


Correlation Analysis between Microbial Diversity and Flavor Compounds in Sichuan Fermented Sausage
-
摘要: 为研究川味发酵香肠微生物多样性与风味物质的相关性,以不同发酵时间的川味香肠为研究对象,采用高通量测序技术结合电子舌、电子鼻和气相色谱离子迁移技术(GC-IMS),分析香肠发酵过程中微生物优势菌群和风味物质的变化规律,通过Pearson相关性分析微生物多样性与挥发性风味物质的相关性。结果表明,随香肠发酵时间延长,微生物的种数和丰度上升,共检测出12种菌属,其中乳酸杆菌属和魏斯氏菌属占优势地位;非挥发性风味物质方面,游离氨基酸、核苷酸和呈苦味、鲜味肽的含量逐渐增多;挥发性风味物质方面,共检测出52种物质。其中,醇类、萜烯类、噻唑等物质浓度逐渐减少,酯类、吡嗪类、醛类浓度逐渐增多。GC-IMS结果进一步显示,发酵前期和中期的特征风味物质相似,主要由柠檬烯、β-蒎烯(M)、乙醇、β-月桂烯、1,1-二乙氧基乙烷(M)组成。发酵至35 d时刺激性风味物质含量显著降低;发酵稳定期的酯类、醛类、吡嗪类等特征风味物质种类和含量趋于稳定。Pearson相关性分析显示:乳酸杆菌属、不动杆菌属、片球菌属、假单胞菌属和金黄杆菌属这5种优势菌属与37种挥发性风味物质呈正相关,其中优势菌属乳酸杆菌属与醛类、醇类、酯类呈正相关(r>0.6),次优势菌属魏斯氏菌属与醇类和酯类呈负相关(r<−0.4);表明微生物发酵优势菌群对香肠风味物质合成密切相关,为开发高效绿色的发酵剂调控川味发酵香肠的风味品质提供了科学依据和理论基础。
-
关键词:
- 发酵香肠 /
- 微生物多样性 /
- 风味物质 /
- 气相色谱离子迁移技术 /
- 相关性
Abstract: To investigate the correlation between microbial diversity and flavor compounds in Sichuan-style fermented sausages, sausages at different fermentation times were studied using high-throughput sequencing technology in conjunction with electronic tongue, electronic nose, and gas chromatography-ion mobility spectrometry (GC-IMS). The changes in dominant microbial communities and flavor compounds during sausage fermentation were analyzed, and the correlation between microbial diversity and volatile flavor compounds was assessed using Pearson correlation analysis. Results showed that with prolonged sausage fermentation time, the number and abundance of microbes increased, with a total of 12 genera detected, dominated by Lactobacillus and Weissella. In terms of non-volatile compounds, the content of free amino acids, nucleotides, and peptides with bitter and umami tastes gradually increased. Regarding volatile flavor compounds, 52 substances were detected. The concentrations of alcohols, terpenes, and thiazoles decreased gradually, while esters, pyrazines, and aldehydes increased. GC-IMS results further revealed that the characteristic flavor compounds in the early and middle stages of fermentation were similar, mainly composed of limonene, β-myrcene (M), ethanol, β-caryophyllene, and 1,1-diethoxyethane (M). The content of pungent flavor compounds significantly decreased at 35 d of fermentation. The types and concentrations of esters, aldehydes, and pyrazines in the stable fermentation period tended to stabilize. Pearson correlation analysis indicated that five dominant genera including Lactobacillus, Acinetobacter, Pediococcus, Pseudomonas, and Chryseobacterium were positively correlated with 37 volatile flavor compounds. Specifically, Lactobacillus was positively correlated with aldehydes, alcohols, and esters (r>0.6), while Weissella, a subdominant genus, was negatively correlated with alcohols and esters (r<−0.4). These findings suggest a close relationship between dominant microbial communities and the synthesis of sausage flavor compounds, providing a scientific basis and theoretical foundation for the development of efficient and environmentally friendly fermentation agents to regulate the flavor quality of Sichuan-style fermented sausages.-
Keywords:
- fermented sausage /
- microbial diversity /
- flavor substances /
- GC-IMS /
- correlation
-
川味香肠属于发酵香肠,又称为川式香肠,是我国西南地区的传统特色发酵肉制品之一,主要以“三椒”(辣椒、花椒、胡椒)作辅料与肉混合灌进肠衣,在自然条件下经过微生物发酵产生有机酸或醇酯类,最终制成美味的香肠[1−2],因其麻辣咸甜、多味生香、风味醇厚独特深受消费者青睐。
香肠的风味成分主要来源于发酵过程中糖类、蛋白质、脂质氧化分解[3],通过微生物的复杂代谢产生氨基酸、核苷酸、寡糖、小分子肽等滋味物质和醇类、酯类、酮类、醛类、萜烯类、吡嗪类等挥发性风味物质,进而赋予香肠特殊的风味[4]。受发酵环境、时间、盐分、原料肉、调味料、辅料等多种因素的影响,发酵香肠微生物菌群复杂多样,微生物自身代谢及菌群间相互作用产生多种功能酶和风味物质,使得香肠风味物质组成和品质特性存在差异。杨小莹等[5]研究发现以不同菌种作为微生物发酵剂都能对发酵香肠中杂菌的生长起到不同程度的抑制作用,有利于发酵香肠发色,弹性及咀嚼度的增加,有利于酯类及其他风味物质的生成。黄郑朝等[6]应用高通量测序技术对不同地区香肠中的细菌群落进行分析,发现加工工艺和气候差异是导致香肠中细菌分布不同的主要因素。牛茵等[7]通过利用高通量测序技术、气相色谱-质谱联用法,分析细菌结构与挥发性物质相关性,发现不同加工环境下羊肉香肠的优势菌种葡萄球菌属(Staphylococcus)、环丝菌属(Brochothrix)、乳酸杆菌属(Lactobacillus)决定其挥发性风味种类。张香美等[8]采用气相色谱离子迁移技术分析香肠不同发酵阶段挥发性风味物质变化趋势,乳杆菌科细菌与醇类、酯类、醛类合成密切相关,葡萄球菌属与2-甲基丁醛、3-甲基丁醛呈正相关。目前,川味香肠发酵过程中微生物多样性与特征风味物质变化之间的关系缺乏深入研究,且存在产品品质不稳定等问题,因此结合发酵时间、利用关键微生物菌群调控特征风味的合成,提升川味发酵香肠的风味品质和营养价值是一项有待深入研究的工作。香肠发酵风味的形成与微生物密切相关,存在一系列消长规律,进而影响风味物质动态变化,此外影响发酵过程的因素众多,难以调控。因此,针对微生物菌群结构多样性与风味物质的相关性开展研究,对于改善川味发酵香肠风味品质,提高产品安全性具有重要意义。
本研究以不同发酵时间的川味香肠为研究对象,利用高通量测序技术研究微生物菌群结构变化,气相色谱离子迁移(GC-IMS)技术、电子鼻、电子舌技术研究滋味成分和挥发性风味物质的动态变化,通过Pearson相关性分析揭示发酵过程中及香肠的微生物优势菌群与挥发性风味物质的相关性,为开发高效绿色的微生物发酵剂奠定基础,以期为调控改善发酵肉制品的风味品质提供参考依据。
1. 材料与方法
1.1 材料与仪器
冷鲜猪前腿瘦肉、猪肥膘和猪小肠(肠衣)、辅料:以肉重计,食盐36 g/kg、味精5 g/kg、白糖5 g/kg、料酒20 g/kg、花椒粉2 g/kg、辣椒粉2.4 g/kg、胡椒粉1.6 g/kg、姜泥1.2 g/kg和五香粉1.0 g/kg 成都市龙泉驿区万达广场永辉超市;E.Z.N.A.® Soil DNA提取试剂 美国Omega BioTek公司。
30B型万能高效粉碎机 靖江市辉恒机械制造有限公司;ME204E型天平 梅特勒—托利多国际贸易有限公司;DZ400真空包装机 温州凯瑞机械科技有限公司; XZ-I型绞肉灌肠一体机 广州旭众食品机械有限公司;Flavour Spec®气相离子迁移谱(GC-IMS)联用仪 德国G.A.S.公司;BSP-250 生化培养箱 上海博讯实业有限公司;DLHR-X 200恒温培养振荡器 北京东联哈尔仪器制造有限公司;FOX 4000电子鼻、α-ASTREE型电子舌 法国Alpha MOS公司;TMS-PRO型质构仪 美国FTC公司;S470 pH计 梅特勒-托利多仪器(上海)有限公司。
1.2 实验方法
1.2.1 香肠制作工艺
参照李鹤等[9]的香肠制作方法稍作修改。分别将鲜猪前腿瘦肉、鲜猪肥膘剔除猪皮、骨结缔组织后,按肥瘦比8:2混合,按比例将辅料与原料充分混合,静置0.5 h,将肠衣洗净后灌肠,灌制时保证肠体紧密适宜,打针眼排气后用60 ℃左右的温水冲洗肠体,放置温度为25±1 ℃,湿度为80%条件下发酵42 d,具体取样时间为7、14、21、28、35、42 d,分别标注为A、B、C、D、E、F组。
1.2.2 菌群结构高通量测序分析
参照SUN等[10]的测序方法稍作修改。用E.Z.N.A.®soil试剂盒提取不同发酵时间香肠样品的总DNA。采用595F-806R为引物对细菌16S rRNA的V3~V4进行PCR扩增,根据Illumina MiSeq平台标准操作规程将纯化后的扩增片段构建PE2×300的文库进行评估。利用Illumina MiSeq公司的MiseqPE300平台进行测序,每组设5个重复。
1.2.3 pH的测定
根据《GB 5009.237-2016 食品安全国家标准 食品pH 值的测定》,对不同发酵时间的川味香肠进行pH 测定,并做出适当修改。取5 g粉碎均质后的样品,与90 mL蒸馏水充分混合,静置20 min。将其过滤后取上清液进行pH测定。进行三次测定,取平均值作为结果。
1.2.4 滋味物质的测定
参考袁灿等[11]研究中的电子舌分析方法,并稍作修改。称取10.0 g香肠样品切碎成小颗粒状,加入100 mL蒸馏水溶解,超声波处理15 min,经过滤后,取80 mL无色透明的滤液进行测定。每个样品的酸味、苦味、甜味、鲜味、咸味、鲜回味、苦回味重复测定8次,保留5组稳定数据,并对测定值进行处理和分析。
1.2.5 气味成分的测定
参照CHEN等[12]的测定方法略作修改。准确称量3 g粉碎均匀A、B、C、D、E、F组的香肠置于5 mL 顶空瓶中,加盖密封,室温静置30 min,而后进行电子鼻测定。不同样品重复测定10次,选取其中3次较为平稳测量数据,并利用仪器自带软件进行处理。参数设置:进样准备时间5 s,进样流量为400 mL/min,采样检测时间为120 s,样品信号采集时间选定为89~93 s,传感器清洗时间为120 s。
1.2.6 挥发性风味物质的测定
参照曹辰辰等[13]的方法略作修改。将A、B、C、D、E、F组香肠去掉肠衣后,碾磨成小颗粒状,准确称取1.00 g置于10 mL进样瓶中。进样条件:孵育温度60 ℃,孵育时间15 min,孵育转速:500 r/min,顶空进样。GC条件:MXT-WAX色谱柱(30 m×0.53 mm),色谱柱温度60 ℃;载气:N2(纯度99.999%);载气程序:初始流速5 mL/min,保持5 min,然后在15 min增加至150 mL/min。使用仪器配套的Laboratory Analytical Viewer(LAV)软件和Library Search(LS)软件和内置数据库进行特征风味物质定性分析,Gallery Plot绘制指纹图谱。
1.3 数据处理
以上实验所得数据采用Microsoft Excel 2019、IBM SPSS Statistics 25进行统计、方差分析,P<0.05表示差异显著;采用Origin2018进行主成分、雷达图、热图等多种绘图分析。
2. 结果与分析
2.1 川味发酵香肠发酵过程中微生物α多样性分析
微生物α多样性指在特定生态环境或生态系统内单个群落本身的多样性情况。常见的α多样性指数包括Good’s Coverage指数(GC率)、OTU数、ACE指数、Chao1指数、Simpson指数和Shannon指数等[14],川味香肠微生物α多样性分析见表1。GC率为本次微生物测序数据覆盖率指数,本研究GC率均大于0.97,说明测定样品的深度充分,能够全面分析香肠样品中微生物种类和多样性,表明此次α多样性分析准确性程度较高。OTU数表示微生态细菌种数,与微生物的物种丰富度密切相关,其数值的高低直接反映了测序样本中微生物种类的多样性,该值越高表示微生物种类越多。发酵7 d时,香肠样品OTU数为1029,说明制作香肠的原材料和辅料中的细菌种类较为复杂,随着发酵的进行,OTU数呈增长趋势,可能是由于微生物适应了发酵环境,其中部分微生物代谢产酸,促使其他菌群生长繁殖,抑制了其他有害杂菌生长繁殖;直至发酵42 d结束,OTU数为1852,总体变化趋势上升,发酵结束OTU总数较发酵初期(7 d)增长了79.98%,在发酵周期中微生物种数变化明显。
表 1 川味香肠发酵微生态细菌群落α多样性分析Table 1. Alpha diversity analysis of fermented microecological bacterial communities in Sichuan sausage指标 发酵时间(d) 7 14 21 28 35 42 GC率 0.98 0.97 0.98 1 0.99 1 OTU数 1029 1356 1568 1687 1765 1852 ACE指数 2865 3321 3542 3589 3615 3752 Chao1指数 2985 3102 2895 3365 3125 3621 Shannon指数 6.56 6.12 5.23 6.98 5.98 7.01 Simpson指数 0.43 0.32 0.56 0.77 0.75 0.73 ACE和Chao1都表示微生物群落中物种的丰度指数,其中ACE是以稀有微生物为基础的生物多样性估计指标,其值越高表示群落种类越丰富。从表1可知,ACE指数随发酵进行,指数呈上升趋势,同OTU数变化趋势基本一致,至发酵42 d结束,ACE指数为3752,较发酵初期增长了30.96%。Chao1指数对稀有物种很敏感,是度量物种丰富度的指标。Chao1指数变化波动较小,总体变化趋势上升,至发酵结束时为3621,这表明川味香肠整个发酵周期微生态群落种数和丰度呈上升趋势。
Shannon指数和Simpson指数为微生物群落的多样性指数。其中Shannon指数的数值范围为1~10左右,数值越大群落多样性越高[14]。从表1可知,Shannon指数在整个香肠发酵周期并未呈现出规律性变化,整体值较高,总体变化呈上升趋势,在发酵21 d时出现最低值5.23;Simpson指数值越小,说明群落多样性越高,由表1可知,Simpson指数在整个香肠发酵周期变化情况为先下降,再上升,然后趋于稳定,发酵至14 d指数最小值为0.32,发酵28 d出现最大值为0.77。说明川味香肠在整个发酵周期中微生态群落多样性复杂,发酵时间不同,样品微生物结构存在明显差异。
2.2 川味发酵香肠发酵过程中微生态细菌群落结构分布及演变分析
2.2.1 门水平
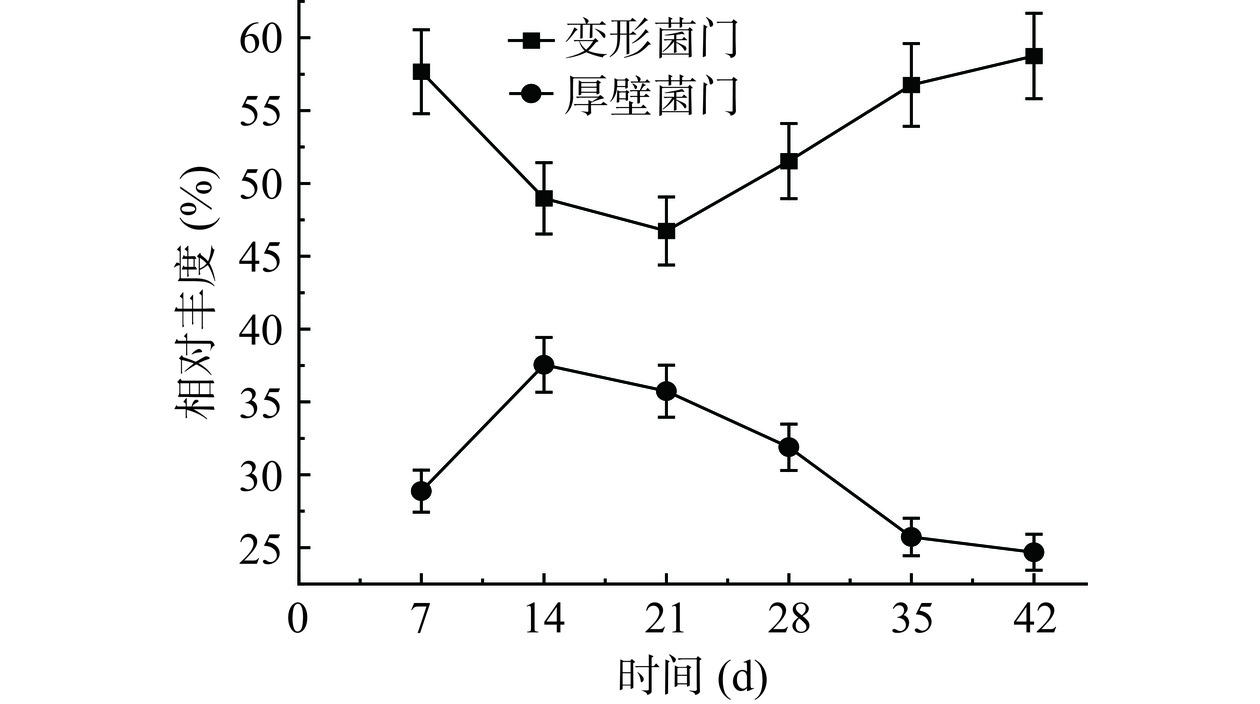
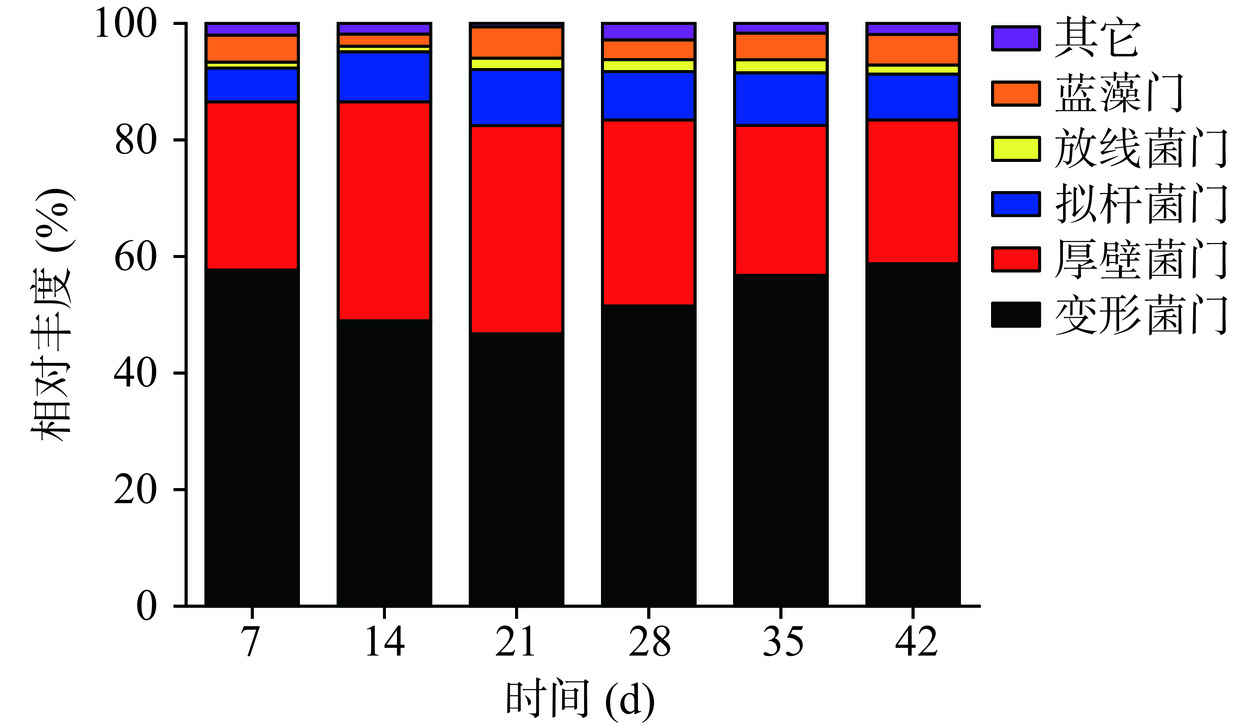
为探究分析川味发酵香肠样品微生物细菌物种组成及丰度,结合微生物群落α多样性分析作为群落结构分布及演变分析的前置条件[15]。根据OTU划分结果和分类信息,川味香肠发酵过程中微生态细菌鉴定出门水平群落结构分布如图1所示,在整个发酵周期中主要的门有以下5种,变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、蓝藻门(Cyanobacteria)、绿菌门(Chlorobiota)、弯曲菌门(Campylobacterota)。其中,变形杆菌门和厚壁菌门在整个发酵周期占优势地位(57.67%~46.74%、37.55%~24.68%),两类占比总和始终在80%以上。康峻[16]研究发现浅发酵香肠制作过程中品质特性与微生物多样性存在密切相关,其中变形菌门、厚壁菌门占优势地位,菌群结构的变化可能会引起香肠中游离氨基酸、脂肪酸,挥发性风味物质含量出现差异。田建军[17]采集了不同地区传统自然发酵和人为调控的发酵肉制品,利用高通量测序技术鉴定发现传统自然发酵肉制品中厚壁菌门(39%)、变形杆菌门(40%)、拟杆菌门(14%)为优势菌门,人为调控发酵肉制品中仅厚壁菌门(92%)为优势菌门。对变形杆菌门和厚壁菌门进行门演变分析,演变情况如图2所示,变形杆菌门在发酵过程中始终占据绝对优势地位,随着发酵的进行,变化趋势为先下降后上升,发酵至28 d,变形杆菌门占比已达50%,发酵进行至42 d时占比高达58.75%;厚壁菌门为次优势菌门,随着发酵的进行,呈现先上升后下降的变化规律,在发酵14 d时占比最高为37.55%。此外,拟杆菌门、放线菌门、蓝藻门这3种菌门也较为活跃,在发酵21 d占比一度超过15%,而后趋于相对稳定。这与王新惠等[18]对广味香肠自然发酵细菌演替规律研究结果一致。
![]() 图 1 川味发酵香肠发酵过程中微生态细菌在门水平的群落结构分布Figure 1. Community structure distribution of during the fermentationprocess of Sichuan fermented sausage microecological bacteria at phylum level
图 1 川味发酵香肠发酵过程中微生态细菌在门水平的群落结构分布Figure 1. Community structure distribution of during the fermentationprocess of Sichuan fermented sausage microecological bacteria at phylum level![]() 图 2 变形杆菌门和厚壁菌门在发酵周期的演变情况Figure 2. Evolution of Proteobacteria and Firmicutes during the fermentation cycle
图 2 变形杆菌门和厚壁菌门在发酵周期的演变情况Figure 2. Evolution of Proteobacteria and Firmicutes during the fermentation cycle2.2.2 属水平
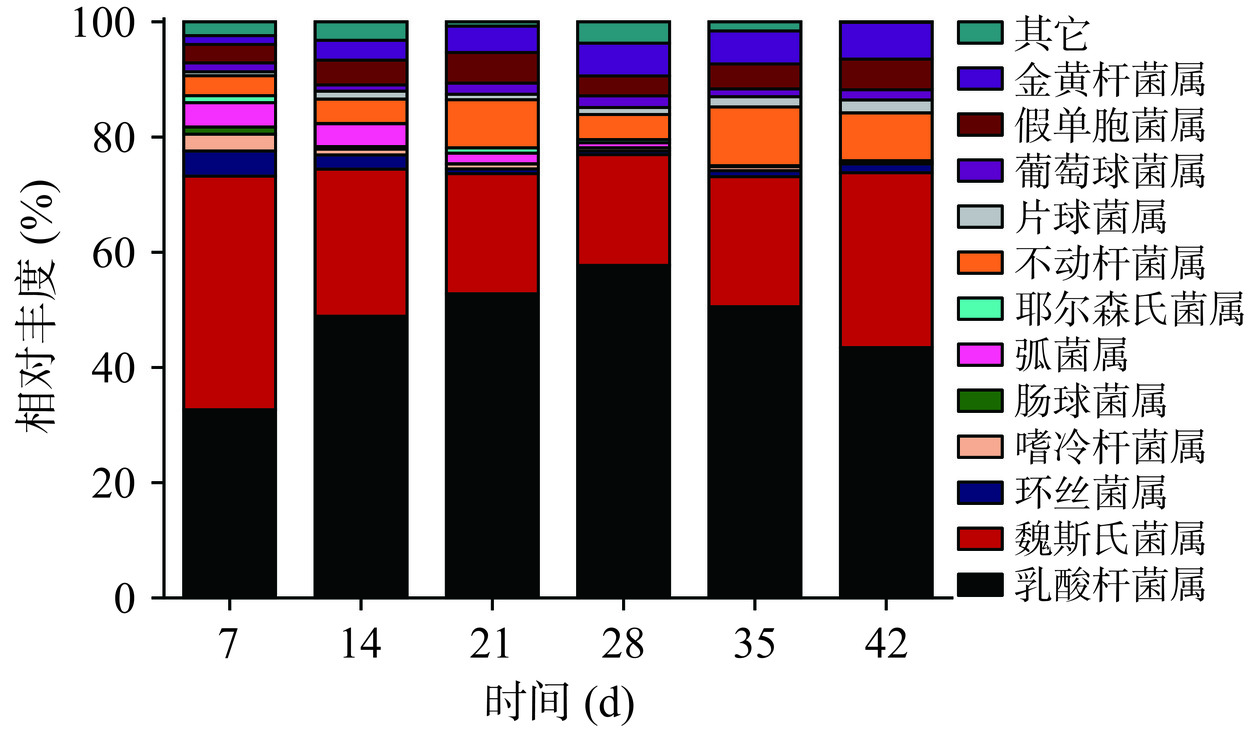
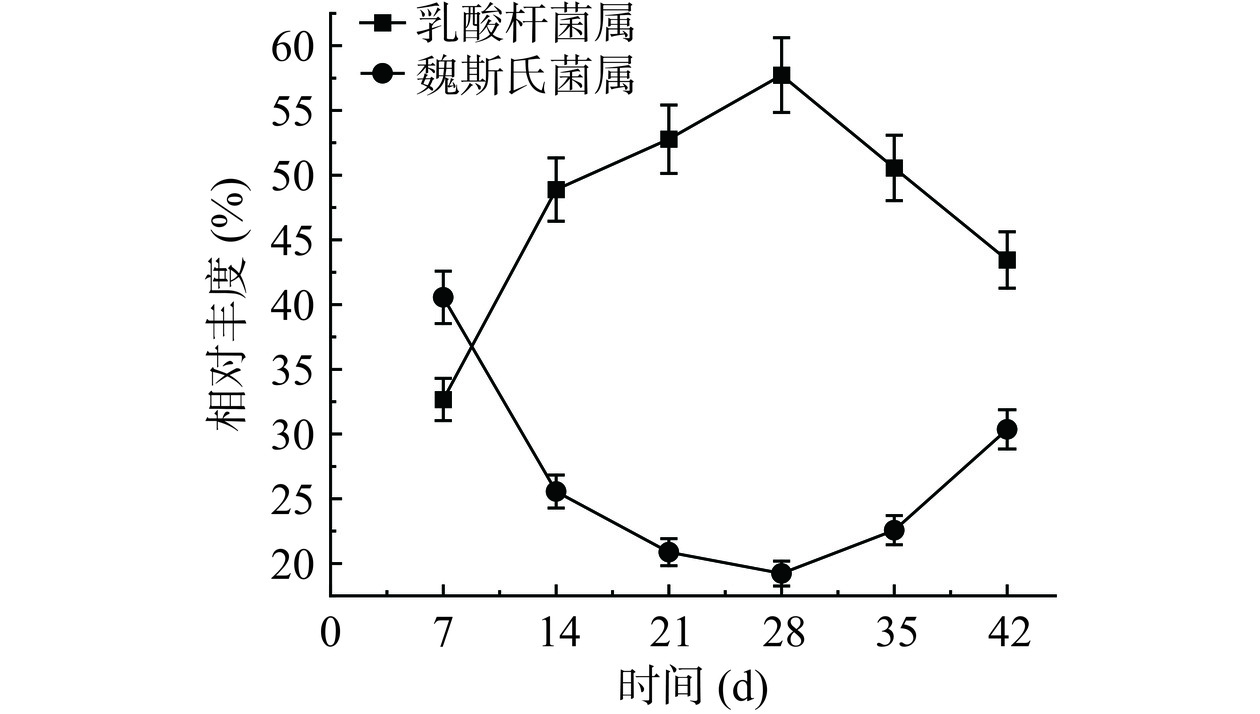
川味香肠发酵微生态细菌在属水平的群落结构分布如图3所示,在整个发酵周期中主要鉴定出乳酸杆菌属(Lactobacillus)、魏斯氏菌属(Weissella)、环丝菌属(Brochothrix)、嗜冷杆菌属(Psychrobacter)、耶尔森氏菌属(Yersinia)、不动杆菌属(Acinetobacter)、片球菌属(Pediococcus)、弧菌属(Vibrio)、葡萄球菌属(Staphylococcus)、肠球菌属(Enterococcus)、假单胞菌属(Pseudomonas)、金黄杆菌属(Chryseobacterium),以上12种属占比超过96%。其中,属于乳酸菌科的乳酸杆菌属和魏斯氏菌属在整个发酵过程中占优势地位,初始占比总和为61.96%,而后保持占比70%以上;王玥[19]在研究发酵香肠微生物多样性中鉴定出15个属的微生物菌群,其中占比最多的是乳酸杆菌属,其他的属有魏斯氏菌属、环丝菌属、葡萄球菌属等。赵改名等[20]在传统自然发酵的腊肉样品中共鉴定出83个细菌属,其中葡萄球菌属、嗜冷杆菌属、乳杆菌属、环丝菌属和魏斯氏菌属为主要优势细菌属。对乳酸杆菌属和魏斯氏菌属进行属演变分析,演变情况如图4所示,乳酸杆菌属是优势菌属,在发酵过程中一直处于主导地位,随着发酵时间持续升高,在发酵28 d达到极值,占比超过57%,而后略有下降,但占比始终在40%以上。魏斯氏菌属为次优势菌属,随着发酵时间先下降后上升,在发酵28 d达到极低值,占比仍超过19%。环丝菌属、嗜冷杆菌属、肠球菌属、弧菌属、耶尔森氏菌属和不动杆菌的初始占比总和为27.07%,呈快速下降,在发酵28 d达到极低值6.96%,而后变化趋于平稳,同张雅琳[21]研究结果相似,川味香肠发酵过程中的致病腐败菌属为环丝菌属,弧菌属、嗜冷杆菌属、肠球菌属、耶尔森氏菌属,并随着发酵时间的延长,占比先降低后升高。整个发酵周期葡萄球菌属、片球菌属、假单胞菌属、金黄杆菌属占比总和呈上升趋势,初始占比为6.97%,发酵结束时占比上升至15.71%。
![]() 图 3 川味发酵香肠发酵过程中微生态细菌在属水平的群落结构分布Figure 3. Community structure distribution during the fermentationprocess of Sichuan fermented sausage microecological bacteria at the genus level
图 3 川味发酵香肠发酵过程中微生态细菌在属水平的群落结构分布Figure 3. Community structure distribution during the fermentationprocess of Sichuan fermented sausage microecological bacteria at the genus level![]() 图 4 乳酸杆菌属和魏斯氏菌属在发酵周期的演变情况Figure 4. Evolution of Lactobacillus and Weisella during the fermentation cycle
图 4 乳酸杆菌属和魏斯氏菌属在发酵周期的演变情况Figure 4. Evolution of Lactobacillus and Weisella during the fermentation cycle2.3 川味发酵香肠发酵过程中的产酸情况分析
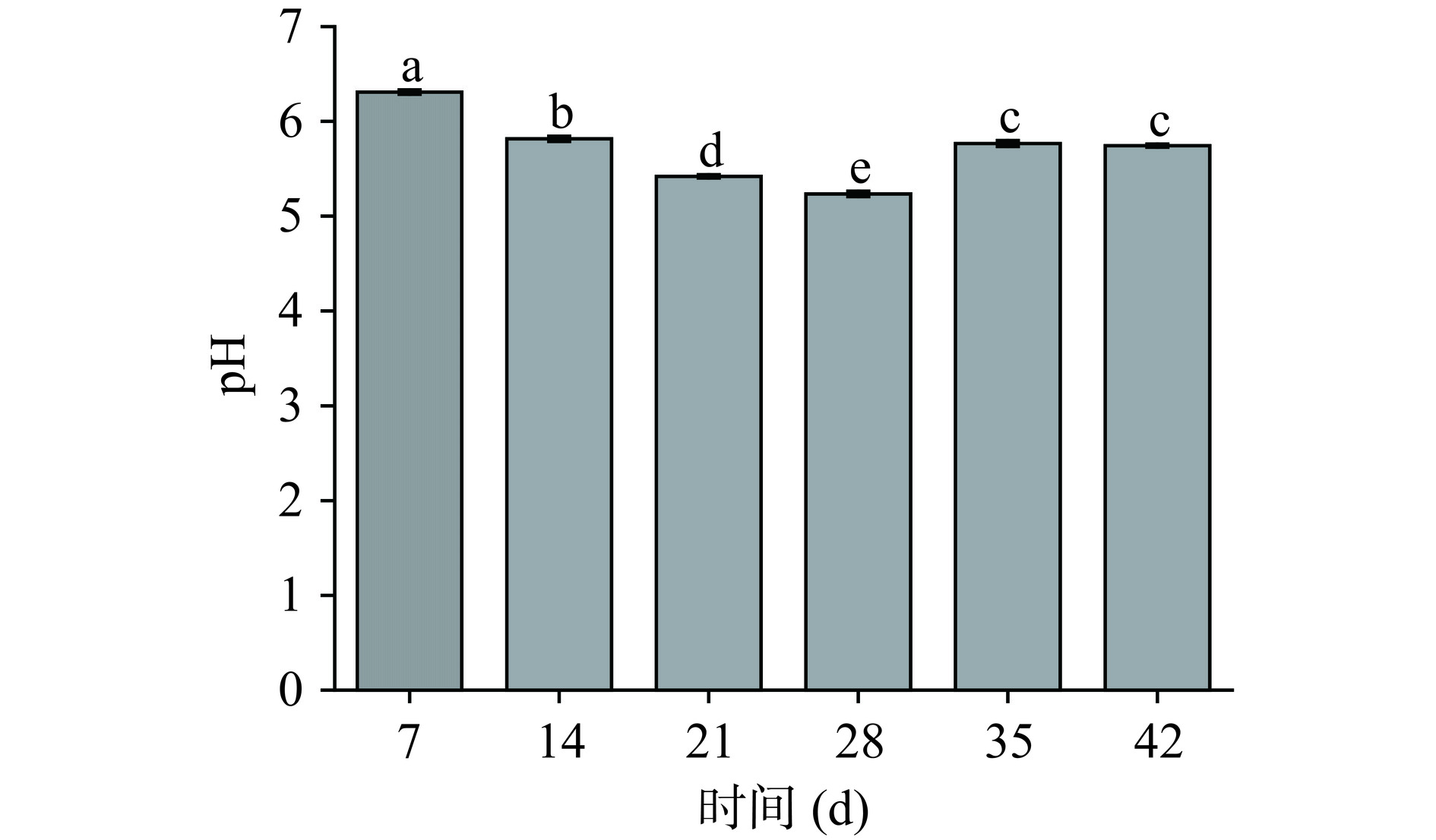
pH表示发酵香肠中游离氢离子的活度,是样品的有效酸度,能检测判断微生物产酸情况。其高低会影响发酵香肠的色泽、口感、风味[22],且较低pH能抑制有害微生物的生长。由图5可知,pH范围在5.235~6.309,发酵初期香肠pH相对最高,可能是因为初始发酵环境中代谢碳水化合物菌群较多,随发酵的进行,香肠pH 呈下降趋势,直至发酵28 d时pH最低,为5.235。香肠内的乳酸菌快速的利用其中原料肉中的组织酶和碳水化合物等发酵产酸,随着乳酸的积累,致使微生物生存环境的pH快速下降;发酵至35 d时,pH开始上升,表明随着发酵时间延长,蛋白质水解成游离氨基酸越多,降解出的碱性氨和胺类物质越多,同酸类物质中和,使pH回升,这与范文教等[23]研究结果相似,随着香肠原料肉蛋白水解成游离氨基酸并进一步降解为碱性的氨和胺类物质,同产生的乳酸中和作用,因而缓冲了pH的下降趋势。有文献表明,当pH较低时,影响了微生物代谢产生初级产物,进而造成次级代谢产物的差异变化,抑制腐败菌生长、延长食品货架期[24]。结合微生物α多样性分析,发现pH对香肠微生物结构变化有着一定影响,当随发酵至21 d时pH较低,OTU数和Simpson指数也较低分别为1568、0.56,且Shannon指数出现最低值5.23;且优势门属变形杆菌门和厚壁菌门、乳酸杆菌属和魏斯氏菌属在发酵中期消长变化明显,表明了川味香肠在发酵过程中的pH是微生物多样性显性指标之一,导致了肠体细菌菌群之间的相互竞争呈消长关系,影响了香肠内部细菌菌群的丰度和多样性的动态变化。
![]() 图 5 川味发酵香肠发酵过程中pH的变化注:不同小写字母表示差异显著(P<0.05)。Figure 5. Changes of pH during fermentation of Sichuan fermented sausage
图 5 川味发酵香肠发酵过程中pH的变化注:不同小写字母表示差异显著(P<0.05)。Figure 5. Changes of pH during fermentation of Sichuan fermented sausage2.4 川味发酵香肠发酵过程中非挥发物质分析
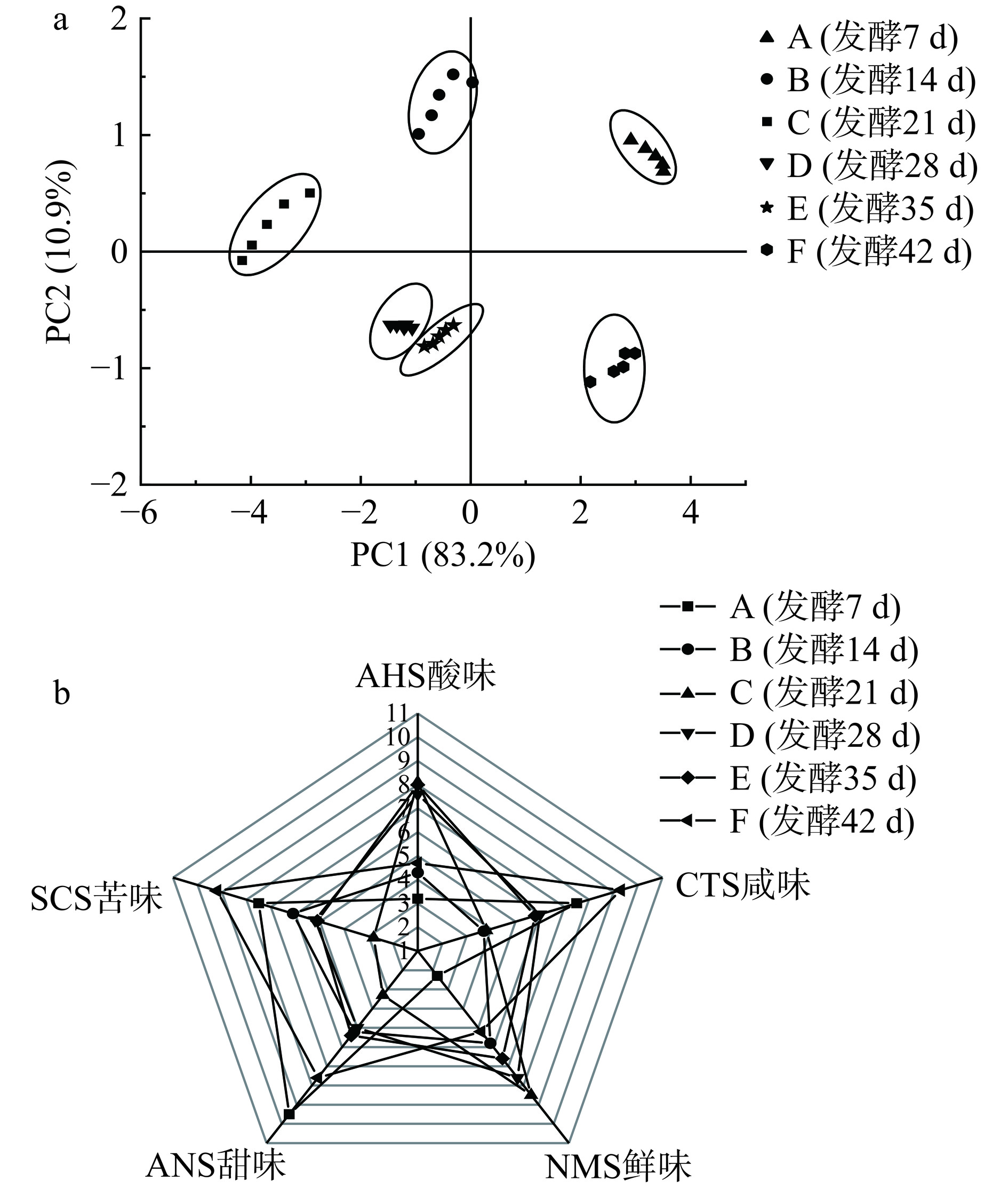
非挥发性物质属于滋味物质,主要依靠味觉器官感觉,经三叉神经传递到大脑而反应出味感。电子舌通过模拟人体的舌器官实现对呈味物质快速、准确的识别,传递为电信号进行表达。利用电子舌能够检测到人体舌头无法辨别的味道,将味觉信号转为电信号,将感官难以描述的差异性更加细化,反映检测样品的整体滋味信息,进一步分析样品之间存在的差异。为比较不同发酵时间的川味发酵香肠的整体滋味差异,对电子舌数据进行主成分分析,由图6a可知,PC1和PC2的累计方差贡献率分别为83.2%、10.9%,D组和E组两组间距较其他组更近,且分布在第三象限,说明该发酵阶段滋味成分差异较小;与其他实验组间距较大,说明发酵28 d与发酵35 d滋味差异显著。由图6b可知,在滋味方面,电子舌传感器对各组样品的响应值都有所差异,说明发酵过程中滋味变化较明显,D组和E组在滋味传感器轮廓大致相同,说明香肠发酵成熟期滋味差异不大,与PCA主成分分析结果一致。在酸性传感器AHS响应值随发酵时间延长而变化,先降低再缓慢回升,与pH变化一致;发酵香肠适当酸度,不仅抑制了有害杂菌,还为其他有益微生物提供生化反应的风味前体物质[25];直至发酵35~42 d,传感器SCS苦味响应值增高,在传感器NMS鲜味、ANS甜味响应值最高,是由于发酵过程中在微生物作用下大量蛋白质转化为游离氨基酸、核苷酸和呈苦味、鲜味肽的含量增多,相互协同增加鲜味[26],说明发酵时间延长会促进香肠中蛋白质代谢水解为滋味物质,且在发酵稳定期川味香肠样品鲜味十足,咸味适中,营养价值高、食用滋味最佳。这与CHENG等[27]研究结果相似,发酵成熟的香肠中呈味氨基酸丰富,占比约为75%。
![]() 图 6 川味发酵香肠发酵过程电子舌主成分分析图(a)及雷达图(b)Figure 6. Principal component analysis (a) and radar (b) diagrams of electronic tongue analysis during the fermentation process of Sichuan fermented sausage
图 6 川味发酵香肠发酵过程电子舌主成分分析图(a)及雷达图(b)Figure 6. Principal component analysis (a) and radar (b) diagrams of electronic tongue analysis during the fermentation process of Sichuan fermented sausage2.5 川味发酵香肠发酵过程中气味分析
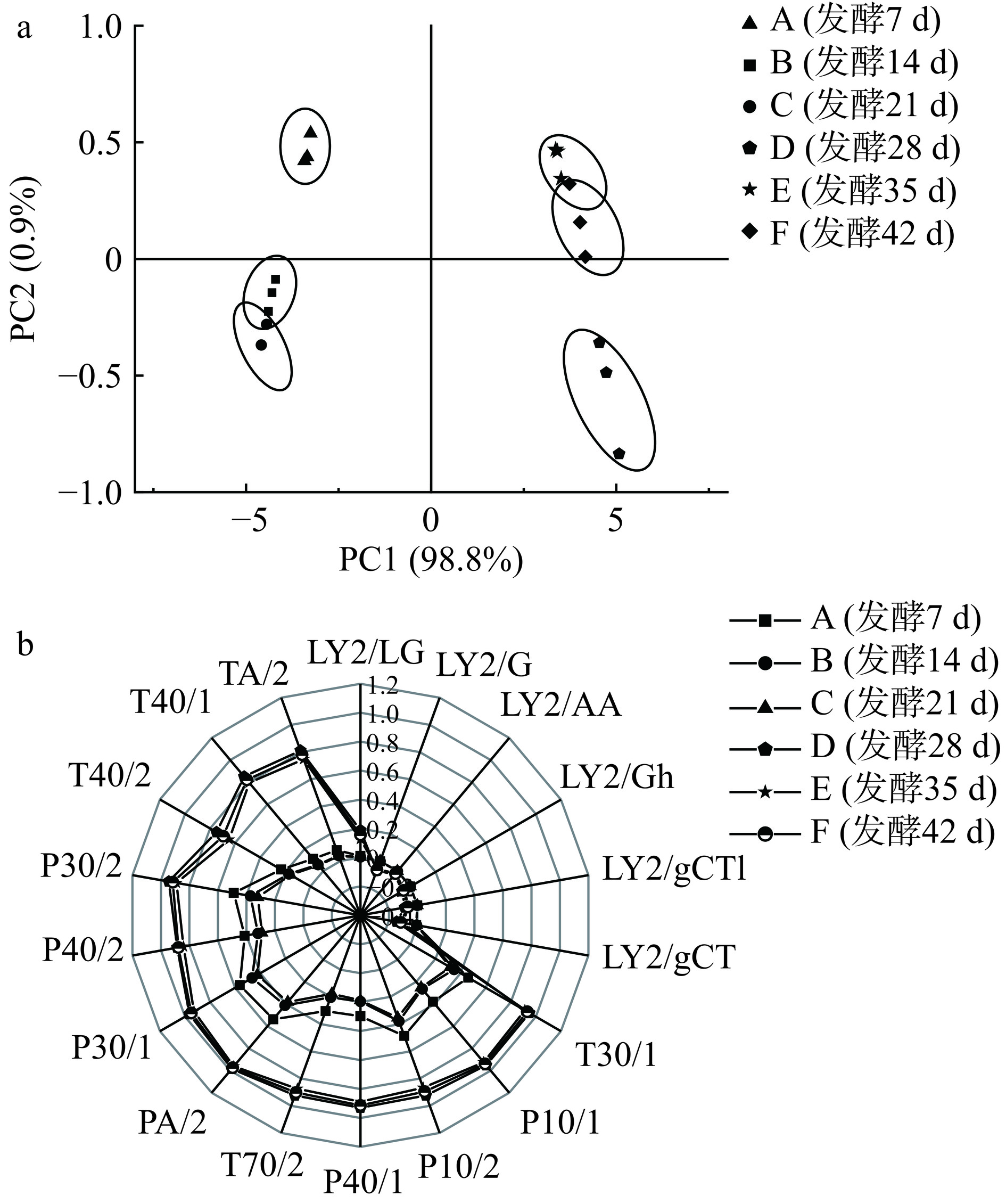
气味成分主要是由挥发性化合物组成,主要靠嗅觉细胞感受,同样经神经传递到大脑后反应为味感。电子鼻是一种模拟人体嗅觉器官的气敏传感器技术,由样品采集系统、检测系统、数据处理系统组成,能识别出待测样品散发的气味,转换成响应值进行分析[28]。通过主成分分析研究发酵过程中香肠整体风味变化,由图7a可知,PC1和PC2的累计方差贡献率分别为98.8%、0.9%,说明对香肠样品气味提取完整能反映出整体气味信息。A组和D组是独立于其他组,且分别在第二、四象限,说明两个发酵阶段香肠样品气味成分差异较大。B组和C组共同分布在第三象限且间距小,E组和F组同分布在第一象限且间距小,说明该四个时期B组和C组、E和F组挥发性风味物质较接近,因此根据挥发性风味物质的分布情况将整个发酵过程分为四个发酵阶段:发酵初期(7 d)、发酵中期(14~21 d)、发酵成熟期(28 d)、发酵稳定期(35~42 d)。此外,由图7b可知,在气味方面,发酵成熟期和发酵稳定期该两个发酵区间风味轮廓相似,说明香肠气味物质相对含量较接近;各组对LY2/G-LY2/CT风味轮廓较相似,其差异主要体现在其余传感器响应值随发酵时间延长而增高,如发酵成熟期响应值明显高于发酵中期;其中传感器P30/1、P30/2、T30/1响应值较高,说明对此类香肠中有机化合物和极性化合物醇、酮类含量更丰富,且影响发酵过程中香肠香气的主要成分为醇类物质;此外,各组发酵风味整体轮廓存在差异性变化,其中变化消长起伏、复杂多样,是因为微生物在发酵环境中能够产生多种蛋白酶和脂肪酶,促进香肠中的氨基酸和脂肪酸等风味前体物质的形成,使风味物质存在差异性变化[29]。
![]() 图 7 川味发酵香肠发酵过程电子鼻主成分分析图(a)及雷达图(b)Figure 7. Principal component analysis (a) and radar (b) diagrams of electronic nose analysis during the fermentation process of Sichuan fermented sausage
图 7 川味发酵香肠发酵过程电子鼻主成分分析图(a)及雷达图(b)Figure 7. Principal component analysis (a) and radar (b) diagrams of electronic nose analysis during the fermentation process of Sichuan fermented sausage2.6 川味发酵香肠发酵过程中挥发性风味物质分析
挥发性风味物质具有成分复杂、含量微小、不稳定、易挥发的特点。相关文献表明,发酵肉制品挥发性风味物质包括醛、酮、醇、酸、酯、杂环化合物和硫化合物等[30]。电子鼻能检测出样品整体气味信息,可实现样品的快速区分,但对具体挥发性风味物质定性、定量还有待深入;气相色谱-离子迁移谱法(GC-IMS)是一种高效、高灵敏度且高分辨率的风味检测技术,该技术基于不同的气相离子在电场中迁移速度的差异,实现化学离子物质进行样品分离鉴定,达到了对风味成分精确分析的目的[31]。因此采用GC-IMS研究分析不同发酵阶段的川味发酵香肠具体挥发性风味物质信息,为进一步探究发酵香肠的风味物质提供了可视化结果。
2.6.1 挥发性风味物质的GC-IMS谱图对比分析
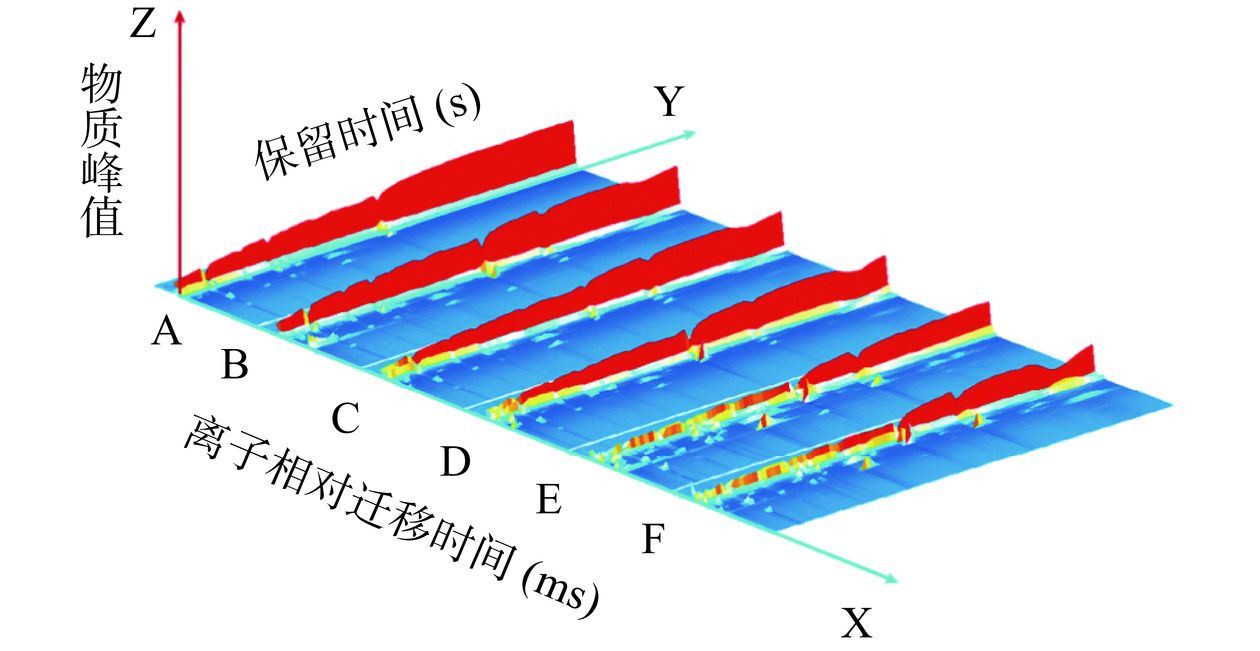
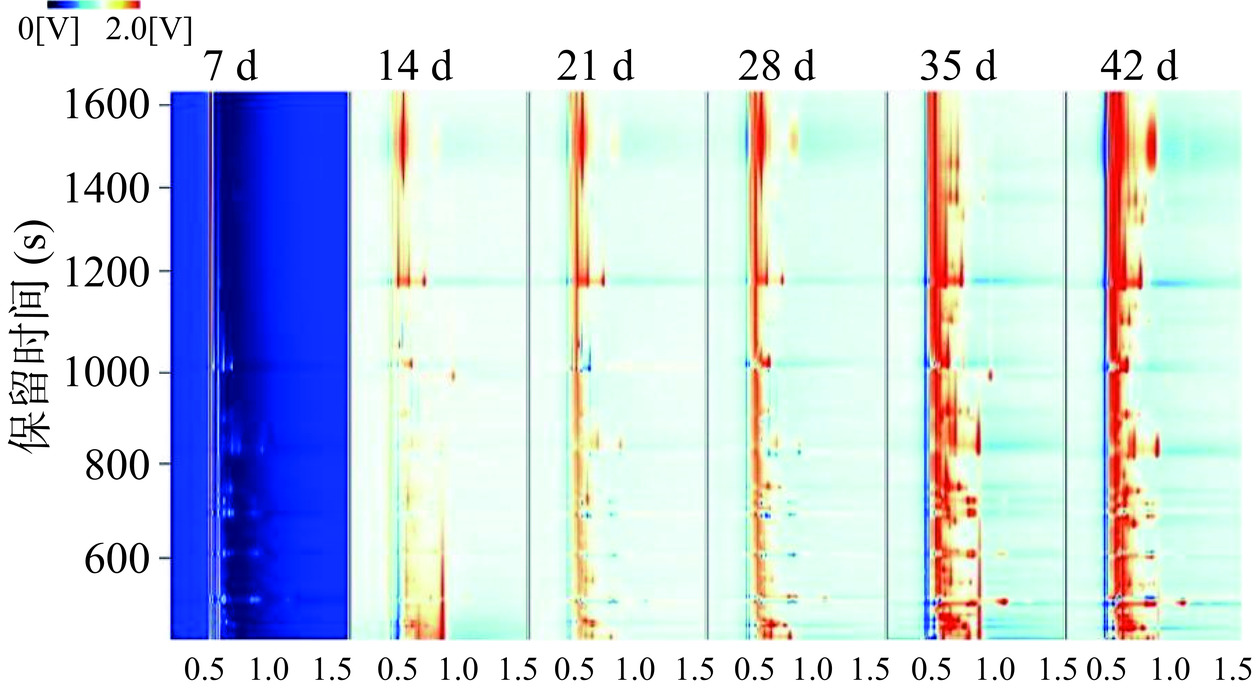
采用仪器配套的GC-IMS LAV分析软件中的Reporter程序对不同发酵时间的川味香肠挥发性风味物质进行三维对比,如图8所示,离子相对迁移时间设X轴,而气相色谱的保留时间则被确定为Y轴,同时,挥发性物质峰值的强度设为垂直空间Z轴,使得挥发性物质变化趋势可视性提高。由图8可看出,不同发酵时间组的川味香肠风味种类和浓度有着明显差异,存在一定变化规律。为了更清晰地观察川味香肠发酵过程中挥发性气味成分的差异,通过降维处理将三维图谱投影到二维平面上,同时采用差异对比模式,以A组样品图谱作参比样品,物质浓度相同颜色抵消为白色背景,浓度高于参比的物质标记为红色,浓度低的为蓝色[32],结果见图9。由图9可知,与样品A相比,经过不同发酵时间香肠样品在挥发性风味物质的种类与相对含量上均呈现出显著的增长,表明在发酵过程中有新的风味物质产生。其中发酵中期挥发性风味物质浓度迅速升高,至发酵成熟期浓度最高,发酵稳定期物质浓度稳定不变,表明川味香肠中的挥发性风味物质随发酵时间延长而显著增加。
![]() 图 8 不同发酵时间组香肠GC-IMS三维谱图Figure 8. Sausage GC-IMS 3D topographic map for different fermentation time groups
图 8 不同发酵时间组香肠GC-IMS三维谱图Figure 8. Sausage GC-IMS 3D topographic map for different fermentation time groups![]() 图 9 不同发酵时间组香肠GC-IMS二维谱图Figure 9. Sausage GC-IMS two dimensional map for different fermentation time groups
图 9 不同发酵时间组香肠GC-IMS二维谱图Figure 9. Sausage GC-IMS two dimensional map for different fermentation time groups2.6.2 挥发性风味物质的GC-IMS定性分析
通过GC-IMS仪器配套的LS软件和系统内置数据库对不同发酵时间的川味香肠挥发性风味物质定性分析,如表2所示,共鉴定出52种挥发性风味物质,其中包括酯类13种(二聚体1个)、醇类10种(二聚体2个)、醛类7种(1个)、萜烯类9种(二聚体3个)、酮类3种、烷烃类2种(二聚体1个)、吡嗪2种、噻唑2种、硫化物2种、胺类1种,各挥发性物质基本信息如下表2所示。
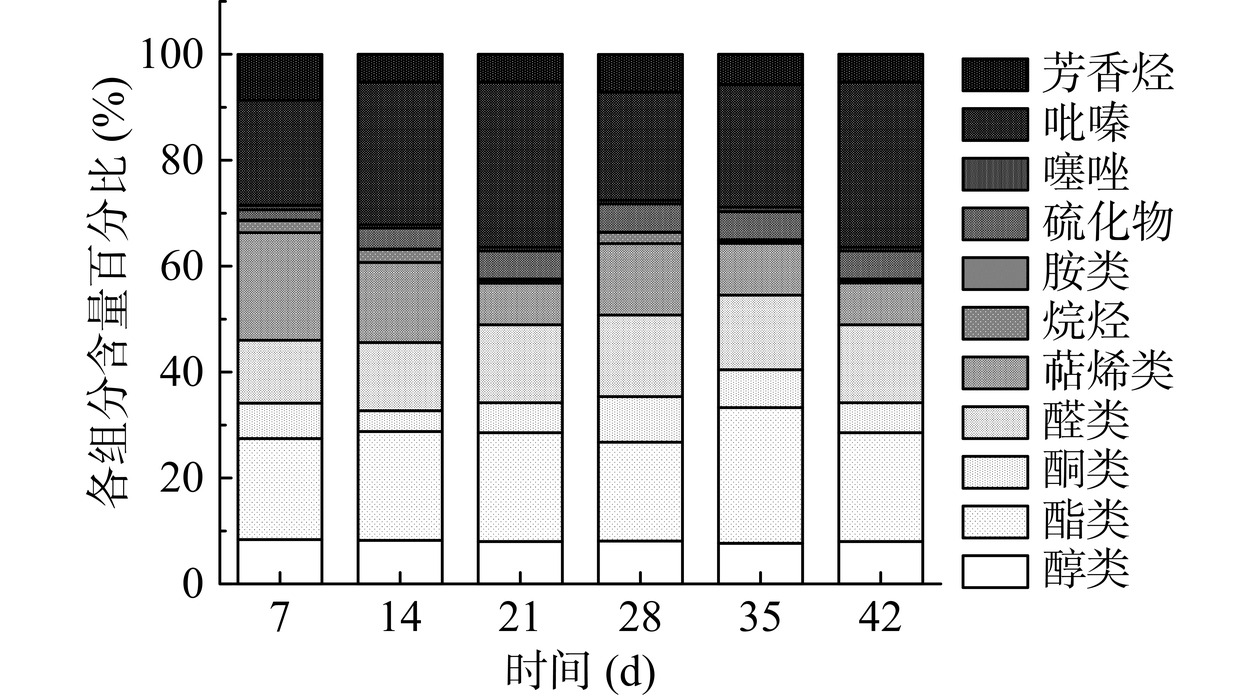
表 2 基于气相色谱-离子迁移鉴定川味发酵香肠挥发性成分Table 2. Identification fied volatile components of Sichuan fermented sausages based on GC-IMS类别 化合物 CAS号# 保留指数 保留时间(s) 迁移时间(ms) 醇类 二丙酮醇(2-Hydroxy-2-methyl-4-pentanone) C123422 1358.2 1207.752 1.53264 2-庚醇(2-Heptanol) C543497 1331.2 1127.021 1.37566 异戊醇(1-Butanol, 3-methyl) C123513 1214.3 755.924 1.50193 正丁醇(1-Butanol) C71363 1137.7 559.287 1.38949 2-辛醇(2-Octanol) C123966 1418.8 1389.637 1.86536 异丁醇(1-Propanol, 2-methyl-) C78831 1077 442.018 1.36987 乙醇单体(Ethanol-M) C64175 922.3 300.818 1.04287 乙醇二聚体(Ethanol-D) C64175 921.5 300.394 1.12915 1-戊烯-3-醇(1-Penten-3-ol) C616251 1199.6 708.061 1.34308 2-丁醇(2-Butanol) C78922 1045.5 403.585 1.31695 酯类 辛酸乙酯(Ethyl octanoate) C106321 1419.1 1390.363 1.47109 己酸乙酯二聚体(Ethyl caproate-D) C123660 1243.6 851.268 1.79796 己酸乙酯单体(Ethyl caproate-M) C123660 1242.7 848.086 1.34778 甲酸异戊酯(Isoamyl formate) C110452 1072.3 436.226 1.26483 异戊酸乙酯单体(Butanoicac id 3-methyl ethyl ester-M) C108645 1058 418.865 1.24865 异戊酸乙酯二聚体(Butanoicac id 3-methyl ethyl ester-D) C108645 1057 417.569 1.6541 丁酸乙酯(Butanoic acid ethyl ester) C105544 1047.3 405.765 1.55688 异丁酸乙酯(Ethyl 2-methy lpropionate) C97621 981.5 334.868 1.5676 丙酸乙酯(Ethyl propanoate) C105373 967.6 326.842 1.45361 2-丁烯酸己酯(Hexyl 2-butenoate) C19089920 1248 865.323 1.45154 戊酸乙酯(Ethyl pentanoate) C539822 1148.9 584.817 1.26987 乙酸甲酯(Methyl acetate) C79209 851.1 259.889 1.19629 酮类 仲辛酮(2-Octanone) C111137 1299.1 1030.654 1.32954 α-侧柏酮(α-Thujone) C546805 1419.7 1392.187 1.37168 3-羟基-2-丁酮(2-Butanone, 3-hydroxy) C513860 1266.2 924.656 1.33186 醛类 正丁醛单体(Butanal-M) C123728 853.5 261.293 1.118 苯甲醛(Benzaldehyde) C100527 1518.3 1687.872 1.15767 正丁醛二聚体(Butanal-D) C123728 849.5 258.966 1.28286 丙烯醛(Acrolein) C107028 842.4 254.894 1.06645 丙醛(Propanal) C123386 840 253.524 1.14891 异丁醛(2-Methyl propanal) C78842 821.7 243.035 1.08961 正己醛(1-Hexanal) C66251 1101.3 476.317 1.26798 萜烯类 β-蒎烯单体(β-Pinene-M) C127913 1121.6 522.457 1.22316 β-蒎烯二聚体(β-Pinene-D) C127913 1119.5 517.72 1.6409 (+)-柠檬烯单体((+)-Limonene-M) C138863 1199.6 708.124 1.21957 β-月桂烯单体(β-Myrcene-M) C123353 1162.6 616.176 1.22329 β-月桂烯二聚体(β-Myrcene-D) C123353 1161.9 614.452 1.64633 罗勒烯((Z)-Ocimene) C470826 1215.7 760.281 1.31781 γ-松油烯(γ-Terpinene) C99854 1249.6 870.59 1.21383 (+)-柠檬烯二聚体((+)-Limonene-D) C138863 1199.5 707.736 1.29659 α-松油烯(α-Terpinene) C99865 1180.2 656.311 1.21549 烷烃 1,1-二乙氧基乙烷单体(1,1-Diethoxy ethane-M) C105577 885.1 279.445 1.04309 1,1-二乙氧基乙烷二聚体(1,1-Diethoxy ethane-D) C105577 889.1 281.729 1.12655 胺类 二丁胺(Dibutylamine) C111922 1072 435.916 1.73091 硫化物 烯丙基硫化物(Allyl propyl sulfide) C27817670 1094 462.752 1.37291 二甲基三硫(Dimethyl trisulfide) C3658808 1362.2 1219.901 1.30366 噻唑 噻唑(Thiazole) C288471 1263 914.13 1.25028 2,4,5-三甲基噻唑(2,4,5-Trimethylthiazole) C13623115 1403.6 1344.092 1.56922 吡嗪 2,3-二乙基吡嗪(2,3-Diethylpyrazine) C15707241 1456.8 1503.541 1.71097 2,4,5-三甲基噻唑(2,4,5-Trimethylthiazole) C13623115 1403.6 1344.092 1.56922 芳香烃 丁苯(Benzene, butyl-) C104518 1306.1 1051.558 1.21373 注:“-M”表示单体,“-D”表示二聚体。 研究文献表明,60.5%的风味物质来源于各种香料,剩下的部分是来自于微生物作用下碳水化合物、脂肪氧化、氨基酸分解代谢等方式。进一步分析香肠发酵过程中挥发性风味物质变化,采用归一法对鉴定出不同类型的化合物进行半定量分析。如图10所示,整体而言,随发酵的进行,醇类、萜烯类、噻唑物质浓度减少,酯类、吡嗪类、醛类浓度增多;发酵稳定期,酯类相对其他挥发性风味物质含量较多,如乙酯类物质为香肠贡献出特殊水果香、坚果香,而该类物质主要是由微生物酯酶催化酯化反应产生的挥发性物质[33]。醇类来源较广泛,主要由微生物碳水化合物的代谢产生,如乙醇、2-丁醇等,部分醇来自于脂肪氧化;醛类主要是通过脂肪氧化和微生物代谢氨基酸和氨基酸的Strecker降解反应;酮类来自于香肠原料肉中脂质氧化分解,产生3-羟基-2-丁酮、α-侧柏酮、仲辛酮,贡献独特的松香和草木的清香[34]。吡嗪化合物通常由美拉德反应或氨基酸分解形成,阈值较低,可赋予肉类强烈香气[35]。由图10分析可知,发酵香肠的挥发性风味组成中酯类(21.52%)、吡嗪(23.53%)、醛类(14.01%)、醇类(8.31%)相对含量最高,对香肠风味贡献有着重要作用。其中萜烯类相对含量从20.29%降至7.87%,萜烯类主要是由制作香肠时所添加辅料花椒、胡椒、辣椒等的挥发性成分形成[36],说明随发酵进行到发酵成熟期香肠中香辛料的辛辣刺激味减少,结合其他风味物质,在香肠发酵稳定期呈现出风味丰富、温和醇香的特点。
![]() 图 10 川味发酵香肠发酵过程中各类挥发性物质相对含量Figure 10. Relative content of various volatile substances during the fermentation process of Sichuan fermented sausage
图 10 川味发酵香肠发酵过程中各类挥发性物质相对含量Figure 10. Relative content of various volatile substances during the fermentation process of Sichuan fermented sausage2.6.3 挥发性风味物质的GC-IMS指纹图谱分析
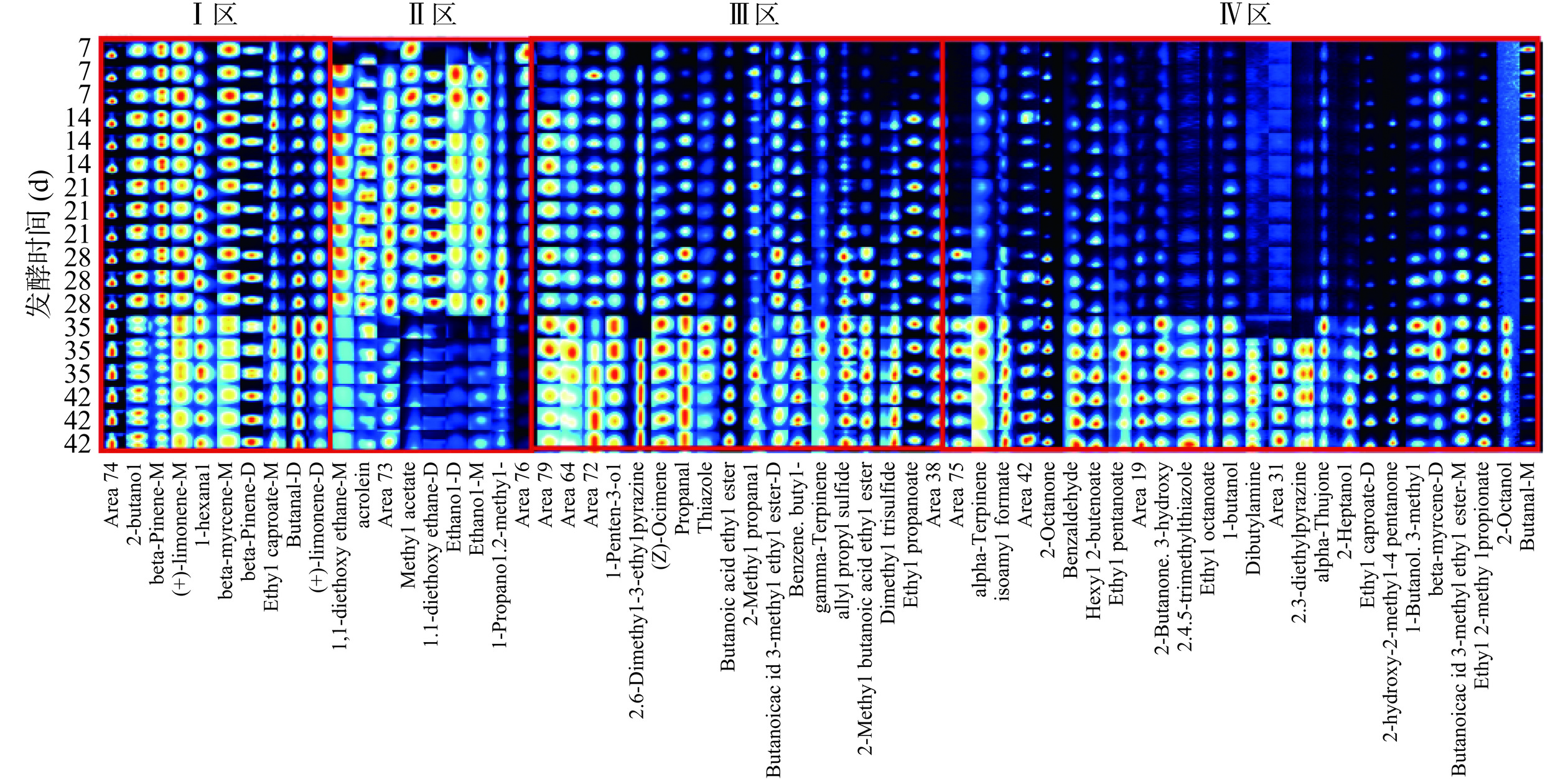
为了更加直观地分析香肠发酵过程挥发性风味物质具体变化规律,利用仪器配套的GC-IMS LAV软件中的Gallery Plot程序进行指纹图谱绘制。每一行均代表一个特定样品中不同挥发性物质的信号峰,而每一列则反映同一挥发性物质在不同样品中的信号表现。背景色为蓝色,点的颜色和面积则用以表示物质的相对含量,其中点越深面积越大表示该物质浓度含量越高[37]。不同发酵时间的川味香肠特征风味GC-IMS指纹图谱见图11。根据图11可看出香肠样品挥发性风味种类和含量差距较大,将图谱划分为四个不同区Ⅰ区、Ⅱ区、Ⅲ区、Ⅳ区进行分析可以看出,随发酵时间延长,Ⅰ区、Ⅲ区、Ⅳ区物质浓度增加,Ⅱ区物质浓度降低。
![]() 图 11 不同发酵时间组香肠特征风味GC-IMS指纹谱图Figure 11. GC-IMS fingerprint of sausage flavor characteristics in different fermentation time groups
图 11 不同发酵时间组香肠特征风味GC-IMS指纹谱图Figure 11. GC-IMS fingerprint of sausage flavor characteristics in different fermentation time groups其中Ⅰ区成分在香肠不同发酵阶段均检出且相对含量较高的风味物质有2-丁醇、正己醛、β-蒎烯、β-月桂烯、柠檬烯等,为发酵香肠贡献出柠檬香气、清新酒香以及香辛料的特殊风味[38]。发酵前期、中期(7~21 d),Ⅱ区成分浓度迅速增加,如异丁酸、乙酸甲酯、乙醇单双分子、丙烯醛、1,1-二乙氧基乙烷单双分子等;从图11可知,发酵前期和中期川味香肠的特征风味物质相似,由柠檬烯、β-蒎烯(M)、乙醇、β-月桂烯、1,1-二乙氧基乙烷(M);发酵成熟期(28 d),Ⅱ区成分浓度相对稳定,Ⅲ区成分浓度呈缓慢增长趋势,如丙酸乙酯、丙醛、异丁醛、1-戊烯-3-醇、γ-松油烯、噻唑、硫化物等;直至发酵稳定期(35~42 d),Ⅱ区成分浓度骤降,异丁酸、丙烯醛产生的辛辣刺激性显著减少,赋予香肠水果香、酒香的乙酸甲酯、乙醇等物质相对减少,由于发酵成熟期乳酸菌属减少、魏斯氏菌属增多,杂菌减少,以及其他优势菌属相互协同对氨基酸、脂质氧化代谢产生一定影响[39];同时,Ⅲ区和Ⅳ区各成分浓度迅速增长,主要以酯类、醛类、酮类、吡嗪类等合成为主,其中丁酸乙酯、丙醛、正丁醛(D)、2-乙基-3,5-二甲基吡嗪、苯甲醛、罗勒烯是发酵稳定期香肠样品主要特征物质,苯甲醛主要源于微生物对苯丙氨酸的代谢作用[40],辛酸乙酯的相对含量从0.51%增加到4.11%,α-侧柏酮的相对含量从0.42%增加到13.56%,为香肠贡献出特殊酒香、薄荷香。综上所述,表明随发酵时间延长,川味香肠挥发性风味成分种类和浓度存在差异,具有一定消长规律,发酵前期、中期挥发性风味物质快速增长,发酵至35 d时刺激性风味物质显著降低,发酵稳定期香肠特征风味种类和含量相对增多,主要以酯类、吡嗪、醛类为特征风味物质。
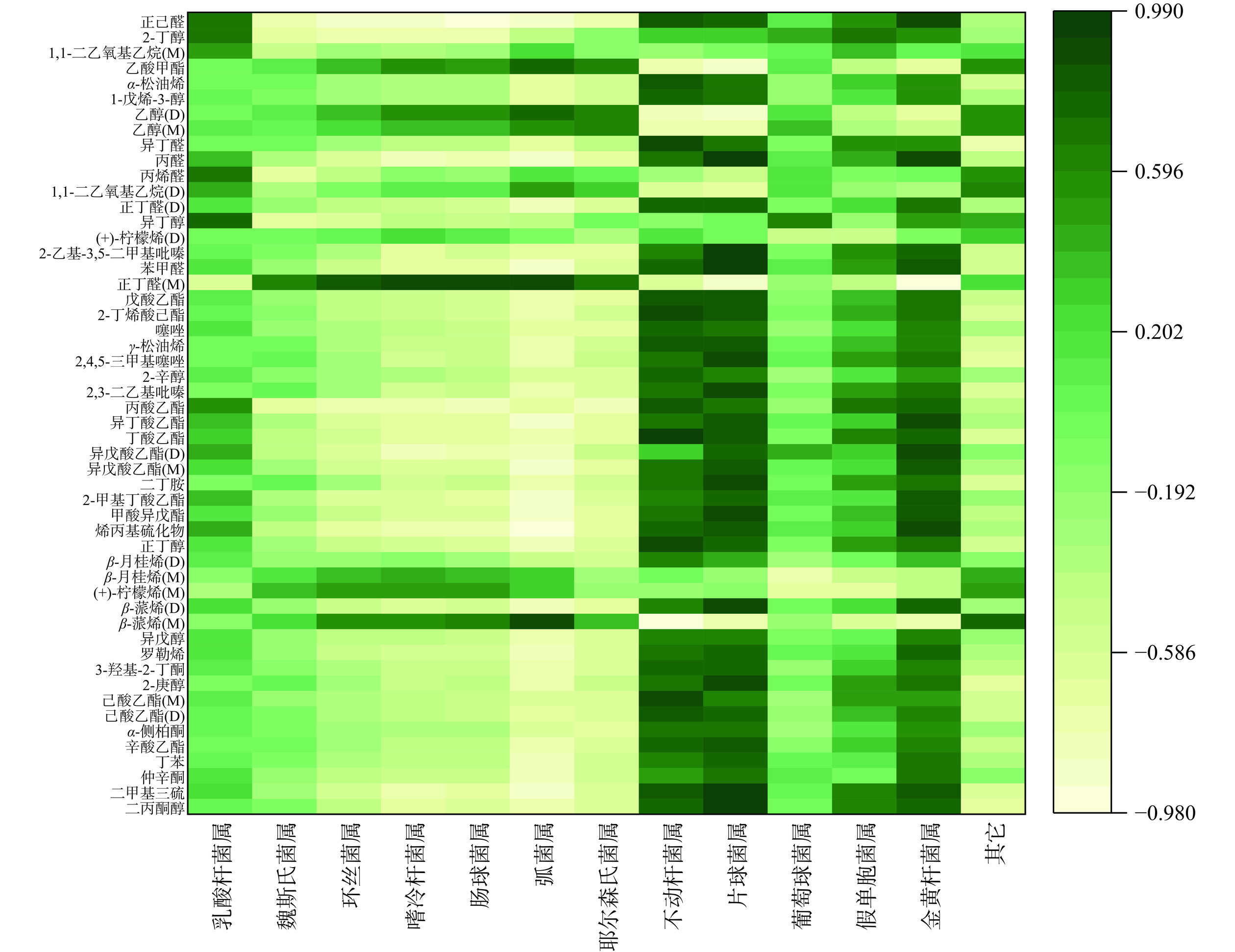
2.7 川味发酵香肠发酵过程中微生物与挥发性风味物质相关性分析
通过高通量测序技术筛选川味香肠中优势菌群,结合Pearson相关性,将筛选出的12种优势菌属与52种特征挥发性风味物质进行深入相关性分析,采用相关性热图直观表示,如图12所示,热图颜色越深,相关性系数越大。乳酸杆菌属与醛类、醇类呈强相关,如正乙醛(r=0.698)、2-丁醇(r=0.705)、丙烯醛(r=0.724)、异丁醇(r=0.761)、丙酸乙酯(r=0.566)呈中等相关,说明该菌属对香肠风味有着重要作用,同刘英丽等[41]研究所述一致;魏斯氏菌属对正丁醛(r=0.603)呈显著正相关,对正己醛(r=−0.756)呈显著负相关,对多数醇类和酯类呈显著负相关,如2-丁醇(r=−0.709)、异丁醇(r=−0.677)、丙酸乙酯(r=-0.656)、异戊酸乙酯(r=0.396);乳酸杆菌属和魏斯氏菌属在整个发酵过程中占优势地位,能产生大量蛋白酶,使蛋白质分解为游离氨基酸,进一步在酶类物质的催化作用下生成酸、杂环类和核苷酸等风味前提物质赋予发酵制品的营养价值和风味[42−43]。葡萄球菌属与醇类、酯类等主要风味物质中的异丁醇(r=0.640)、2-丁醇(r=0.456)、异戊酸乙酯(r=0.401)呈正相关,与魏斯氏菌属呈负相关;乳酸杆菌属、不动杆菌属、片球菌属、假单胞菌属、金黄杆菌属五类菌属与37种风味物质呈正相关,其中酯类和酮类为主要物质,且变化趋势相同,说明川味香肠中挥发性风味与该五类菌属密切相关,对风味影响最大。与之相反作用的菌属环丝菌属、嗜冷杆菌属、肠球菌属、弧菌属、耶尔森氏菌属均与乙酸甲酯、乙醇、正丁醛、β-月桂烯(M)(r>0.70)呈显著正相关,与其他风味成分均呈负相关,风味贡献较小,其中环丝菌属和嗜冷杆菌属对月桂烯、β-蒎烯等萜烯类挥发物质贡献特殊的川式香辣风味。此外,非优势菌属对醇类、酯类、烷烃有风味贡献,如乙酸甲酯、乙醇单体二聚体、丙烯醛、1,1-二氧基乙烷(D)、异丁醇、β-月桂烯、β-蒎烯均呈正相关。各优势菌属在发酵过程中相互协同,发挥作用,存在一定消长规律。综上所述,表明川味发酵香肠的挥发性风味物质与优势菌群的结构变化有关,微生物多样性将影响产品风味的丰富性,因此可利用微生物调控发酵香肠的风味品质和产品安全性。
![]() 图 12 属水平优势菌群与挥发性风味物质相关关系分析热图Figure 12. Heatmap of correlation between dominant flora and volatile flavor substances
图 12 属水平优势菌群与挥发性风味物质相关关系分析热图Figure 12. Heatmap of correlation between dominant flora and volatile flavor substances结合川味发酵香肠发酵过程中微生态演变情况,乳酸杆菌属相对含量较多,产酸降低产品的pH,抑制有害杂菌的生长,与醛类、醇类、酯类等风味的合成相关,与魏斯氏菌属相互协同产生挥发性物质,改善产品风味和质地。葡萄球菌属含量较低,但该类菌属的超氧化物歧化酶和过氧化氢酶活性较强,能够明显减缓发酵香肠制品的过度氧化,贡献香肠特有的奶油香味[44]。不动杆菌属、片球菌属、假单胞菌属、金黄杆菌属均参与蛋白质、脂质代谢且与重要风味物质的形成联系紧密[16],可抑制自然发酵香肠中其他有害微生物生长与不良风味的合成,提高香肠制品安全性。赵银峰[45]将戊糖片球菌和肉葡萄球菌复配接种在川味香肠中,1-辛烯-3-醇、3-甲基-1-丁醇和己酸乙烯酯等风味物质的浓度显著升高,提高了制品风味品质多样性;牛茵等[46]研究发现葡萄球菌属、明串珠菌属、假单胞菌属、嗜冷杆菌属、弧菌属等菌群动态变化参与挥发性物质生成,如氨基酸、脂肪酸、碳水化合物代谢途径,对挥发性风味物质有着积极作用。因此将乳酸杆菌属、葡萄球菌属、片球菌属、环丝菌属、嗜冷杆菌属作发酵剂接种于香肠,同其他优势菌群协同、竞争参与蛋白质、脂质及碳水化合物代谢分解,生成游离氨基酸和系列小分子肽、核苷酸以及醇、酯、醛类、酮类等风味物质,对于提高香肠的质量、营养价值和风味均有着重要意义。
3. 结论
本研究发现川味发酵香肠微生物菌群结构与风味物质存在显著相关性,结果表明:微生物α-多样性分析显示香肠发酵周期细菌群落复杂,共检测出5种门,12种菌属,门水平上,变形菌门和厚壁菌门占绝对优势;属水平上,乳酸杆菌属和魏斯氏菌属占绝对优势。在香肠发酵的整个过程中共检测出52种挥发性物质,发酵前期和发酵中期川味香肠的特征风味物质为柠檬烯、β-蒎烯(M)、乙醇、β-月桂烯、1,1-二乙氧基乙烷(M);发酵至35 d时,产生刺激性风味物质的异丁酸、丙烯醛含量显著降低,发酵稳定期香肠主要以酯类、吡嗪、醛类为主要特征风味物质。Pearson相关性分析显示乳酸杆菌属、不动杆菌属、片球菌属、假单胞菌属、金黄杆菌属等5种优势菌属对37种风味物质呈正相关,乳酸杆菌属与醛类、醇类、酯类呈正相关(r>0.6),Weissella与醇类和酯类呈负相关(r<−0.4)。此外,魏斯氏菌属、葡萄球菌属与正丁醛、正己醛、2-丁醇、异丁醇、丙酸乙酯、异戊酸乙酯等挥发性风味物质的形成有着紧密联系。因此可将其上述优势菌属作发酵剂调控发酵香肠的风味品质,为研发推广改善发酵香肠风味品质的绿色高效微生物发酵剂提供依据。
-
![]()
图 1 川味发酵香肠发酵过程中微生态细菌在门水平的群落结构分布
Figure 1. Community structure distribution of during the fermentationprocess of Sichuan fermented sausage microecological bacteria at phylum level
![]()
图 2 变形杆菌门和厚壁菌门在发酵周期的演变情况
Figure 2. Evolution of Proteobacteria and Firmicutes during the fermentation cycle
![]()
图 3 川味发酵香肠发酵过程中微生态细菌在属水平的群落结构分布
Figure 3. Community structure distribution during the fermentationprocess of Sichuan fermented sausage microecological bacteria at the genus level
![]()
图 4 乳酸杆菌属和魏斯氏菌属在发酵周期的演变情况
Figure 4. Evolution of Lactobacillus and Weisella during the fermentation cycle
![]()
图 5 川味发酵香肠发酵过程中pH的变化
注:不同小写字母表示差异显著(P<0.05)。
Figure 5. Changes of pH during fermentation of Sichuan fermented sausage
![]()
图 6 川味发酵香肠发酵过程电子舌主成分分析图(a)及雷达图(b)
Figure 6. Principal component analysis (a) and radar (b) diagrams of electronic tongue analysis during the fermentation process of Sichuan fermented sausage
![]()
图 7 川味发酵香肠发酵过程电子鼻主成分分析图(a)及雷达图(b)
Figure 7. Principal component analysis (a) and radar (b) diagrams of electronic nose analysis during the fermentation process of Sichuan fermented sausage
![]()
图 8 不同发酵时间组香肠GC-IMS三维谱图
Figure 8. Sausage GC-IMS 3D topographic map for different fermentation time groups
![]()
图 9 不同发酵时间组香肠GC-IMS二维谱图
Figure 9. Sausage GC-IMS two dimensional map for different fermentation time groups
![]()
图 10 川味发酵香肠发酵过程中各类挥发性物质相对含量
Figure 10. Relative content of various volatile substances during the fermentation process of Sichuan fermented sausage
![]()
图 11 不同发酵时间组香肠特征风味GC-IMS指纹谱图
Figure 11. GC-IMS fingerprint of sausage flavor characteristics in different fermentation time groups
![]()
图 12 属水平优势菌群与挥发性风味物质相关关系分析热图
Figure 12. Heatmap of correlation between dominant flora and volatile flavor substances
表 1 川味香肠发酵微生态细菌群落α多样性分析
Table 1 Alpha diversity analysis of fermented microecological bacterial communities in Sichuan sausage
指标 发酵时间(d) 7 14 21 28 35 42 GC率 0.98 0.97 0.98 1 0.99 1 OTU数 1029 1356 1568 1687 1765 1852 ACE指数 2865 3321 3542 3589 3615 3752 Chao1指数 2985 3102 2895 3365 3125 3621 Shannon指数 6.56 6.12 5.23 6.98 5.98 7.01 Simpson指数 0.43 0.32 0.56 0.77 0.75 0.73  下载: 导出CSV
下载: 导出CSV
表 2 基于气相色谱-离子迁移鉴定川味发酵香肠挥发性成分
Table 2 Identification fied volatile components of Sichuan fermented sausages based on GC-IMS
类别 化合物 CAS号# 保留指数 保留时间(s) 迁移时间(ms) 醇类 二丙酮醇(2-Hydroxy-2-methyl-4-pentanone) C123422 1358.2 1207.752 1.53264 2-庚醇(2-Heptanol) C543497 1331.2 1127.021 1.37566 异戊醇(1-Butanol, 3-methyl) C123513 1214.3 755.924 1.50193 正丁醇(1-Butanol) C71363 1137.7 559.287 1.38949 2-辛醇(2-Octanol) C123966 1418.8 1389.637 1.86536 异丁醇(1-Propanol, 2-methyl-) C78831 1077 442.018 1.36987 乙醇单体(Ethanol-M) C64175 922.3 300.818 1.04287 乙醇二聚体(Ethanol-D) C64175 921.5 300.394 1.12915 1-戊烯-3-醇(1-Penten-3-ol) C616251 1199.6 708.061 1.34308 2-丁醇(2-Butanol) C78922 1045.5 403.585 1.31695 酯类 辛酸乙酯(Ethyl octanoate) C106321 1419.1 1390.363 1.47109 己酸乙酯二聚体(Ethyl caproate-D) C123660 1243.6 851.268 1.79796 己酸乙酯单体(Ethyl caproate-M) C123660 1242.7 848.086 1.34778 甲酸异戊酯(Isoamyl formate) C110452 1072.3 436.226 1.26483 异戊酸乙酯单体(Butanoicac id 3-methyl ethyl ester-M) C108645 1058 418.865 1.24865 异戊酸乙酯二聚体(Butanoicac id 3-methyl ethyl ester-D) C108645 1057 417.569 1.6541 丁酸乙酯(Butanoic acid ethyl ester) C105544 1047.3 405.765 1.55688 异丁酸乙酯(Ethyl 2-methy lpropionate) C97621 981.5 334.868 1.5676 丙酸乙酯(Ethyl propanoate) C105373 967.6 326.842 1.45361 2-丁烯酸己酯(Hexyl 2-butenoate) C19089920 1248 865.323 1.45154 戊酸乙酯(Ethyl pentanoate) C539822 1148.9 584.817 1.26987 乙酸甲酯(Methyl acetate) C79209 851.1 259.889 1.19629 酮类 仲辛酮(2-Octanone) C111137 1299.1 1030.654 1.32954 α-侧柏酮(α-Thujone) C546805 1419.7 1392.187 1.37168 3-羟基-2-丁酮(2-Butanone, 3-hydroxy) C513860 1266.2 924.656 1.33186 醛类 正丁醛单体(Butanal-M) C123728 853.5 261.293 1.118 苯甲醛(Benzaldehyde) C100527 1518.3 1687.872 1.15767 正丁醛二聚体(Butanal-D) C123728 849.5 258.966 1.28286 丙烯醛(Acrolein) C107028 842.4 254.894 1.06645 丙醛(Propanal) C123386 840 253.524 1.14891 异丁醛(2-Methyl propanal) C78842 821.7 243.035 1.08961 正己醛(1-Hexanal) C66251 1101.3 476.317 1.26798 萜烯类 β-蒎烯单体(β-Pinene-M) C127913 1121.6 522.457 1.22316 β-蒎烯二聚体(β-Pinene-D) C127913 1119.5 517.72 1.6409 (+)-柠檬烯单体((+)-Limonene-M) C138863 1199.6 708.124 1.21957 β-月桂烯单体(β-Myrcene-M) C123353 1162.6 616.176 1.22329 β-月桂烯二聚体(β-Myrcene-D) C123353 1161.9 614.452 1.64633 罗勒烯((Z)-Ocimene) C470826 1215.7 760.281 1.31781 γ-松油烯(γ-Terpinene) C99854 1249.6 870.59 1.21383 (+)-柠檬烯二聚体((+)-Limonene-D) C138863 1199.5 707.736 1.29659 α-松油烯(α-Terpinene) C99865 1180.2 656.311 1.21549 烷烃 1,1-二乙氧基乙烷单体(1,1-Diethoxy ethane-M) C105577 885.1 279.445 1.04309 1,1-二乙氧基乙烷二聚体(1,1-Diethoxy ethane-D) C105577 889.1 281.729 1.12655 胺类 二丁胺(Dibutylamine) C111922 1072 435.916 1.73091 硫化物 烯丙基硫化物(Allyl propyl sulfide) C27817670 1094 462.752 1.37291 二甲基三硫(Dimethyl trisulfide) C3658808 1362.2 1219.901 1.30366 噻唑 噻唑(Thiazole) C288471 1263 914.13 1.25028 2,4,5-三甲基噻唑(2,4,5-Trimethylthiazole) C13623115 1403.6 1344.092 1.56922 吡嗪 2,3-二乙基吡嗪(2,3-Diethylpyrazine) C15707241 1456.8 1503.541 1.71097 2,4,5-三甲基噻唑(2,4,5-Trimethylthiazole) C13623115 1403.6 1344.092 1.56922 芳香烃 丁苯(Benzene, butyl-) C104518 1306.1 1051.558 1.21373 注:“-M”表示单体,“-D”表示二聚体。
下载: 导出CSV
-
[1] 杨瑞征, 李栋钢, 杜前程, 等. 川味香肠调料加工工艺研究[J]. 中国调味品,2015,40(6):72−75. [YANG Ruizheng, LI Donggang, DU Qiancheng, et al. Research on processing technology of Sichuan sausage condiment[J]. China Condiment,2015,40(6):72−75.] YANG Ruizheng, LI Donggang, DU Qiancheng, et al. Research on processing technology of Sichuan sausage condiment[J]. China Condiment, 2015, 40(6): 72−75.
[2] 乔兴, 徐培, 易宇文, 等. 松茸牦牛肉香肠制作工艺[J]. 食品工业,2023,44(10):42−45. [QIAO Xing, XU Pei, YI Yuwen, et al. The processing technology of matsutake yak meat sausages[J]. The Food Industry,2023,44(10):42−45.] QIAO Xing, XU Pei, YI Yuwen, et al. The processing technology of matsutake yak meat sausages[J]. The Food Industry, 2023, 44(10): 42−45.
[3] 董玺梅, 阮志强, 蒋雪薇. 酱香风味传统发酵食品中微生物与风味形成的相关性[J]. 中国食品学报,2022,22(7):397−406. [DONG Ximei, RUAN Zhiqiang, JIANG Xuewei. Correlation between microbes and flavor development in traditional fermented foods with jiang-flavor[J]. Journal of Chinese Institute of Food Science and Technology,2022,22(7):397−406.] DONG Ximei, RUAN Zhiqiang, JIANG Xuewei. Correlation between microbes and flavor development in traditional fermented foods with jiang-flavor[J]. Journal of Chinese Institute of Food Science and Technology, 2022, 22(7): 397−406.
[4] LORENZO J M, FRANCO D. Fat effect on physico-chemical, microbial and textural changes through the manufactured of dry-cured foal sausage Lipolysis, proteolysis and sensory properties[J]. Meat Science,2012,92(4):704−714. doi: 10.1016/j.meatsci.2012.06.026
[5] 杨小莹, 肖珊, 庞煜, 等. 不同发酵剂对发酵香肠品质的影响[J]. 食品与发酵工业,2023,49(18):242−249. [YANG Xiaoying, XIAO Shan, PANG Yu, et al. Different starters on the quality of fermented sausage[J]. Food and Fermentation Industry,2023,49(18):242−249.] YANG Xiaoying, XIAO Shan, PANG Yu, et al. Different starters on the quality of fermented sausage[J]. Food and Fermentation Industry, 2023, 49(18): 242−249.
[6] 黄郑朝, 宋莲军, 黄现青, 等. 基于高通量测序对中国不同区域传统发酵香肠细菌多样性的研究[J]. 食品与发酵工业,2019,45(18):15−21. [HUANG Zhengchao, SONG Lianjun, HUANG Xianqing, et al. Bacterial community in traditional fermented sausages from different regions of China analyzed by high throughput sequencing[J]. Food and Fermentation Industry,2019,45(18):15−21.] HUANG Zhengchao, SONG Lianjun, HUANG Xianqing, et al. Bacterial community in traditional fermented sausages from different regions of China analyzed by high throughput sequencing[J]. Food and Fermentation Industry, 2019, 45(18): 15−21.
[7] 牛茵, 尹礼国, 杨梓垚, 等. 不同加工环境自然发酵羊肉香肠细菌多样性与挥发性风味物质关联分析[J]. 现代食品科技,2023,39(1):270−280. [NIU Yin, YIN Liguo, YANG Ziyao, et al. Correlation analysis of bacterial communities and volatile flavor compounds of naturally fermented mutton saus et al ages from different processing environments[J]. Modern Food Science and Technology,2023,39(1):270−280.] NIU Yin, YIN Liguo, YANG Ziyao, et al. Correlation analysis of bacterial communities and volatile flavor compounds of naturally fermented mutton saus et al ages from different processing environments[J]. Modern Food Science and Technology, 2023, 39(1): 270−280.
[8] 张香美, 叶翠, 卢涵, 等. 发酵香肠制作过程中菌群演替及挥发性风味成分变化规律[J]. 中国食品学报,2022,22(5):282−290. [ZHANG Xiangmei, YE Cui, LU Han, et al. The Succession of bacterial flora and the variation of volatile flavor components during the production of fermented sausage[J]. Journal of Chinese Institute of Food Science and Technology,2022,22(5):282−290.] ZHANG Xiangmei, YE Cui, LU Han, et al. The Succession of bacterial flora and the variation of volatile flavor components during the production of fermented sausage[J]. Journal of Chinese Institute of Food Science and Technology, 2022, 22(5): 282−290.
[9] 李鹤, 陈羽, 李明国, 等. 川味香肠调料配方的优化及风味分析[J]. 食品工业科技,2017,38(20):221−225,249. [LI He, CHEN Yu, LI Mingguo, et al. Optimizing condiment recipes of Sichuan sausage and its flavor analysis[J]. Science and Technology of Food Industry,2017,38(20):221−225,249.] LI He, CHEN Yu, LI Mingguo, et al. Optimizing condiment recipes of Sichuan sausage and its flavor analysis[J]. Science and Technology of Food Industry, 2017, 38(20): 221−225,249.
[10] SUN Q X, CHEN Q, LI F F, et al. Biogenic amine inhibition and quality protection of Harbin dry sausages by inoculation with Staphylococcus xylosus and Lactobacillus plantarum[J]. Food Control, 2016, 68:358-366.
[11] 袁灿, 何莲, 胡金祥, 等. 基于电子舌和电子鼻结合氨基酸分析鱼香肉丝调料风味的差异[J]. 食品工业科技,2022,43(9):48−55. [YUAN Can, HE Lian, HU Jinxiang, et al. The flavour difference of yu-shiang shredded pork seasoning revealed by electronic nose electronic tongue and automatic amino acid analyzer[J]. Science and Technology of Food Industry,2022,43(9):48−55.] YUAN Can, HE Lian, HU Jinxiang, et al. The flavour difference of yu-shiang shredded pork seasoning revealed by electronic nose electronic tongue and automatic amino acid analyzer[J]. Science and Technology of Food Industry, 2022, 43(9): 48−55.
[12] CHEN Q, HU Y Y, WEN R X, et al. Characterisation of the flavour profile of dry fermented sausages with different NaCl substitutes using HS-SPME-GC-MS combined with electronic nose and electronic tongue[J]. Meat Science,2021,172(1):108338.
[13] 曹辰辰, 冯美琴, 孙健, 等. 功能性发酵剂对发酵香肠氧化稳定性及挥发性风味物质的影响[J]. 食品科学,2019,40(20):106−113. [CAO Chenchen, FENG Meiqin, SUN Jian, et al. Effect of functional starter culture on antioxidant and volatile compound in fermented sausages[J]. Food Science,2019,40(20):106−113.] CAO Chenchen, FENG Meiqin, SUN Jian, et al. Effect of functional starter culture on antioxidant and volatile compound in fermented sausages[J]. Food Science, 2019, 40(20): 106−113.
[14] WANG X H, WANG S H, ZHAO H. Unraveling microbial community diversity and succession of Chinese Sichuan sausages during spontaneous fermentation by high-throughput sequencing[J]. Journal of Food Science and Technology,2019,56(7):3254−3263. doi: 10.1007/s13197-019-03781-y
[15] LU X M, LU P Z. Characterization of bacterial communities in sediments receiving various wastewater efiluents with high-throughput sequencing analysis[J]. Microbial Ecology,2014,67(3):612−623. doi: 10.1007/s00248-014-0370-0
[16] 康峻. 浅发酵香肠制作过程中品质特性与微生物多样性的相关性研究[D]. 成都:成都大学, 2021. [KANG Jun. Study on the correlation between quality characteristics and microbial diversity in the production of shallow fermented sausage[D]. Chengdu:Chengdu University, 2021.] KANG Jun. Study on the correlation between quality characteristics and microbial diversity in the production of shallow fermented sausage[D]. Chengdu: Chengdu University, 2021.
[17] 田建军. 传统发酵肉制品中微生物多样性、功能乳酸菌代谢产物及基因序列分析[D]. 呼和浩特:内蒙古农业大学, 2019. [TIAN Jianjun. Analysis of microbial diversity, metabolites and gene sequence of functional lactobacillus in traditional fermented meat products[D]. Hohhot:Inner Mongolia Agricultural University, 2019.] TIAN Jianjun. Analysis of microbial diversity, metabolites and gene sequence of functional lactobacillus in traditional fermented meat products[D]. Hohhot: Inner Mongolia Agricultural University, 2019.
[18] 王新惠, 张雅琳, 孙劲松, 等. 基于高通量测序技术分析广味香肠中细菌群落结构和演替规律[J]. 中国食品学报,2021,21(3):334−340. [WANG Xinhui, ZHANG Yalin, SUN Jinsong, et al. Bacterial community structure and succession law in cantonese sausage based on high-throughput sequencing technology[J]. Journal of Chinese Institute of Food Science and Technology,2021,21(3):334−340.] WANG Xinhui, ZHANG Yalin, SUN Jinsong, et al. Bacterial community structure and succession law in cantonese sausage based on high-throughput sequencing technology[J]. Journal of Chinese Institute of Food Science and Technology, 2021, 21(3): 334−340.
[19] 王玥. 乳酸链球菌素/植物乳杆菌复合物对发酵香肠生物胺及微生物多样性的影响[D]. 雅安:四川农业大学, 2023. [WANG Yue. Effect of nisin/Lactobacillus plantarum complex on biogenic amine and sausage microbial diversity[D]. Yaan:Sichuan Agricultural University, 2023.] WANG Yue. Effect of nisin/Lactobacillus plantarum complex on biogenic amine and sausage microbial diversity[D]. Yaan: Sichuan Agricultural University, 2023.
[20] 赵改名, 李珊珊, 崔文明, 等. 不同来源腊肉中细菌菌群结构与风味相关性分析[J]. 食品与发酵工业,2021,47(13):246−253. [ZHAO Gaiming, LI Shanshan, CUI Wenming, et al. Correlation analysis of bacterial community structure and flavor in different Chinese bacon[J]. Food and Fermentation Industries,2021,47(13):246−253.] ZHAO Gaiming, LI Shanshan, CUI Wenming, et al. Correlation analysis of bacterial community structure and flavor in different Chinese bacon[J]. Food and Fermentation Industries, 2021, 47(13): 246−253.
[21] 张雅琳. 川味香肠发酵过程菌群结构变化与组胺生成相关性的研究[D]. 成都:成都大学, 2020. [ZHANG Yalin. Study on the correlation between microbial community structure changing and histamine production of sichuan-flavor sausage during fermentation[D]. Chengdu:Chengdu University, 2020.] ZHANG Yalin. Study on the correlation between microbial community structure changing and histamine production of sichuan-flavor sausage during fermentation[D]. Chengdu: Chengdu University, 2020.
[22] 牟燕, 赖茂佳, 易宇文, 等. 微生物发酵剂对川味牦牛肉香肠品质的影响[J]. 中国酿造,2024,43(2):188−193. [MOU Yan, LAI Maojia, YI Yuwen, et al. Effect of microbial starter on the quality of Sichuan yak sausage[J]. China Brewing,2024,43(2):188−193.] MOU Yan, LAI Maojia, YI Yuwen, et al. Effect of microbial starter on the quality of Sichuan yak sausage[J]. China Brewing, 2024, 43(2): 188−193.
[23] 范文教, 易宇文, 贾洪锋, 等. 动力学模型预测川味发酵香肠货架期的研究[J]. 食品科学,2013,34(20):334−337. [FAN Wenjiao, YI Yuwen, JIA Hongfeng, et al. Shelf life prediction of sichuan-type sausage through kinetic models[J]. Food Science,2013,34(20):334−337.] FAN Wenjiao, YI Yuwen, JIA Hongfeng, et al. Shelf life prediction of sichuan-type sausage through kinetic models[J]. Food Science, 2013, 34(20): 334−337.
[24] ANGMO K, KUMARIA, MONIKA, et al. Antagonistic activities of lactic acid bacteria from fermented foods and beverage of Ladakh against Yersinia enterocolitica in refrigerated meat[J]. Food Bioscience, 2016, 13: 26-31.
[25] 王宇良, 李志溥, 苏泽佳, 等. 制曲方式对豉香型白酒酒曲理化因子及细菌群落的影响[J]. 食品科学,2023,44(22):211−217. [WANG Yuliang, LI Zhipu, SU Zejia, et al. Effect of preparation methods on the physicochemical properties and bacterial community of chixiangxing baijiu qu[J]. Food Science,2023,44(22):211−217.] WANG Yuliang, LI Zhipu, SU Zejia, et al. Effect of preparation methods on the physicochemical properties and bacterial community of chixiangxing baijiu qu[J]. Food Science, 2023, 44(22): 211−217.
[26] ARO A M J, NYAM-OSOR P, TSUJI K, et al. The effect of starter cultures on proteolytic changes and amino acid content in fermented sausages[J]. Food Chemistry,2010,119(1):279−285. doi: 10.1016/j.foodchem.2009.06.025
[27] CHENG J R, LIU X M, ZHANG Y S. Characterization of Cantonese sausage fermented by a mixed starter culture[J]. Journal of Food Processing and Preservation,2018,42(6):e13623. doi: 10.1111/jfpp.13623
[28] 贾一鸣, 朱丽霞, 张锐利. 无损检测技术在苹果品质检测中的应用[J]. 食品科技,2021,46(4):262−267. [JIA Yiming, ZHU Lixia, ZHANG Ruili. Application of nondestructive testing technology in apple quality testing[J]. Food Science and Technology,2021,46(4):262−267.] JIA Yiming, ZHU Lixia, ZHANG Ruili. Application of nondestructive testing technology in apple quality testing[J]. Food Science and Technology, 2021, 46(4): 262−267.
[29] 李莹, 白凤翎, 励建荣. 发酵海产品中微生物形成挥发性代谢产物研究进展[J]. 食品科学,2015,36(15):255−259. [LI Ying, BAI Fengling, LI Jianrong. Advances in research microbial volatile organic compounds from fermented seafood[J]. Food Science,2015,36(15):255−259.] LI Ying, BAI Fengling, LI Jianrong. Advances in research microbial volatile organic compounds from fermented seafood[J]. Food Science, 2015, 36(15): 255−259.
[30] 李俊霞, 王卫, 王新惠, 等. 发酵香肠风味物质的来源和形成[J]. 食品科技,2015,40(5):160−165. [LI Junxia, WANG Wei, WANG Xinhui, et al. The source and formation of flavor substances in fermented sausage[J]. Food Science and Technology,2015,40(5):160−165.] LI Junxia, WANG Wei, WANG Xinhui, et al. The source and formation of flavor substances in fermented sausage[J]. Food Science and Technology, 2015, 40(5): 160−165.
[31] 谢景丽, 李元景, 陈志强, 等. 离子迁移谱及其联用技术在食品检测中的应用[J]. 现代食品,2018(6):100−106. [XIE Jingli1, LI Yuanjing, CHEN Zhiqiang, et al. Application of ion mobility spectrometry and related hyphenated techniques in food test[J]. Modern Food,2018(6):100−106.] XIE Jingli1, LI Yuanjing, CHEN Zhiqiang, et al. Application of ion mobility spectrometry and related hyphenated techniques in food test[J]. Modern Food, 2018(6): 100−106.
[32] LI H Q, XI B, YANG X L, et al. Evaluation of change in quality indices and volatile flavor components in raw milk during refrigerated storage[J]. LWT, 2022, 165.
[33] GUO L, LUO Y, ZHOU Y, et al. Exploring microbial dynamics associated with flavours prod uction during highland barley wine fermentation[J]. Food Research International,2019,130:108971.
[34] TAN C, FENG B, ZHANG X M, et al. Biopolymer-coated liposomes by electrostatic adsorption of chitosan (chitosomes) as novel delivery systems for carotenoids[J]. Food Hydrocolloids, 2016, 52:774−784.
[35] 张骞, 冯平, 杨海花. 不同加工方式的肉制品中香味物质的形成机制与提取[J]. 食品与发酵工业,2019,45(14):270−276. [ZHANG Qian, FENG Ping, YANG Haihua. Compositions, formation mechanisms and extractions of aroma substances in differently processed meat products[J]. Food and Fermentation Industries,2019,45(14):270−276.] ZHANG Qian, FENG Ping, YANG Haihua. Compositions, formation mechanisms and extractions of aroma substances in differently processed meat products[J]. Food and Fermentation Industries, 2019, 45(14): 270−276.
[36] 张根生, 于敏, 程健博, 等. 香辛料对发酵香肠风味影响的研究现状[J]. 肉类研究,2015,29(5):31−34. [ZHANG Gensheng, YU Min, CHENG Jianbo, et al. Current status of studies on spices effect on the flavor of fermented sausages[J]. Meat Research,2015,29(5):31−34.] ZHANG Gensheng, YU Min, CHENG Jianbo, et al. Current status of studies on spices effect on the flavor of fermented sausages[J]. Meat Research, 2015, 29(5): 31−34.
[37] 姚文生, 蔡莹暄, 刘登勇, 等. 不同材料熏制鸡腿肉挥发性物质GC- IMS指纹图谱分析[J]. 食品科学技术学报,2019,37(6):37−45. [YAO Wensheng, CAI Yingxuan, LIU Dengyong, et al. Volatile compounds analysis in chicken thigh smoked with different materials by gc- ims fingerprint[J]. Journal of Food Science and Technology,2019,37(6):37−45.] YAO Wensheng, CAI Yingxuan, LIU Dengyong, et al. Volatile compounds analysis in chicken thigh smoked with different materials by gc- ims fingerprint[J]. Journal of Food Science and Technology, 2019, 37(6): 37−45.
[38] 尤天棋, 何济坤, 李丹, 等. 川味五香香肠的常规理化和微生物特性及挥发性风味物质分析[J]. 食品工业科技,2024,45(14):224−233. [YOU Tianqi, HE Jikun, LI Dan, et al. Regular physicochemical and microbiological properties and volatile flavors of Sichuan Wu xiang-style sausage[J]. Science and Technology of Food Industry,2024,45(14):224−233.] YOU Tianqi, HE Jikun, LI Dan, et al. Regular physicochemical and microbiological properties and volatile flavors of Sichuan Wu xiang-style sausage[J]. Science and Technology of Food Industry, 2024, 45(14): 224−233.
[39] GIANELLI M P, OLIVARES A, FLORES M. Key aroma components of a dry-cured sausage with high fatcontent (sobrassada)[J]. Food Science & Technology International,2011,17(1):63−71.
[40] 扈莹莹, 王妍, 于晶, 等. 脂肪添加量对发酵香肠脂质和蛋白质氧化及挥发性化合物形成的影响[J]. 食品科学,2019,40(18):8−14. [HU Yingying, WANG Yan, YU Jing, et al. Effect of fat addition on lipid and protein oxidation and volatile compound formation in fermented sausages[J]. Food Science,2019,40(18):8−14.] doi: 10.7506/spkx1002-6630-20181022-233 HU Yingying, WANG Yan, YU Jing, et al. Effect of fat addition on lipid and protein oxidation and volatile compound formation in fermented sausages[J]. Food Science, 2019, 40(18): 8−14. doi: 10.7506/spkx1002-6630-20181022-233
[41] 刘英丽, 万真, 杨梓妍, 等. 乳酸菌对萨拉米香肠风味形成的研究进展[J]. 食品科学,2020,41(23):273−282. [LIU Yingli, WAN Zhen, YANG Ziyan, et al. Recent progress in understanding the contribution of lactic acid bacteria to flavor formation in salami:A review[J]. Food Science,2020,41(23):273−282.] LIU Yingli, WAN Zhen, YANG Ziyan, et al. Recent progress in understanding the contribution of lactic acid bacteria to flavor formation in salami: A review[J]. Food Science, 2020, 41(23): 273−282.
[42] SALVUCCI E, LEBLANC G J, PÉREZ G. Technological properties of Lactic acid bacteria isolated from raw cereal material[J]. LWT-Food Science and Technology,2016,70:185−191. doi: 10.1016/j.lwt.2016.02.043
[43] ZHAO C C, EUN J B. Shotgun metagenomics approach reveals the bacterial community and metabolic pathways in commercial hongeo product, a traditional Korean fermented skate product[J]. [J]. Food Research International,2020,131:109030. doi: 10.1016/j.foodres.2020.109030
[44] 郝倩男, 万嗣宝, 王凤娜, 等. 萨拉米香肠发酵菌株肉糖葡萄球菌冻干保护剂的筛选[J]. 食品科技,2018,43(7):243−249. [HAO Qiannan, WAN Sibao, WANG Fengna, et al. Screening of Staphylococcus carnosus protective agents for salami sausage fermentation[J]. Food Science and Technology,2018,43(7):243−249.] HAO Qiannan, WAN Sibao, WANG Fengna, et al. Screening of Staphylococcus carnosus protective agents for salami sausage fermentation[J]. Food Science and Technology, 2018, 43(7): 243−249.
[45] 赵银峰. 戊糖片球菌和肉葡萄球菌对四川香肠理化、代谢物影响及相关性研究[D]. 成都:成都大学, 2023. [ZHAO Yinfeng. Effects and correlation of Pediococcus pentosaceus and Staphylococcus carnosus on the physicochemical and metabolite profiles of Sichuan sausage[D]. Chengdu:Chengdu University, 2023.] ZHAO Yinfeng. Effects and correlation of Pediococcus pentosaceus and Staphylococcus carnosus on the physicochemical and metabolite profiles of Sichuan sausage[D]. Chengdu: Chengdu University, 2023.
[46] 牛茵, 吴双慧, 何济坤, 等. 基于宏基因组技术分析自然发酵羊肉香肠中微生物多样性及挥发性风味功能基因[J]. 食品科学,2024,45(2):139−148. [NIU Yin, WU Shuanghui, HE Jikun, et al. Metagenomic analysis of microbial diversity and functional genes responsible for volatile flavor of naturally fermented mutton sausage[J]. Food Science,2024,45(2):139−148.] NIU Yin, WU Shuanghui, HE Jikun, et al. Metagenomic analysis of microbial diversity and functional genes responsible for volatile flavor of naturally fermented mutton sausage[J]. Food Science, 2024, 45(2): 139−148.
 下载:
下载:












计量
- 文章访问数: 49
- HTML全文浏览量: 7
- PDF下载量: 7



