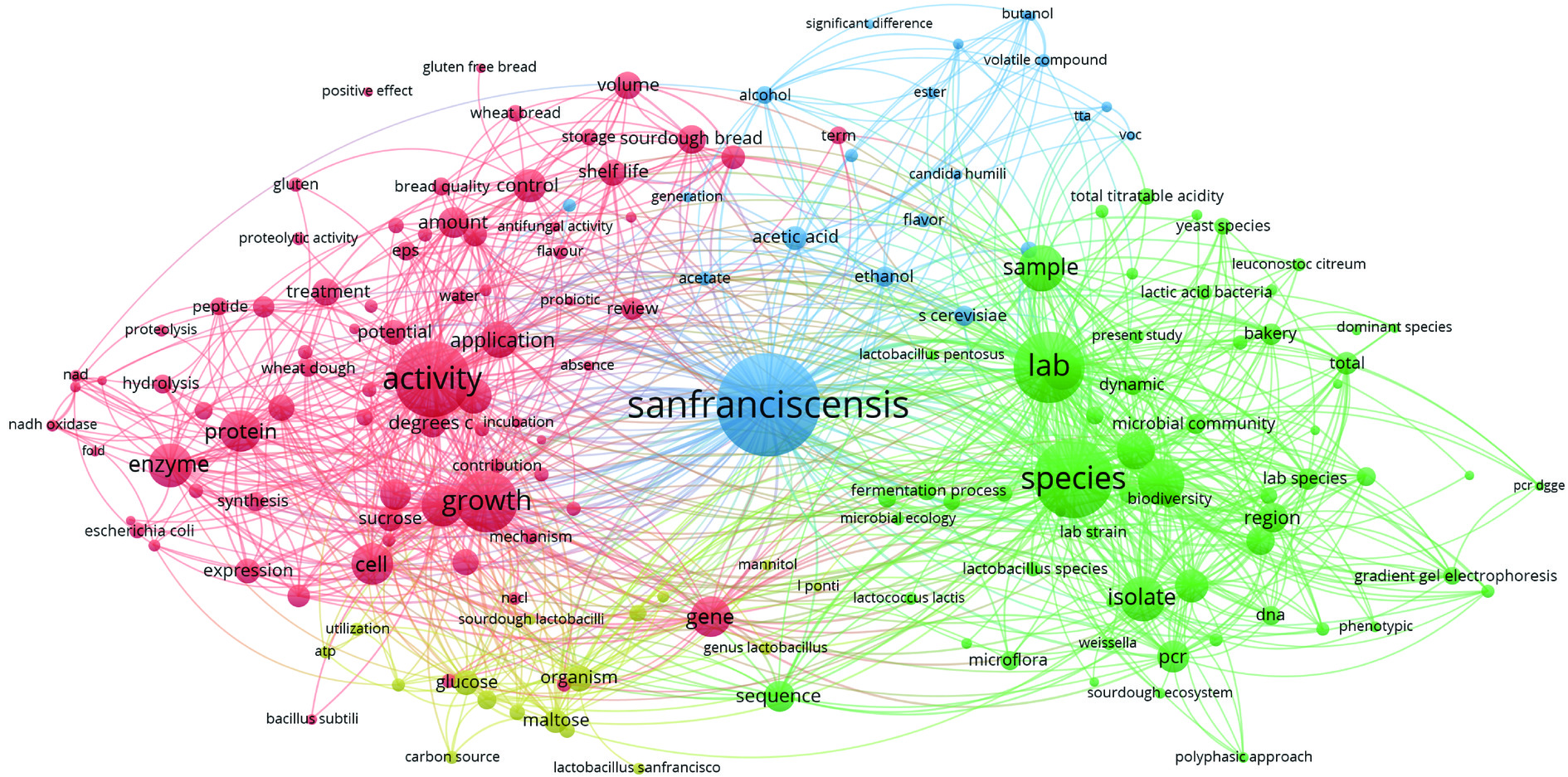
图 2 1971-2024年旧金山果实乳杆菌学术论文发表数量Figure 2. Number of academic literature on F. sanfranciscensis from 1971 to 2024以“Fructilactobacillus sanfranciscensis”or“Lactobacillus sanfranciscensis”or“Lactobacillus sanfrancisco”为检索词,于2024年7月30日在WOS数据库进行检索,对F. sanfranciscensis相关文章的年发文量进行统计分析,结果检索到531篇F. sanfranciscensis相关研究文献,其中,517篇在Web of Science核心数据库中,自20世纪90年代以来关于F. sanfranciscensis的研究热度逐年增加,尤其是近20年来,年均发文量可达21篇。使用VOSviewer_1.6.20版软件对所纳入的文献进行聚类分析并绘制图谱,聚类图中节点越大,关键词在文献中出现次数越多;不同节点表示不同的聚类,根据F. sanfranciscensis相关研究关键词聚类,如图3所示,研究方向主要涉及F. sanfranciscensis菌株的分离筛选鉴定,生长及代谢活性、对葡萄糖、麦芽糖、胞外多糖等的代谢途径及代谢产物的调控和原位酶的表达,对面团和面包的香气形成、质地及货架期等方面的影响。
![]() 图 3 旧金山果实乳杆菌WOS关键词聚类图Figure 3. High-frequency key words of WOS literature on F. sanfranciscensis
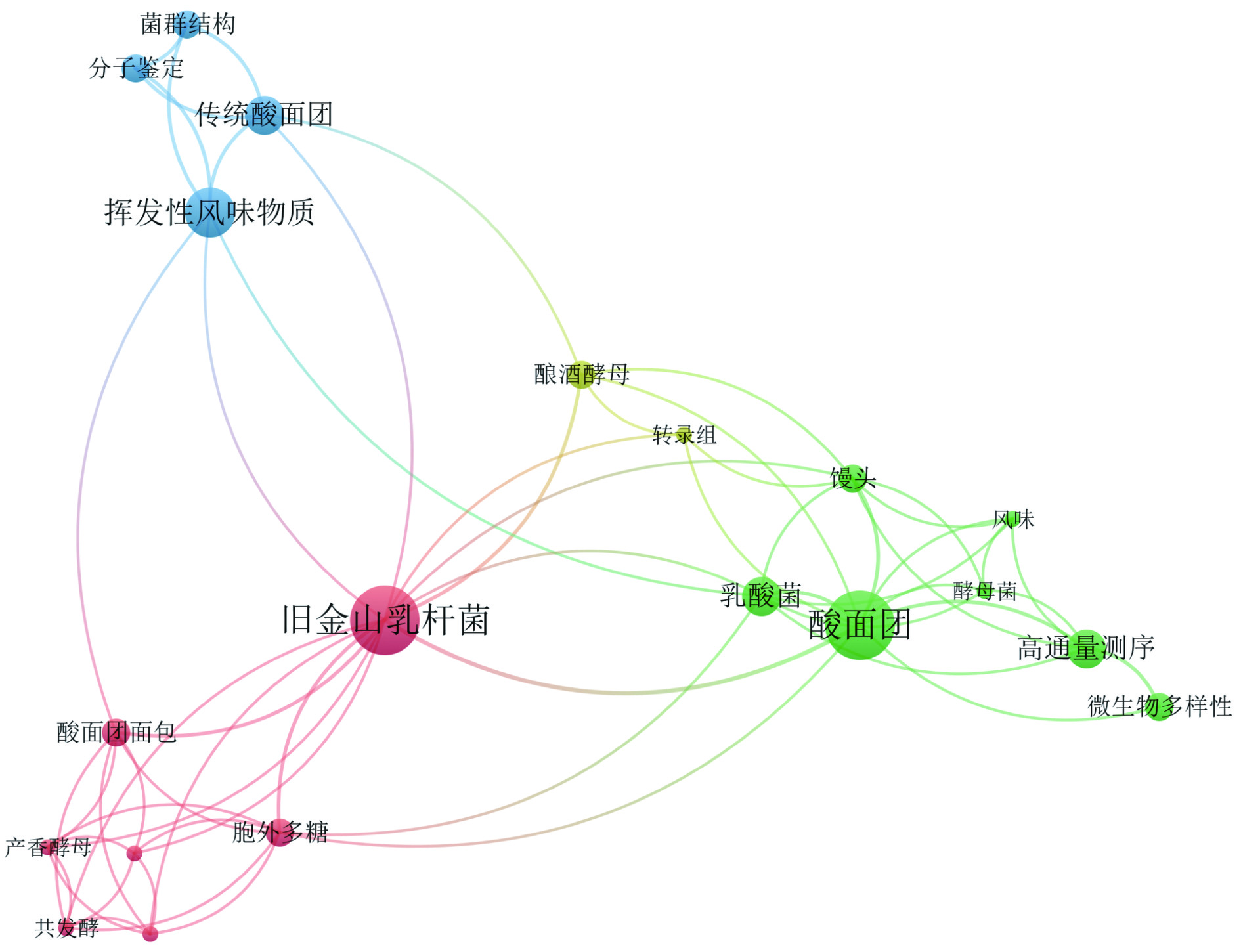
图 3 旧金山果实乳杆菌WOS关键词聚类图Figure 3. High-frequency key words of WOS literature on F. sanfranciscensis以“旧金山果实乳杆菌”或“旧金山乳杆菌”为主题词在CNKI数据库进行检索,只检索到13篇中文相关学术论文结果。如图4显示,在CNKI数据库中,F. sanfranciscensis的研究方向主要涉及酸面团中乳酸菌的多样性,F. sanfranciscensis代谢产生的生物活性物质(胞外多糖)对面包的影响,与酵母菌共发酵在酸面团、馒头及面包的风味物质等方面产生的影响。
![]() 图 4 旧金山果实乳杆菌CNKI关键词聚类图Figure 4. High-frequency key words of CNKI literature on F. sanfranciscensis
图 4 旧金山果实乳杆菌CNKI关键词聚类图Figure 4. High-frequency key words of CNKI literature on F. sanfranciscensis2. 旧金山果实乳杆菌的生物学特征
2.1 旧金山果实乳杆菌的基本特征
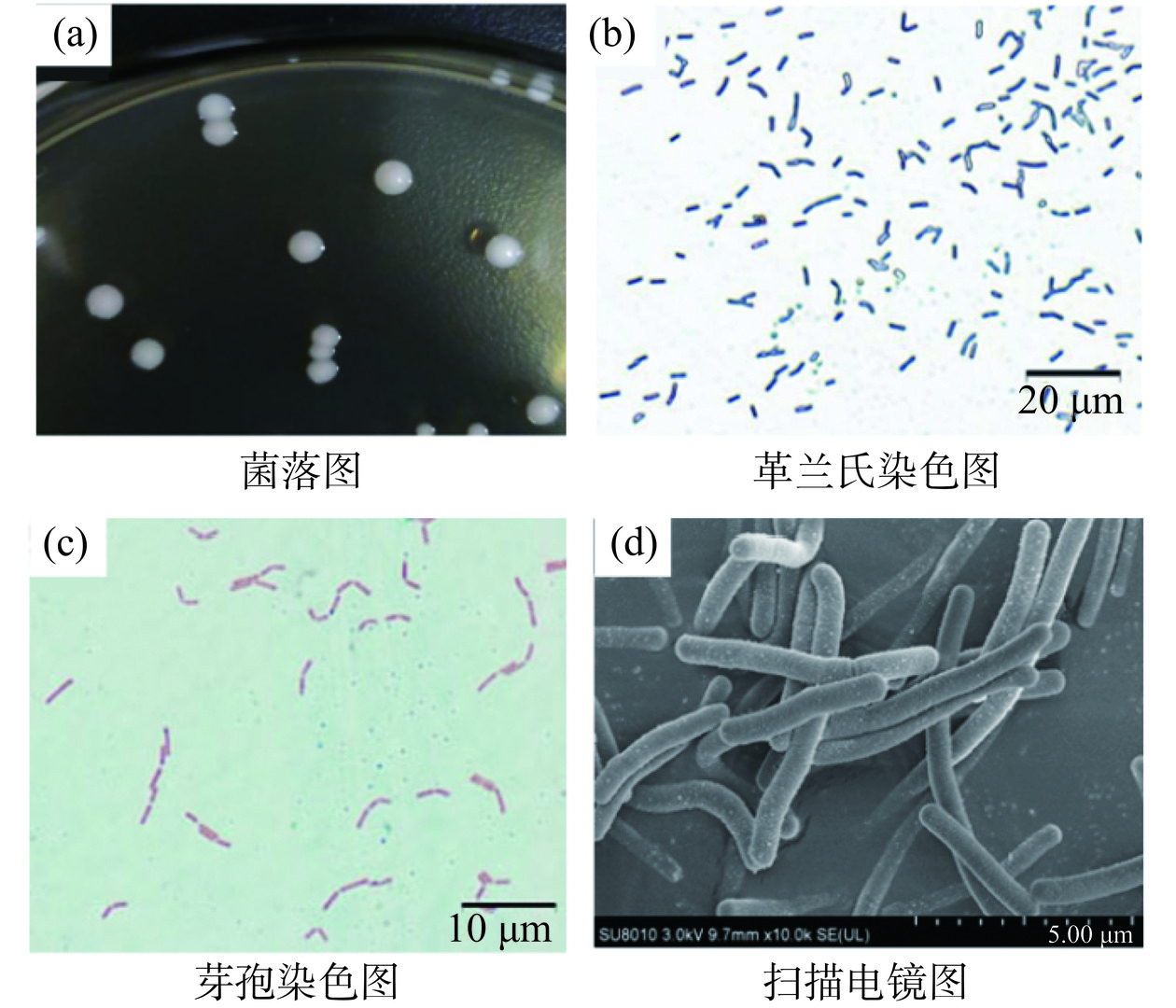
F. sanfranciscensis是异型发酵乳酸菌,G+菌,过氧化氢酶阴性,不产芽孢,不形成孢子,无鞭毛,不运动,细胞常呈短或中长杆状,微弯曲,菌体两端钝圆,通常为(0.6~0.8)×(2~4) µm,单独或成对出现;菌落光滑有光泽,白色不透明呈乳头状规则圆形,F. sanfranciscensis Ls-1001菌株的基本形态图见图5。该菌种属于兼性厌氧,微好氧型乳酸菌,适量CO2可以促进其生长;在13~40 ℃之间存活,最适生长温度为30 ℃,最适pH为5.5[7,22]。在系统发育上,F. sanfranciscensis属于细菌界、厚壁菌门、杆菌纲、乳杆菌目、乳杆菌科、果实乳杆菌属。
![]()
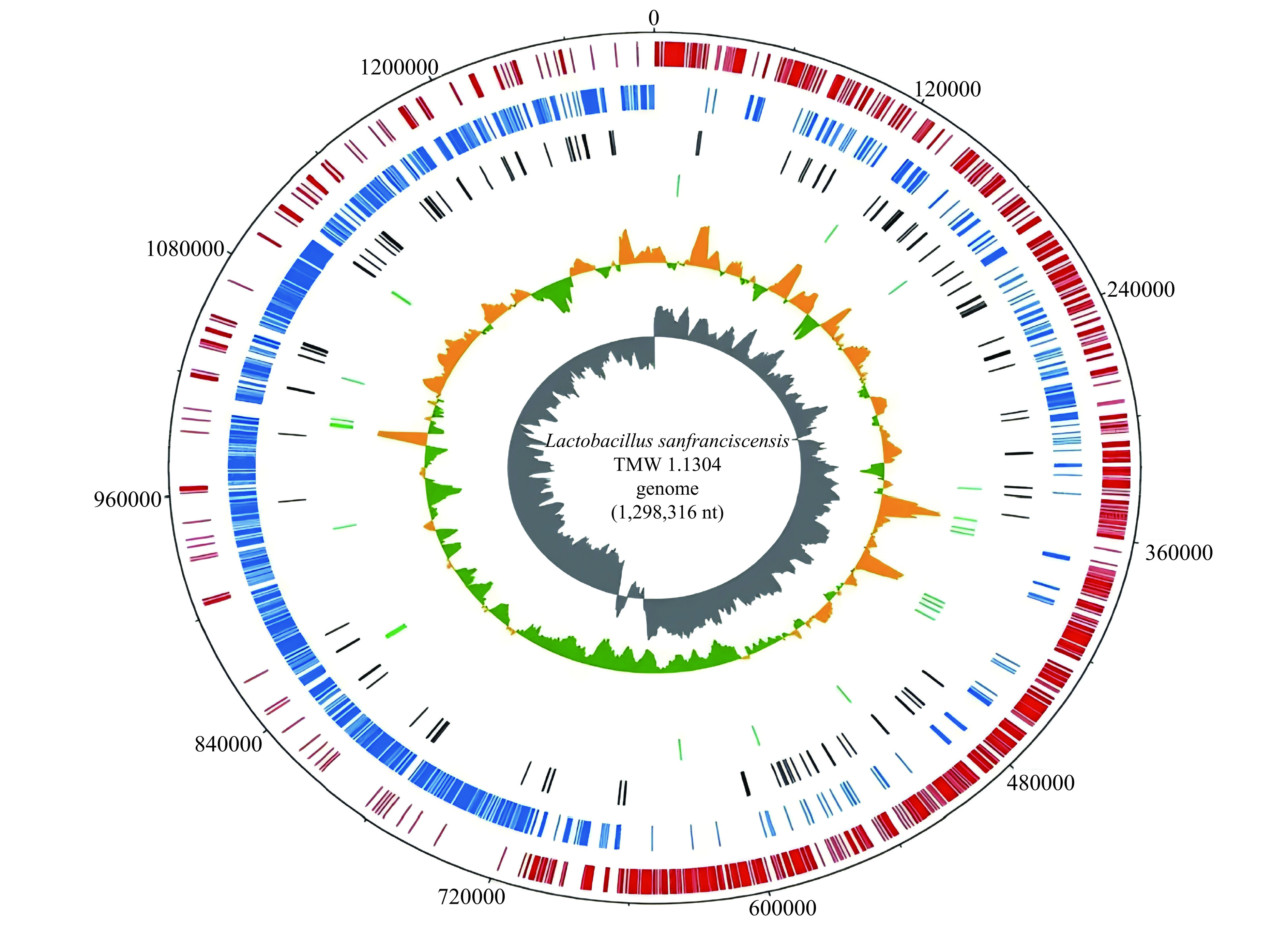
2.2 旧金山果实乳杆菌的基因组特征
F. sanfranciscensis的基因组特征具有生物多样性。He等[24]比较了45株F. sanfranciscensis的基因组,45株菌的基因组相对较小(1.26 Mbp到1.37 Mbp),平均值为1.31 Mbp;GC含量从33.07%到35.20%,GC含量的平均值34.25%;核心基因的主要功能与核苷酸、脂质转运、氨基酸以及碳水化合物代谢有关。此外,Rogalski等[6]通过结合生理学数据和比较基因组学方法研究了24株F. sanfranciscensis,核心基因组仅占泛基因组的43.14%,这些菌株基因组功能的差异主要存在于麦芽糖、果糖和蔗糖代谢以及核苷酸代谢、电子受体的使用和胞外多糖形成等方面。2006年,F. sanfranciscensis TMW 1.1304菌株从商业酸面团中分离出来,之后于2011年,Vogel等[7]对该菌株测序,菌株的基因组圈图见图6,菌株基因组序列包含1298316 bp的环状染色体和pLS1和pLS2这2个质粒,大小分别为58739 bp、18715 bp,GC含量为34.71%。相较于其它乳酸菌而言,F. sanfranciscensis基因组相对较小,每Mbp基因组的核糖体RNA操纵子编码密度高,有助于F. sanfranciscensis的快速生长,也为其适应酸面团中的竞争性生活提供了良好的基础[4,6−7]。
2.3 旧金山果实乳杆菌适应酸面团的代谢特性
酸面团是一种复杂的微生物生态系统,主要由乳酸菌和酵母菌组成,它们通过发酵影响酸面团的酸度、风味和质地,Ⅰ型酸面团由坚硬的面团制成,酸性环境(pH通常在3.8至4.5)、环境温度(通常 < 30 ℃)、水分含量(通常在50%~60%)等属性共同塑造了酸面团的微生物多样性和稳定性[4−5]。F. sanfranciscensis作为传统Ⅰ型酸面团中的关键乳酸菌之一,该菌种对pH和温度的生长要求与I型酸面团发酵条件相匹配,因而其具有快速生长的能力以适应酸面团环境[25]。F. sanfranciscensis的碳水化合物代谢能力也是其适应酸面团环境的关键因素,该菌种是一种专性异型发酵乳酸菌,可将己糖(如葡萄糖、果糖等)发酵成乳酸、乙酸、乙醇、CO2,并将戊糖(如阿拉伯糖、木糖)发酵成乳酸和乙酸,但在碳水化合物代谢方面存在菌株差异。生理和基因组分析表明,F. sanfranciscensis优先利用谷物中的麦芽糖,在麦芽糖磷酸化酶的作用下将麦芽糖转化为1-磷酸-葡萄糖和葡萄糖,为该菌种提供能量和生长所需的碳源,而且这种能量代谢方式不会受到葡萄糖抑制。此外,该菌种还可以代谢果糖促进本身的生长代谢,或者利用果糖作为外部电子受体,在甘露醇脱氢酶的作用下将果糖还原成甘露醇,进而促进NAD循环利用[6]。在蔗糖代谢方面,F. sanfranciscensis菌株特有的蔗糖代谢是由细胞外果聚蔗糖酶活性支持的,而F. sanfranciscensis与酵母菌共培养时,酵母菌可利用蔗糖酶水解蔗糖提供额外的葡萄糖和果糖供F. sanfranciscensis生长[6,25],麦芽糖和果糖的联合使用则是F. sanfranciscensis适应酸面团环境及其与K. humilis共存的关键因素[6]。值得注意的是,F. sanfranciscensis在以戊糖(包括木糖和阿拉伯糖)为唯一碳源上不生长,可能是由于阿拉伯糖和木糖代谢所必需的许多基因的缺失或突变[6]。此外,在酸面团环境中,F. sanfranciscensis具备利用NADH氧化还原酶来抵御氧化胁迫以及调节硫醇代谢的能力,并可通过代谢戊糖获取能量,除此之外,谷胱甘肽和半胱氨酸代谢也是F. sanfranciscensis应对氧化应激反应的关键因素[22,25]。F. sanfranciscensis对特定糖类的偏好和对某些代谢途径的限制,反映了其在长期进化过程中形成的特定生态适应性,这种适应性使得F. sanfranciscensis能够在酸面团生态位中稳定存在,并在发酵面食中发挥重要作用。
3. 旧金山果实乳杆菌对发酵面食的影响
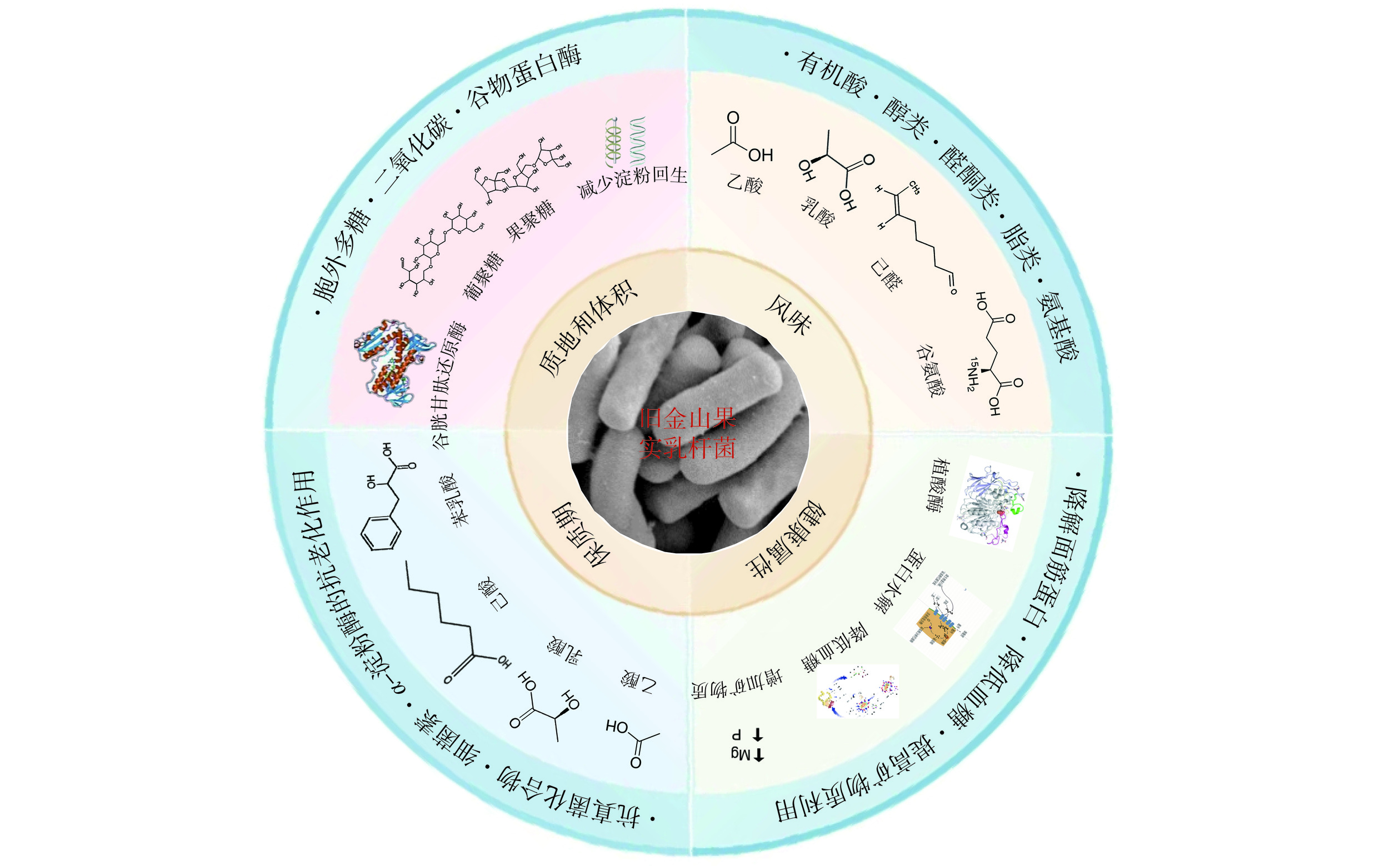
F. sanfranciscensis代谢产生抗氧化化合物、抗真菌肽、短链脂肪酸、有机酸、胞外多糖及挥发物,在改善酸面团感官、质地、货架期、风味和多种营养健康属性方面具有重要作用[4−5,26],F. sanfranciscensis对发酵面食的影响如图7所示。
![]() 图 7 旧金山果实乳杆菌对发酵面食的影响Figure 7. Influence of F. sanfranciscensis on the quality of fermented pasta
图 7 旧金山果实乳杆菌对发酵面食的影响Figure 7. Influence of F. sanfranciscensis on the quality of fermented pastaF. sanfranciscensis有助于酸面团及发酵面食的芳香特性,这主要是由于其在发酵过程中代谢碳水化合物产生的酸化效应,乙酸乙酯、醇类(乙醇、1-丙醇、2-甲基-1-戊醇、1-庚醇和1-辛醇)、醛类(3-甲基-1-丁醛、庚醛、反式-2-庚醛、辛醛和壬醛)和乙酸是该菌种产生的主要化合物[15]。研究表明F. sanfranciscensis与己醛等醛类含量呈正相关,还与挥发性有机化合物(包括高水平的乙酸含量)的生成相关[27−29]。F. sanfranciscensis可代谢果糖产生甘露醇[30],脱酰胺作用将谷氨酰胺转化为谷氨酸,或将精氨酸代谢为鸟氨酸,这些代谢产物的产生进一步丰富了面包风味[4,15]。此外,F. sanfranciscensis和S. cerevisiae之间的相互作用被认为是通过改变3-甲基-1-丁醇、苯乙醇、丁醇、乙酸及乙醇等挥发性代谢物的相对产量来影响面包风味的重要因素[31]。从应用的角度来看,可通过选择F. sanfranciscensis与酵母菌的组合并适当整合小麦粉中的可溶性碳水化合物来丰富发酵面食的风味。
F. sanfranciscensis发酵有助于增强发酵面食的营养特性,可用于制备适合乳糜泻患者、糖尿病患者和对面包酵母过敏人群食用的替代功能性发酵面食产品[15]。植酸存在于所有谷物中,并与面粉中的矿物质形成不溶性复合物,从而降低其生物利用度,因此在饮食中过量的植酸会产生负面影响,Subaşı等[32]利用F. sanfranciscensis发酵酸面团制备的全麦面包具有更高的营养价值,F. sanfranciscensis的酸化作用激活了植酸酶活性,从而降低发酵面食中植酸含量,同时低pH也会导致植酸复合物的溶解,因此,经过F. sanfranciscensis发酵提高了矿物质的生物利用度,同时获得较高的膳食纤维、游离酚含量和抗氧化活性。F. sanfranciscensis发酵酸面团营造酸性条件会降解淀粉周围的面筋网络,并通过限制淀粉颗粒的溶胀和糊化来减少淀粉分解(乳酸可防止淀粉糊化并减少其消化),乙酸、丙酸、酚类化合物、氨基酸和肽的产生也会降低发酵面食的升糖指数(glycemic index,GI)[33]。此外,F. sanfranciscensis通过参与谷蛋白水解和麸质解聚,利用自身的谷胱甘肽还原酶生成还原氧化型谷胱甘肽降低麸质含量,从而加速过敏原降解[34];郭晶斐等[35]利用F. sanfranciscensis Gm4发酵有效降解了麦谷蛋白大聚体,并降低了麸质蛋白的致敏性,因而,F. sanfranciscensis可用于开发无麸质发酵面食。此外,Omedi等[36]利用F. sanfranciscensis发酵酸面团表现出血管紧张素-I转换酶抑制活性,这可能是由于菌株特异性蛋白水解活性。
发酵面食在储存过程中,会发生不同的物理化学变化(如老化、变硬)和微生物变化(如变质、霉菌生长),从而降低发酵面食的质量。变质和真菌污染是导致发酵面食保质期缩短的主要原因,并因产品和储存时间的不同而有所差异[5],发酵面食产品中最常见的腐败真菌包括青霉属、曲霉属、粘菌属、镰刀菌属和根霉属等[12]。F. sanfranciscensis通过产生己酸、乙酸、甲酸及类似于弱酸性防腐剂苯乳酸等物质发挥抑菌活性[4],延长发酵面食的货架期。张国华等[23]研究表明F. sanfranciscensis Ls-1001有一定的抑菌性,对金黄色葡萄球菌、大肠杆菌和白色念珠菌具有抑制作用。F. sanfranciscensis CB1产生的已酸是主要有效的抗霉菌物质,可与其他酸如丙酸、丁酸和戊酸起协同作用[37],在抑制面包中的镰刀菌、青霉菌、曲霉菌和白色念珠菌的生长中发挥关键作用。除了产生有机酸外,F. sanfranciscensis代谢产生的另一种具有抑菌活性的物质是细菌素,细菌素是抗菌肽或小蛋白质,可抑制通常与生产菌株密切相关的微生物。从F. sanfranciscensis C57中分离出的细菌素具有良好的热稳定性(100 ℃,20 min),对脂肪酶和α-淀粉酶不敏感,具有杀菌或溶菌作用,可抑制单核细胞增生李斯特菌[38]。此外,通常认为面包变硬主要是由淀粉组分的再结晶引起的,涉及支链淀粉,而F. sanfranciscensis可通过产生特定有机酸和降低硬度和淀粉回流率来延缓面包老化,酸化程度相当低,淀粉回生速率则不受影响,而由F. sanfranciscensis 57、L. plantarum 13和S. cerevisiae 141共同发酵的酸面团能够延缓老化速率[15]。有机酸和细菌素的产生、淀粉水解和面筋蛋白水解是F. sanfranciscensis的理想特性,对延缓发酵面食的老化和延长酸面团产品货架期起到积极作用,可用于生产不添加化学物质且可以延长保质期的发酵面食。
F. sanfranciscensis通过自身的代谢活动来改善面团的流变性、体积和质地,比如产生具有亲水胶体活性的胞外多糖,改善烘焙食品的结构和体积、增加其柔软度和延迟老化[4]。研究表明,添加F. sanfranciscensis胞外多糖可增加面团的粘性和面包的体积,改善面团面筋网络结构、降低面团强度和弹性、减弱有机酸的作用并产生低聚糖,有助于提高无麸质食品营养价值[39−40]。F. sanfranciscensis可产生用于改善烘焙食品的结构特性的果聚糖[41],Seitter等[42]以市售亲水胶体为对照,研究发现使用添加的酸面团(含F. sanfranciscensis中提取的果聚糖)发酵制备的面包体积更大。除此之外,F. sanfranciscensis发酵后酸面团的pH在4.0以下,此时谷物蛋白酶被激活,在谷物蛋白酶的作用下酸面团中面筋的解聚作用使得面团更柔软[43]。有研究表明,除了谷物蛋白酶对面筋的解聚作用,游离硫醇的积累(特别是F. sanfranciscensis的谷胱甘肽还原酶活性)也可能有助于改善面包体积和面包屑质地[44−45]。此外,F. sanfranciscensis可通过影响淀粉的胶凝和膨胀特性以及面粉的糊状特性来影响面包质地[4]。F. sanfranciscensis通过产生胞外多糖、激活谷物蛋白酶和减少淀粉回生,来改善面团流变性质、增加面包体积和柔软度,并可能通过谷胱甘肽还原酶活性进一步提升发酵面食的品质。
4. 结论
F. sanfranciscensis是酸面团中常见的乳酸菌,本文系统分析了F. sanfranciscensis在酸面团中的普遍性,生物学特性,分类学地位变迁及对馒头、面包等发酵面食风味、质地和营养价值等方面的影响。目前,该菌种已列入了欧洲、韩国的可食用菌种名单,但尚未列入我国的《可用于食品的菌种名单》中,究其原因是我国酸面团尚未实现产业化,用于发酵酸面团的F. sanfranciscensis的研究基础相对薄弱,理论和技术支撑有待完善。因此,开展该菌种/株的安全性评价、全基因组测序、高密度培养及对发酵馒头、包子、馕等传统东方面食品质影响方面的研究,可为推动F. sanfranciscensis列入我国的食品用微生物菌种名单奠定理论基础,同时为传统面食发酵剂产业化应用提供科学依据和技术支持。
-
![]()
图 1 旧金山果实乳杆菌的分类学地位变迁
Figure 1. Evolution of the taxonomic status of F. sanfranciscensis
![]()
图 2 1971-2024年旧金山果实乳杆菌学术论文发表数量
Figure 2. Number of academic literature on F. sanfranciscensis from 1971 to 2024
![]()
图 3 旧金山果实乳杆菌WOS关键词聚类图
Figure 3. High-frequency key words of WOS literature on F. sanfranciscensis
![]()
图 4 旧金山果实乳杆菌CNKI关键词聚类图
Figure 4. High-frequency key words of CNKI literature on F. sanfranciscensis
![]()
-
[1] 陈绍军, 吴兆苏. 从我国小麦、面食及其加工工具的发展历史试谈馒头的起源问题[J]. 农业考古,1994(1):219−225. [CHEN Shaojun, WU Zhaosu. On the origin of steamed bread from the development history of wheat, pasta and their processing tools in China[J]. Agricultural Archaeology,1994(1):219−225.] CHEN Shaojun, WU Zhaosu. On the origin of steamed bread from the development history of wheat, pasta and their processing tools in China[J]. Agricultural Archaeology, 1994(1): 219−225.
[2] EMERALD M, RAJAURIA G, KUMAR V. Novel fermented grain-based products [M]//OJHA K S, TIWARI B K. Novel Food Fermentation Technologies. Cham; Springer International Publishing. 2016:263-277.
[3] VAN KERREBROECK S, MAES D, DE VUYST L. Sourdoughs as a function of their species diversity and process conditions, a meta-analysis[J]. Trends in Food Science & Technology,2017,68:152−159.
[4] ZHANG G, TU J, SADIQ F A, et al. Prevalence, genetic diversity, and technological functions of the Lactobacillus sanfranciscensis in sourdough:A review[J]. Comprehensive Reviews in Food Science and Food Safety,2019,18(4):1209−1226. doi: 10.1111/1541-4337.12459
[5] ARORA K, AMEUR H, POLO A, et al. Thirty years of knowledge on sourdough fermentation:A systematic review[J]. Trends in Food Science & Technology,2021,108:71−83.
[6] ROGALSKI E, EHRMANN M A, VOGEL R F. Intraspecies diversity and genome-phenotype-associations in Fructilactobacillus sanfranciscensis[J]. Microbiological Research,2021,243:126625. doi: 10.1016/j.micres.2020.126625
[7] VOGEL R F, PAVLOVIC M, EHRMANN M A, et al. Genomic analysis reveals Lactobacillus sanfranciscensis as stable element in traditional sourdoughs[J]. Microbial Cell Factories, 2011, 10 Suppl 1(Suppl 1):S6.
[8] 陈卫. 乳酸菌科学与技术[M]. 北京:科学出版社, 2018:828−830. [CHEN Wei. Science and technology of lactic acid bacteria[M]. Beijing:Science Press, 2018:828−830.] CHEN Wei. Science and technology of lactic acid bacteria[M]. Beijing: Science Press, 2018: 828−830.
[9] 韩国食品药品安全部公告第 2024 - 35 号[EB/OL]. (2024.07. 10, 修订) [2024-09-30]. [Bulletin of the ministry of food and drug safety No. 2024-35 [EB/OL]. (2024.07. 10, Revised) [2024-09-30]. https://various.foodsafetykorea.go.kr/fsd/#/ext/Document/FC.] Bulletin of the ministry of food and drug safety No. 2024-35 [EB/OL]. (2024.07. 10, Revised) [2024-09-30]. https://various.foodsafetykorea.go.kr/fsd/#/ext/Document/FC.
[10] 姚粟, 王鹏辉, 白飞荣, 等. 中国传统发酵食品用微生物菌种名单研究[J]. 第二版. 食品与发酵工业,2022,48(1):272−307. [YAO Su, WANG Penghui, BAI Feirong, et al. Research on the inventory of microbial food cultures in Chinese traditional fermented foods[J]. 2nd edition. Food and Fermentation Industries,2022,48(1):272−307.] YAO Su, WANG Penghui, BAI Feirong, et al. Research on the inventory of microbial food cultures in Chinese traditional fermented foods[J]. 2nd edition. Food and Fermentation Industries, 2022, 48(1): 272−307.
[11] SUO B, CHEN X, WANG Y. Recent research advances of lactic acid bacteria in sourdough:Origin, diversity, and function[J]. Current Opinion in Food Science,2021,37:66−75. doi: 10.1016/j.cofs.2020.09.007
[12] BOIOCCHI F, PORCELLATO D, LIMONTA L, et al. Insect frass in stored cereal products as a potential source of Lactobacillus sanfranciscensis for sourdough ecosystem[J]. Journal of Applied Microbiology,2017,123(4):944−955. doi: 10.1111/jam.13546
[13] ZHENG Z, ZHAO M, ZHANG Z, et al. Lactic acid bacteria are prevalent in the infrabuccal pockets and crops of ants that prefer aphid honeydew[J]. Frontiers in Microbiology,2022,12:785016. doi: 10.3389/fmicb.2021.785016
[14] KLINE L, SUGIHARA T F. Microorganisms of the San Francisco sourdough bread process:II. Isolation and characterization of undescribed bacterial species responsible for the souring activity[J]. Applied Microbiology,1971,21(3):459−465. doi: 10.1128/am.21.3.459-465.1971
[15] YAZAR G, TAVMAN Ş. Functional and technological aspects of sourdough fermentation with Lactobacillus sanfranciscensis[J]. Food Engineering Reviews,2012,4(3):171−190. doi: 10.1007/s12393-012-9052-1
[16] BACK W, BOHAK I, EHRMANN M, et al. Revival of the species Lactobacillus lindneri and the design of a species specific oligonucleotide probe[J]. Systematic and Applied Microbiology,1996,19(3):322−325. doi: 10.1016/S0723-2020(96)80058-0
[17] SPICHER G, SCHRÖDER R. Die Mikroflora des Sauerteiges[J]. Zeitschrift Für Lebensmittel-Untersuchung und Forschung,1978,167(5):342−354.
[18] WEISS N, SCHILLINGER U. Lactobacillus sanfrancisco sp. nov. , nom. rev[J]. Systematic and Applied Microbiology, 1984, 5(2):230-232.
[19] TRÜPER H G, CLARI L. Taxonomic note :Erratum and correction of further specific epithers formed as substantives (nonus) 'in apposition'[J]. International Journal of Systematic and Evolutionary Microbiology, 1998, 48:615.
[20] ZHENG J, WITTOUCK S, SALVETTI E, et al. A taxonomic note on the genus Lactobacillus:Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae[J]. International Journal of Systematic and Evolutionary Microbiology,2020,70(4):2782−2858. doi: 10.1099/ijsem.0.004107
[21] 于学健, 胡海蓉, 曹艳花, 等. 乳杆菌属分类学地位变迁后菌种名称英解汉译检索表(三)[J]. 食品与发酵工业,2020,46(17):296−300. [YU Xuejian, HU Hairong, CAO Yanhua, et al. Retrieval table of English interpretation and Chinese translation of bacterial species names after the taxonomic status change of Lactobacillus (3)[J]. Food and Fermentation Industries,2020,46(17):296−300.] YU Xuejian, HU Hairong, CAO Yanhua, et al. Retrieval table of English interpretation and Chinese translation of bacterial species names after the taxonomic status change of Lactobacillus (3)[J]. Food and Fermentation Industries, 2020, 46(17): 296−300.
[22] GÄNZLE M G, EHMANN M, HAMMES W P. Modeling of growth of Lactobacillus sanfranciscensis and Candida milleri in response to process parameters of sourdough fermentation[J]. Applied and Environmental Microbiology,1998,64(7):2616−2623. doi: 10.1128/AEM.64.7.2616-2623.1998
[23] 张国华, 张纬珍, 梁武龙, 等. 一株高产胞外多糖乳酸菌的鉴定及其特性研究[J]. 中国食品学报,2020,20(12):229−238. [ZHANG Guohua, ZHANG Weizhen, LIANG Wulong, et al. Studies on identification and characterization of a high-yield exopolysaccharides lactic acid bacteria[J]. Journal of Chinese Institute of Food Science and Technology,2020,20(12):229−238.] ZHANG Guohua, ZHANG Weizhen, LIANG Wulong, et al. Studies on identification and characterization of a high-yield exopolysaccharides lactic acid bacteria[J]. Journal of Chinese Institute of Food Science and Technology, 2020, 20(12): 229−238.
[24] HE X, YU Y, KEMPERMAN R, et al. Comparative genomics reveals genetic diversity and variation in metabolic traits in Fructilactobacillus sanfranciscensis strains[J]. Microorganisms,2024,12(5):845. doi: 10.3390/microorganisms12050845
[25] GÄNZLE M G, ZHENG J. Lifestyles of sourdough lactobacilli – Do they matter for microbial ecology and bread quality?[J]. International Journal of Food Microbiology,2019,302:15−23. doi: 10.1016/j.ijfoodmicro.2018.08.019
[26] SADEGHI A, EBRAHIMI M, HAJINIA F, et al. Food Omics as a promising strategy to study the effects of sourdough on human health and nutrition, as well as product quality and safety; Back to the future[J]. Trends in Food Science & Technology,2023,136:24−47.
[27] SABACH O, BUHNIK-ROSENBLAU K, KESTEN I, et al. The rise of the sourdough:Genome-scale metabolic modeling-based approach to design sourdough starter communities with tailored-made properties[J]. International Journal of Food Microbiology,2023,407:110402. doi: 10.1016/j.ijfoodmicro.2023.110402
[28] BOYACI GUNDUZ C P, AGIRMAN B, GAGLIO R, et al. Evaluation of the variations in chemical and microbiological properties of the sourdoughs produced with selected lactic acid bacteria strains during fermentation[J]. Food Chemistry:X,2022,14:100357.
[29] SANMARTÍN G, SÁNCHEZ-ADRIÁ I E, PRIETO J A, et al. Bioprospecting of sourdough microbial species from artisan bakeries in the city of Valencia[J]. Food Microbiology,2024,120:104474. doi: 10.1016/j.fm.2024.104474
[30] SAHIN A W, ZANNINI E, COFFEY A, et al. Sugar reduction in bakery products:Current strategies and sourdough technology as a potential novel approach[J]. Food Research International,2019,126:108583. doi: 10.1016/j.foodres.2019.108583
[31] LIU T, SHI Y, LI Y, et al. The mutual influence of predominant microbes in sourdough fermentation:Focusing on flavor formation and gene transcription[J]. Foods,2022,11(15):2373. doi: 10.3390/foods11152373
[32] SEIS SUBAŞı A, ERCAN R. The effects of wheat variety, sourdough treatment and sourdough level on nutritional characteristics of whole wheat bread[J]. Journal of Cereal Science,2023,110:103637. doi: 10.1016/j.jcs.2023.103637
[33] FEKRI A, ABEDINZADEH S, TORBATI M, et al. Considering sourdough from a biochemical, organoleptic, and nutritional perspective[J]. Journal of Food Composition and Analysis,2024,125:105853. doi: 10.1016/j.jfca.2023.105853
[34] JÄNSCH A, KORAKLI M, VOGEL R F, et al. Glutathione reductase from Lactobacillus sanfranciscensis DSM20451T:Contribution to oxygen tolerance and thiol exchange reactions in wheat sourdoughs[J]. Applied and Environmental Microbiology,2007,73(14):4469−4476. doi: 10.1128/AEM.02322-06
[35] 郭晶斐, 贺霞霞, 涂建, 等. 酸面团中优势乳酸菌和酵母菌对麦谷蛋白结构的影响[J]. 中国酿造,2022,41(4):142−146. [GUO Jingfei, HE Xiaxia, TU Jian, et al. Effect of dominant Lactobacillus and yeast in sourdough on glutenin structure[J]. China Brewing,2022,41(4):142−146.] doi: 10.11882/j.issn.0254-5071.2022.04.024 GUO Jingfei, HE Xiaxia, TU Jian, et al. Effect of dominant Lactobacillus and yeast in sourdough on glutenin structure[J]. China Brewing, 2022, 41(4): 142−146. doi: 10.11882/j.issn.0254-5071.2022.04.024
[36] OMEDI J O, HUANG W, SU X, et al. Effect of five lactic acid bacteria starter type on angiotensin-I converting enzyme inhibitory activity and emulsifying properties of soy flour sourdoughs with and without wheat bran supplementation[J]. Journal of Cereal Science,2016,69:57−63. doi: 10.1016/j.jcs.2016.02.008
[37] CORSETTI A, GOBBETTI M, ROSSI J, et al. Antimould activity of sourdough lactic acid bacteria:Identification of a mixture of organic acids produced by Lactobacillus sanfrancisco CB1[J]. Applied Microbiology and Biotechnology,1998,50(2):253−256. doi: 10.1007/s002530051285
[38] CORSETTI A, GOBBETTI M, SMACCHI E. Antibacterial activity of sourdough lactic acid bacteria isolation of a bacteriocin like inhibitory substance from Lactobacillus sanfrancisco[J]. Food Microbiology,1996,13:447−456. doi: 10.1006/fmic.1996.0051
[39] 苏珊, 王玉婷, 吴洁, 等. 酸面团中的优势微生物及其相互作用研究进展[J]. 食品与发酵工业,2023,49(7):316−323. [SU Shan, WANG Yuting, WU Jie, et al. Research progress on dominant microorganisms and their interactions in sourdough[J]. Food and Fermentation Industries,2023,49(7):316−323.] SU Shan, WANG Yuting, WU Jie, et al. Research progress on dominant microorganisms and their interactions in sourdough[J]. Food and Fermentation Industries, 2023, 49(7): 316−323.
[40] CODA R, XU Y, MORENO D S, et al. Performance of Leuconostoc citreum FDR241 during wheat flour sourdough type I propagation and transcriptional analysis of exopolysaccharides biosynthesis genes[J]. Food Microbiology,2018,76:164−172. doi: 10.1016/j.fm.2018.05.003
[41] CORSETTI A, SETTANNI L. Lactobacilli in sourdough fermentation[J]. Food Research International,2007,40(5):539−558. doi: 10.1016/j.foodres.2006.11.001
[42] SEITTER M, FLEIG M, SCHMIDT H, et al. Effect of exopolysaccharides produced by Lactobacillus sanfranciscensis on the processing properties of wheat doughs[J]. European Food Research and Technology,2020,246(3):461−469. doi: 10.1007/s00217-019-03413-x
[43] CLARKE C I, ARENDT E K. A review of the application of sourdough technology to wheat breads[J]. Advances in Food and Nutrition Research,2005,49:137−161.
[44] XU D, TANG K, HU Y, et al. Effect of glutathione dehydrogenase of Lactobacillus sanfranciscensis on gluten properties and bread volume in Type I wheat sourdough bread[J]. Journal of Agricultural and Food Chemistry,2018,66(37):9770−9776. doi: 10.1021/acs.jafc.8b03298
[45] TANG K X, ZHAO C J, GÄNZLE M G. Effect of glutathione on the taste and texture of Type I sourdough bread[J]. Journal of Agricultural and Food Chemistry,2017,65(21):4321−4328. doi: 10.1021/acs.jafc.7b00897
-
期刊类型引用(1)
1. 康潇云. 食品安全检测中的生物传感器技术应用与创新. 食品安全导刊. 2024(29): 187-189 .  百度学术
百度学术
其他类型引用(0)
 下载:
下载:

 下载:
下载:






计量
- 文章访问数: 85
- HTML全文浏览量: 20
- PDF下载量: 21
- 被引次数: 1



