


Research Progress on Absorption and Transformation Mechanism and Biological Function of Selenium-enriched Edible Fungi
-
摘要: 富硒食用菌含硒多糖、硒蛋白、硒多肽等多种高生物活性成分,但因为种类、硒生物转化方式和栽培技术等因素的差异,其硒形态组成和有机硒转化率不同,使富硒食用菌表现出不同的生物活性和作用机理。因此,了解硒在食用菌内的各种存在形态,跟踪硒在食用菌内的分布、吸收和转化规律,能更有效地将硒转化为易于人体吸收且无毒的有机硒化合物。本文总结并讨论了代表性富硒食用菌中不同形态硒在总硒中的占比、不同形态硒的吸收和转化过程、总硒的阈值、富硒技术的现状以及基于该吸收转化机制和富硒技术带来的产业问题,并从免疫活性、抗肿瘤活性和抗氧化活性的角度综述了富硒食用菌中硒不同转化形态的生物功能。以期为食用菌的精准富硒调控提供参考依据,为高活性的不同形态硒的开发提供科学支撑,并提出了富硒食用菌发展的方向。Abstract: Selenium-enriched edible fungi are rich in selenium polysaccharides, selenoproteins, selenium polypeptides and other bioactive components. However, the selenium forms and organic selenium transformation rate of selenium-enriched edible fungi are affected by several factors, such as species, selenium biotransformation methods and cultivation techniques. Subsequently, edible fungi show different biological activities and mechanisms of action. Therefore, it is particularly important to study the forms, distribution, absorption and transformation of selenium in edible fungi. The paper reviews the proportion of different forms of selenium in total selenium, the process of absorption and transformation of different selenium forms, the threshold of total selenium, the present selenium-enriched technology and the industrial problems of representative selenium-enriched edible fungi. The immunological activity, antitumor activity and antioxidant activity of different selenium forms in selenium-enriched edible fungi are also discussed in this paper. This review is expected to provide a reference basis for the precise selenium enrichment of edible fungi, to serve as a scientific support for the development of highly active different forms of selenium, and to propose directions for the development of selenium-enriched edible fungi.
-
硒(Selenium,Se)是包括人类在内的生物体发挥重要生物学功能的必需微量元素,也是某些强化食品的重要组成部分[1−3]。人体通过膳食摄入硒后,在机体内以硒代半胱氨酸(Selenocrysteine,SeCys)残基形式参与到相应蛋白质中发挥生物活性作用,具有解除体内重金属毒性、维持心脏健康、保护视器官、抗氧化、抗肿瘤、防癌和提高免疫力的作用[4−5]。
在人类对硒漫长而逐渐深入的认知过程中发现,硒具有敏锐的生物学效应,人体对硒的需要量和中毒剂量阈值较窄,硒缺乏或硒过量都不利于人体健康[6]。缺硒(<40 μg/d)会改变人体甲状腺激素代谢,导致人体免疫力下降,增加患癌风险。严重缺硒(<17 μg/d)将引发克山病(高发于严重缺硒地区的儿童和育龄期妇女的心肌病)、大骨节病(高发于青少年时期的畸形性骨关节疾病)等缺硒相关疾病[7]。硒过量(>400 μg/d)则会产生毒害作用[8],使人体产生皮肤敏感、脱甲脱发、牙齿损害、凝血功能和神经系统功能受损等症状[9]。因此制定适宜不同人群的每日硒摄入量十分重要。根据国家和地区的不同,人体硒日摄入量的范围也不同,具体如表1所示。通过对我国13个省市的营养调查发现,成人硒摄入量仅有26.36 μg/d,远未达到中国营养学会推荐的每日硒摄入量(60 μg/d)[5,10−11]。低水平的硒摄入量对人体健康有潜在危险,然而仅依靠天然食物的摄入难以满足人体对硒摄入的要求,因此科学补硒、从饮食中补充硒,研发安全高效的补硒食品对维持人体健康具有重要意义。
表 1 不同国家/地区人群的硒日摄入量(μg/d)Table 1. Daily intake of selenium by population in different countries/regions (μg/d)国家/地区 适宜人群 平均需要量
(Estimated average
requirement,EAR)推荐摄入量
(Recommend nutrient
intake,RNI)适宜摄入量
(Adequate intake,AI)可耐受最高摄入量
(Tolerable upper intake level,UL)参考文献 中国 婴儿 0~6月 / / 15 55 [10,12] 7~12月 / / 20 80 婴幼儿 1~6周岁 20~25 25~30 / 100~150 儿童 7~13周岁 35~45 40~55 / 200~300 青少年 14~17周岁 50 60 / 350 成人 正常人群 50 60~250 / 400 孕妇 / 50 / 400 乳母 / 65 / 400 苏联 成人 / 50 / 400 [13] 美国 成人 / 60~150 / 400 [14] 加拿大 成人 / 98~224 / 400 [15] 孟加拉 成人 / 63~122 / 400 [16] 注:1岁以下婴儿没有需要量研究资料,只能提出AI值。 当前市场上的补硒剂主要有两类,一类是无机硒,如亚硒酸钠(Na2SeO3)、硒酸钠(Na2SeO4);另一类是有机硒,包括由富硒产地出产的天然产物、有机硒制剂、人工生物转化的各种动植物和微生物产品。相比无机硒,有机硒补剂尤其是富含硒代蛋氨酸的产品在吸收率、安全性和生理活性上更优越。基于天然有机硒资源的限制性和有机硒补剂的优良性,国内外研究者在利用生物转化富集外源硒(主要是无机硒)来生产有机硒补剂方面进行了大量的研究。目前市面在售的有机硒补剂主要有酵母硒、麦芽硒、海藻硒、硒代半胱氨酸和硒代蛋氨酸等几类,其中应用酵母硒和硒代蛋氨酸的人体临床研究案列是相对较多的。美国在酵母的硒转化方面研究的最早[17],在我国徐辉碧是国内率先研制出硒酵母的学者[18]。此后,不断有研究证实食用菌具有一定的耐硒和富硒能力,通过食用菌转化硒,能将环境中的无机硒转化为高效安全的有机硒,并且其中的硒具有很高的生物活性[19],是一种理想的富硒载体。我国食用菌分布全国各地,富含多糖、蛋白、多肽等多种高生物活性成分,同时具有许多人体必需的微量元素。目前已有大量研究以食用菌为富硒载体,培育出富硒酵母、富硒木耳、富硒香菇、富硒灵芝、富硒平菇、富硒金针菇、富硒螺旋藻和富硒猴头菌等[19]富硒食用菌产品。
在“土壤/基料-食用菌-人体”食物链中,硒通过土壤/基料进入食用菌,人体会吸收利用食用菌中70%~90%的硒[20]。然而,我国有72%地域为不同程度的缺硒区,由东北至西南更是形成了一条低硒带,使得硒在“土壤-食用菌”食物链中会存在硒元素流失的现象[21]。因此,充分了解富硒食用菌对硒的吸收、转化机制以及子实体中各部位对硒的吸收特性,选取适宜的生物富硒方式和农学措施提高硒在食用菌中的转化,能够提高其抗病能力、产量、生物功能和产品价值[3],使富硒食用菌中的有机硒更利于人体吸收利用,是实现科学化、规模化、市场化栽培富硒食用菌子实体,提供人类优质食用菌硒源的重要途径。本文对食用菌领域的富硒机制和富硒产品功能的国内外研究进展进行了综述,对今后的研究方向和产业发展方向提出了建议,供食品科技人员参考。
1. 食用菌对硒的吸收转化机制
食用菌中硒的形态组成对其毒性和营养性有着重要影响[22],其化学形态可分为两类,一是无机硒,包括元素硒(Se(0))、亚硒酸盐(SeO32-,Se(IV))和硒酸盐(SeO42−,Se(VI))等形式;二是有机硒,包括低分子量硒化合物如硒代半胱氨酸(Selenocysteine,SeCys)、硒代蛋氨酸(Selenomethionine,SeMet)以及一些挥发性甲基硒化物(二甲基硒、二甲基二硒),和高分子量硒化物主要是含硒酶、含硒蛋白等[23],具体如表2。
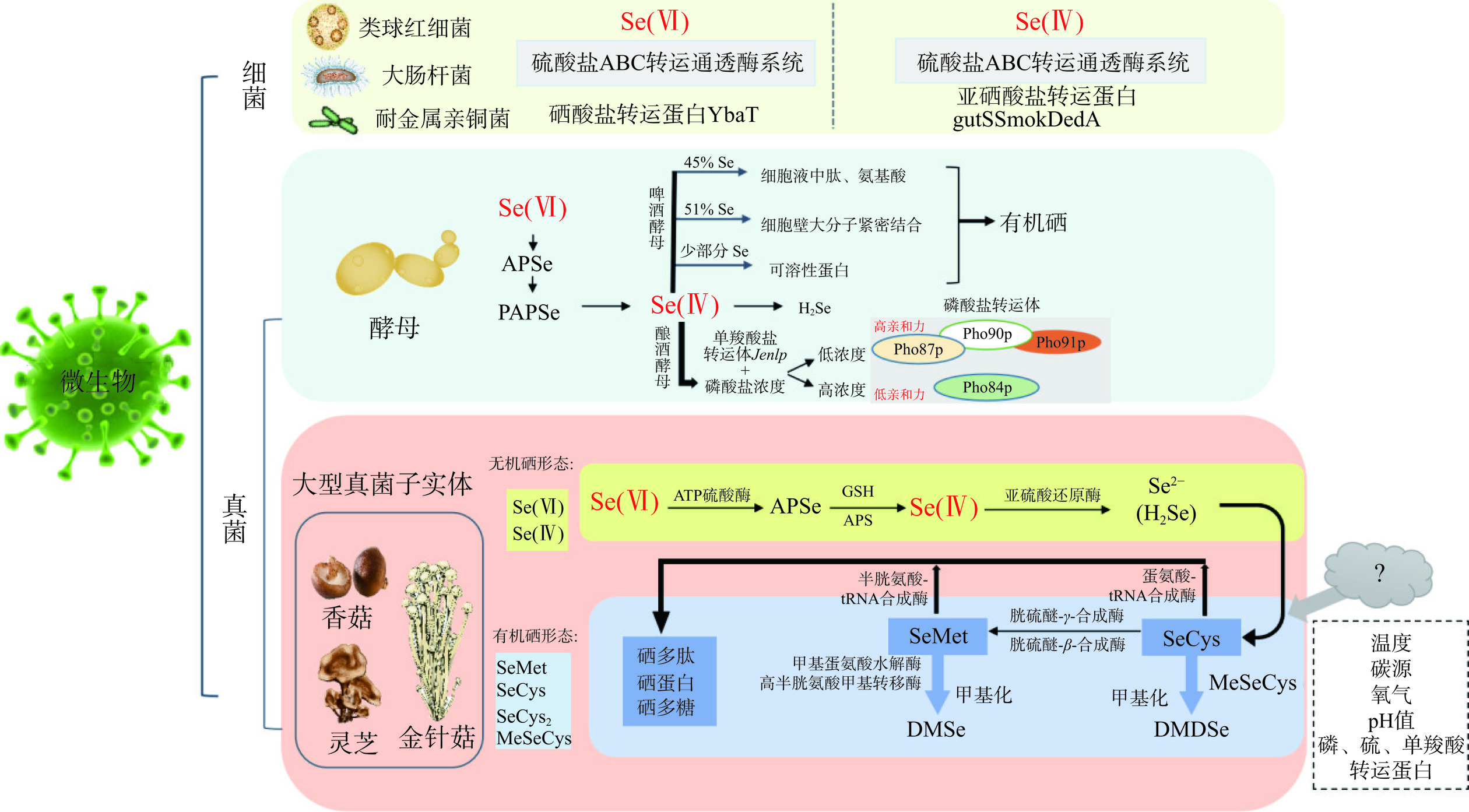
表 2 食用菌中硒的主要形态Table 2. Main forms of selenium in edible fungi类别 名称 分子式 缩写 无机硒 元素硒 Se Se(0) 亚硒酸盐 SeO32− Se(IV) 硒酸盐 SeO42− Se(VI) 硒化物 Se2− Se(-II) 有机硒 低分子量 硒代蛋氨酸 C5H11NO2Se SeMet 硒代胱氨酸 C7H12O4Se2 SeCys2 硒代半胱氨酸 C3H7NO2Se SeCys 甲基硒代半胱氨酸 C4H9NO2Se MeSeCys 二甲基硒 C2H6Se DMSe 二甲基二硒 C2H6Se2 DMDSe 高分子量 含硒酶 / / 含硒蛋白 / / 与植物对硒的吸收转化过程相比,微生物对硒的吸收转化并不需要叶绿体的参与。对于硒在微生物中的吸收转运机制,有关酵母的研究最早,细菌次之[24]。硒在酵母细胞内的代谢涉及还原、氧化、同化、转运和甲基化等过程[25],而在大型真菌中的转化过程目前还尚不清楚[26]。如图1所示,在酵母细胞中,Se(VI)在ATP-硫酸化酶催化下转化为APSe,接着APSe在相应的激酶作用下转化为PAPSe,PAPSe再经过对应还原酶作用被还原为Se(IV),最后在硫酸盐还原酶作用下转化为终产物硒化氢(H2Se)[27]。其中,啤酒酵母对于Se(IV)的吸收中有45%的硒与细胞液中的肽、氨基酸结合,51%的硒与酵母细胞壁的大分子组分紧密结合,剩余硒则与可溶性蛋白质结合,以此将无机态硒被转变为有机态[28]。在酿酒酵母中单羧酸转运体Jen1p参与Se(IV)的转运,此外还依赖于生长介质中的磷酸盐浓度,低浓度的磷酸盐通过高亲和力的磷酸盐转运体Pho84p;高浓度磷酸盐通过低亲和力的磷酸盐转运体Pho87p、Pho90p和Pho91p[29]。在细菌中Se(IV)和Se(VI)均可通过硫酸盐ABC转运通透酶系统进入细胞。Se(IV)还可通过相关的转运蛋白gutS、SmoK、DedA进入细胞,Se(VI)可通过转运蛋白YbaT进入细胞[30−31]。
![]() 图 1 硒在微生物中的吸收转化过程(食用菌为大型真菌)注:APSe:5’-磷酸硒腺苷;GSH:谷胱甘肽;APS:5’-磷硫酸腺苷还原酶;H2Se:硒化氢。Figure 1. Absorption and transformation process of selenium in microorganism (edible fungi are macrofungi)
图 1 硒在微生物中的吸收转化过程(食用菌为大型真菌)注:APSe:5’-磷酸硒腺苷;GSH:谷胱甘肽;APS:5’-磷硫酸腺苷还原酶;H2Se:硒化氢。Figure 1. Absorption and transformation process of selenium in microorganism (edible fungi are macrofungi)在真菌的研究中发现,硒主要以胞外结合和离子转运的方式在细胞内累积与代谢[25,32]。食用菌细胞壁上的大量硫基、氨基等离子团具有生物吸附作用,在菌丝体中细胞壁的几丁质和其他成分是吸收硒的主要部位,硒在栽培过程中通过生物转化到子实体中使其高效富硒。例如在富硒糙皮侧耳的细胞壁中分布56%的硒[33]。真菌对不同硒形态的吸收受到温度、氧气、pH、碳源和转运蛋白(主要是磷酸盐、硫酸盐和单羧酸盐转运蛋白)等因素的影响[34]。当前很少有关于食用菌对硒的吸收通道方面的研究,仅有研究发现,添加呼吸抑制剂能减少金针菇菌丝中亚硒酸盐的吸收量(11%),并推测金针菇吸收亚硒酸盐的主要方式是被动运输[34]。
根据硒在酵母的吸收转化以及耐硒真菌对植物生长和硒积累的作用相关研究[25,35]推测,食用菌对硒的吸收转化机制可能包括硒的吸收、转化、转运和积累过程,但目前关于食用菌对不同形态硒的吸收通道和转化过程尚不明确。如图1所示,Se(VI)被食用菌吸收后,由ATP硫酸酶激活生成APSe,在谷胱甘肽(GSH)和5’-磷硫酸腺苷还原酶(APS)的共同作用下被还原成Se(IV),之后由亚硫酸还原酶还原成硒化物(Se2−),Se2−在硒代半胱氨酸合成酶的作用下合成SeCys,再经过胱硫醚-β-裂解酶和胱硫醚-γ-合成酶的作用合成SeMet,而SeCys和SeMet分别由半胱氨酸t-RNA合成酶和蛋氨酸t-RNA合成酶合成硒蛋白。SeCys和SeMet也可通过甲基化形成二甲基二硒化物。
食用菌将无机硒转化为对人体有益的高活性有机硒,这一转化过程涉及多种酶类和代谢途径的参与,如巴西伞菌(Agaricus blazei)、白灵菇(Lentinula edodes)、牛肝菌属(Boletus)、蘑菇属(Agaricus)等食用菌通过硒还原酶代谢途径将硒酸盐还原为硒化物形式;香菇属(Agaricus)、松茸菌(Matsutake)等通过硒甲基化代谢途径(硒甲基转移酶)将硒酸盐甲基化为硒化物[36−37]。这些代谢途径的发挥通常与食用菌的菌株、生长环境、培养条件等因素有关[38]。研究人员试图通过组学技术来探究有机硒在农作物中的分布和功能,如Zhao等[39]通过对富硒金顶侧耳进行转录组测序并分析相关的差异表达基因(共1036个差异表达基因,987个上调基因,49个下调基因)和涉及的20条代谢途径发现,与硒代谢密切相关的硫代谢和ABS转运蛋白均十分富集。Xu等[40]对灵芝进行了基因组测序,对可能的硒代谢途径进行了全基因组的功能注释,发现硒主要参与硒代蛋氨酸(SeMet)、硒代胱氨酸(SeCys)、甲基硒代半胱氨酸(MeSeCys)等含硒氨基酸(有机硒形态)的合成,但关于硒多糖以及其它含硒化合物的合成途径尚未作解释。
通过对富硒酵母中硒的分布与吸收转运的大量研究[36,41−42],推测转化后的有机硒可能通过转运蛋白、离子通道、硫氢化物反应、细胞器定位、蛋白质结合等分子机制运输到富硒食用菌的细胞内,并在细胞内积累。这些机制将有助于富硒食用菌稳定储存和有效利用硒。具体的硒转运和积累机制可能会因不同的富硒食用菌种类而有所差异[43]。此外,在硒的转运和积累过程中可能还会受到细胞内信号传导、基因表达调控、环境等多种因素的调控[44]。
多数食用菌对有机硒的吸收显著强于无机硒(硒酸盐/亚硒酸盐)[45],但其具体机制尚不清晰,有研究推测食用菌通过水通道和钾离子通道共同参与主动吸收硒代蛋氨酸,并通过根系的蛋氨酸转运蛋白进行转运,但其深入的机制仍需要进一步的研究。
2. 食用菌富硒技术的现状
基于硒的吸收转化机制的富硒技术是富硒食用菌产业的核心,对富硒食用菌产业的发展起着举足轻重的作用。目前国际上主要通过向栽培基料中直接添加不同浓度的无机硒(Na2SeO3和Na2SeO4)、有机硒(SeMet)、硒制品(纳米硒)或富硒农作物的废弃物(富硒小麦和水稻秸秆),再结合人工培养的技术实现对硒的生物强化[46]。这些富硒技术的富硒效果受到土壤/基料的酸碱性、食用菌种类、硒源种类、施硒方式、施硒浓度、施硒时期等多种因素影响,硒的吸收转化率具有较大差异。
硒的吸收转化率似乎与食用菌的种类更相关。例如将亚硒酸钠(12 mg/L)施于猴头菌的培养基中,猴头菌含硒量达到139.1 mg/kg,且有机硒转化率达到98.37%[45],而将相同硒源(55 mg/kg)加入秀珍菇的栽培原料中,其子实体含硒量仅到达2.812 mg/kg[39]。此外,以相同浓度的亚硒酸钠(0~1.5 mmol/L)分别对灵芝和茶树菇进行富硒,发现灵芝子实体的硒含量大约是茶树菇的2倍,其中灵芝的有机硒转化率可以达到100%[47]。
同一类食用菌,硒含量与硒源似乎更相关。例如在黑木耳基料中添加0~250 mg/kg的生物纳米硒,其子实体含硒量为0.03~85.7 mg/kg,而以亚硒酸钠为硒源,其子实体中硒含量能大于100 mg/kg[48]。再如使用同等浓度的硒脲和亚硒酸钠处理云芝蘑菇,经亚硒酸钠处理后的硒含量显著更高,但经硒脲处理的含SeMet量更高[49]。还发现,不同的硒源经食用菌吸收转化后其主要的硒形态组成也不一样。例如分别以富硒酵母和亚硒酸钠对双孢菇进行富硒,通过富硒酵母培育的双孢菇中主要硒形态是SeMet、Se(IV)、SeCys2、Se(IV),比使用亚硒酸钠培育的子实体中的硒形态组成更丰富[50−51]。
施硒方式也是影响食用菌中主要硒形态的重要因素。例如分别以亚硒酸钠浸泡和灌溉香菇,经浸泡的香菇中主要硒形态是Se(IV)、SeMet和SeCys2,而通过灌溉后其硒形态还增加了MeSeCys[52]。然而,同样以灌溉亚硒酸钠的方式分别处理香菇和双孢菇后[53−55],两者子实体中的主要硒形态组成相同。
在碱性土壤/基料中,硒酸盐的吸收转化率高。采用在土壤中施加硒肥的方式,部分农作物的硒吸收率只有12%[56]。在不同生长阶段施硒,食用菌对硒的吸收转化率也不相同[57−59]。此外,依据硒元素与不同离子间的拮抗或协同作用,可以添加调控剂来影响富硒效果[60]。表3总结了代表性食用菌种类、硒源种类、硒源浓度和施硒方式对硒吸收转化的影响。
表 3 代表性食用菌对硒的吸收和转化Table 3. Absorption and transformation of selenium by representative edible fungi食用菌种类 施硒方式 硒源 培养基中硒浓度 子实体硒含量(mg/kg) 主要硒形态组成/有机硒含量(mg/kg)/
有机硒转化率(%)参考文献 猴头菌 / 亚硒酸钠 12 mg/L 菌丝139.1 98.37% [61] 秀珍菇 / 亚硒酸钠 55 mg/kg 2.812 / [62] 大球盖菇 / 亚硒酸钠 150 mg/L 4727. 68 96. 27% [63] 黑木耳 / 生物纳米硒 0-250 mg/kg 0.03-85.70 / [64] / 亚硒酸钠 250 mg/kg >100 / 灵芝 / 亚硒酸钠 0~1.5 mmol/L 28~73 100% [47] 茶树菇 / 亚硒酸钠 0~1.5 mmol/L 12~34 84% [47] 巴西菇 / / / 9.15 SeCys2 5.734 mg/kg;
MeSeCys 1.84 mg/kg;SeMet 0.22 mg/kg[65] 平菇 / 富硒酵母 / 3 / [66] 拌料 亚硒酸钠 / 267.6 SeMet,SeCys2,Se(IV),Se(VI) [67] 香菇 浸泡 亚硒酸钠 / / Se(IV),SeMet,SeCys2 [52] 灌溉 亚硒酸钠 / / 无机硒,MeSeCys,SeCys,SeMet [53] 浸泡 硒酸钠 / 174.5 Se(VI),SeMet [67] 金针菇 拌料 亚硒酸钠 5 mg/kg 菌盖86.7
菌柄45.3/ [68] 双孢菇 拌料 富硒酵母 / 160 SeMet,Se(IV),SeMet,SeCys2, [50] 拌料 亚硒酸钠 / / Se(IV),SeCys2 [51] 喷施 亚硒酸钠 10~40 mg/kg 12.2~415 MeSeCys,SeCys2,SeMet [54] 灌溉 亚硒酸钠 10~40 mg/L 菌盖347
菌柄415无机硒,SeCys,MeSeCys,SeMet [54−55] 3. 基于硒的吸收转化机制和富硒技术带来的产业问题
基于硒的吸收转化机制,目前的富硒技术可以将食用菌硒含量富集到一个很高的水平,但是我国的富硒技术主要以亚硒酸钠为硒源,硒元素容易流失,栽培基料利用率不高,而且有时某些有毒的硒化合物(如甲基硒酸、乙基硒酸)会残留在食用菌中造成硒中毒[46]。此外,大量研究报道表明亚硒酸钠(无机硒)在食用菌的富集过程中会带入铬等重金属,使得产品重金属含量超标风险增加[69−71]。因此需要通过改良栽培基料配方、精细调控过程参数等开发新型富硒调控技术,解决现有技术导致的高活性有机硒占比低(硒流失)、伴随危害重金属超标、栽培基料成本增加、培育过程繁琐等问题,提高富硒食用菌栽培的经济性、安全性和品质。
4. 富硒食用菌中主要硒转化形态的生理活性
硒多糖、硒蛋白和硒多肽作为有机硒的主要转化形态,兼具有硒和多糖、蛋白、多肽的生理活性,相比于无机硒的高毒性和不宜吸收,有机硒毒副作用低,更易被人体吸收,且具有增强免疫、抗癌、抗氧化等生理活性,因此被越来越广泛地应用于医学及生命科学研究中[72]。
4.1 免疫活性
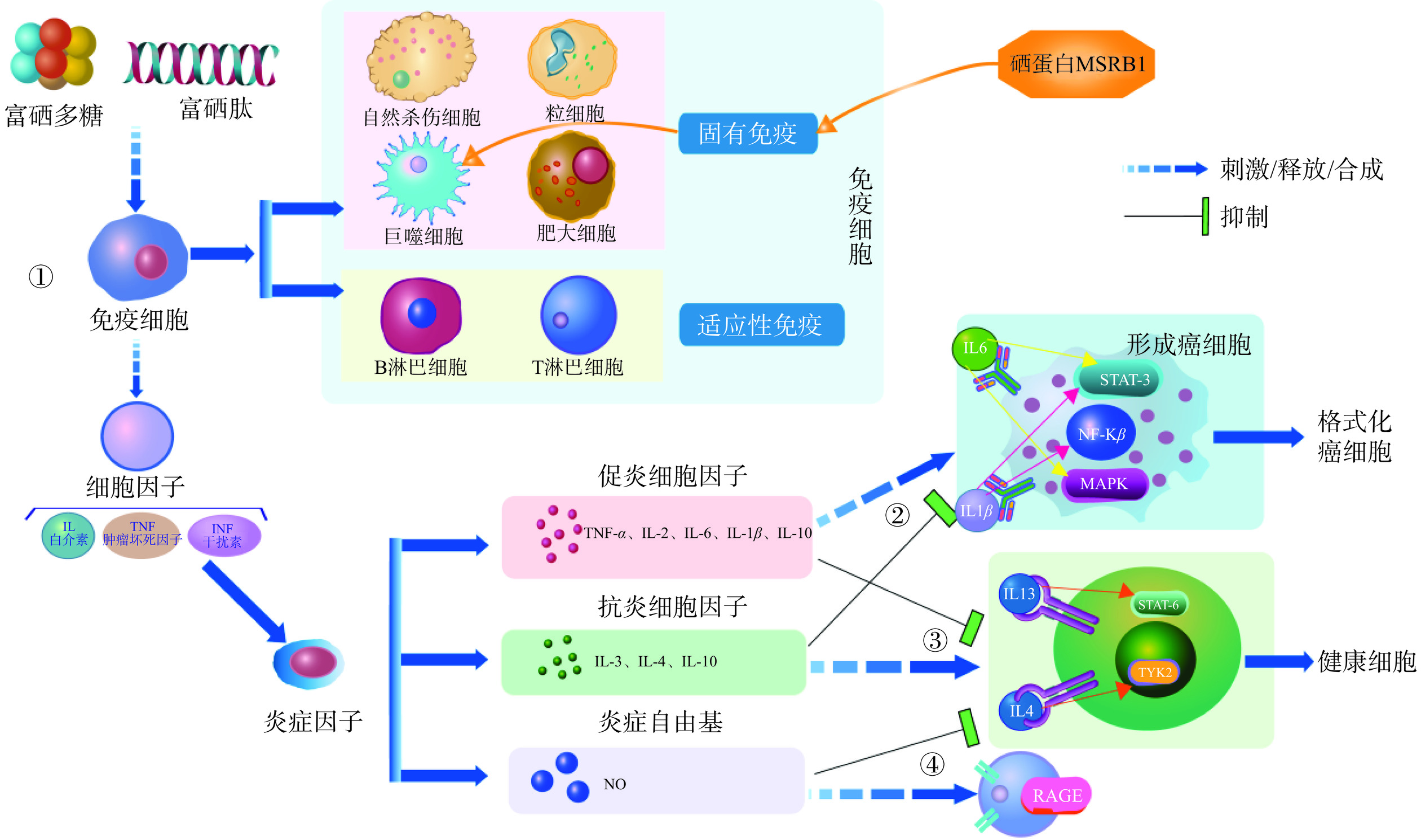
免疫活性是多糖最显著的生物学活性之一[73],而硒也显著影响机体的免疫系统。越来越多的药理及临床研究表明食用菌硒多糖可以通过促进细胞因子产生、影响细胞信号转导等途径调节和增强细胞的免疫功能。本文通过图2总结了细胞的免疫基本模型[74]。
![]() 图 2 硒转化形态(功能组分)的免疫活性注:富硒多糖/富硒肽可通过刺激免疫细胞(分为发挥固有免疫和适应性免疫的细胞)释放细胞因子。其中,硒蛋白MSRB1是通过促进巨噬细胞中抗炎细胞因子的表达来发挥机体固有免疫功能;促炎细胞因子的大量合成(如IL-6、IL-1β)或抑制抗炎细胞因子释放(如IL-13、IL-4),会激活细胞内MAPK/NF-κB/STAT-3信号通路上相关蛋白的表达,使细胞产生病变,形成癌细胞,最终正常细胞被癌细胞格式化;释放抗炎细胞因子(如IL-13、IL-4)或抑制促炎细胞因子(如IL-6、IL-1β)的合成能表现出强大的抗炎活性,从而避免免疫反应发生以及癌细胞的形成,维持正常细胞的健康;抑制炎症自由基NO的合成能表现出强大的抗炎活性,而炎症自由基NO的大量释放会激活体内RAGE信号通路上相关蛋白的表达,使机体产生炎症反应(但当NO作为一种免疫信号时其效果反之)。Figure 2. Immunoactivity of the transformed form of selenium
图 2 硒转化形态(功能组分)的免疫活性注:富硒多糖/富硒肽可通过刺激免疫细胞(分为发挥固有免疫和适应性免疫的细胞)释放细胞因子。其中,硒蛋白MSRB1是通过促进巨噬细胞中抗炎细胞因子的表达来发挥机体固有免疫功能;促炎细胞因子的大量合成(如IL-6、IL-1β)或抑制抗炎细胞因子释放(如IL-13、IL-4),会激活细胞内MAPK/NF-κB/STAT-3信号通路上相关蛋白的表达,使细胞产生病变,形成癌细胞,最终正常细胞被癌细胞格式化;释放抗炎细胞因子(如IL-13、IL-4)或抑制促炎细胞因子(如IL-6、IL-1β)的合成能表现出强大的抗炎活性,从而避免免疫反应发生以及癌细胞的形成,维持正常细胞的健康;抑制炎症自由基NO的合成能表现出强大的抗炎活性,而炎症自由基NO的大量释放会激活体内RAGE信号通路上相关蛋白的表达,使机体产生炎症反应(但当NO作为一种免疫信号时其效果反之)。Figure 2. Immunoactivity of the transformed form of selenium大量研究表明硒多糖通过刺激免疫细胞释放细胞因子白介素(IL)、干扰素(INF)、肿瘤坏死因子(TNF)等发挥功能(图2)。其中由免疫细胞释放促炎细胞因子和抗炎细胞因子是免疫系统发挥功能的关键[75]。例如富硒姬松茸多糖能显著提高荷瘤小鼠的免疫细胞(自然杀伤NK细胞)的杀伤活性,促进淋巴细胞转化进而提高机体免疫[73]。此外,还发现富硒灰树花多糖不仅能够调节免疫细胞的活性促进细胞免疫反应,还能够调节肠道微生物群落改善肠道健康,并通过肠道免疫系统增强机体免疫功能[76]。
硒多糖能通过影响免疫细胞信号NO、环磷酸腺苷和环磷鸟嘌呤核苷的转导与活化等实现增强免疫力的作用。Wang等[66]通过体外细胞实验发现,富硒平菇多糖可通过上调共刺激分子CD80/CD86的表达,激活NF-κB蛋白,促进小鼠单核巨噬细胞(RAW264.7细胞)分泌细胞因子(如TNF-α、IL-1β和IL-6)和NO,从而显著增强RAW264.7细胞的增殖和免疫能力。
硒多糖还能进一步通过影响细胞因子和免疫细胞信号在免疫和炎症相关信号通路中的产生,应对各种炎症的发生。研究表明富硒黑木耳多糖通过减少小鼠海马体中促炎细胞因子(TNF-α、IL-6和IL-1β)的释放,下调了蛋白如RAGE(晚期糖基化终产物的受体)、细胞外调节的蛋白激酶(ERK)、c-Jun N-末端激酶(JNK)和核酸核酸酶p蛋白亚基(p38)在RAGE/MAPK/NF-κB通路的表达,使其能持续抑制由D-半乳糖诱导的神经炎症[77]。而富硒灰树花多糖可以强烈刺激免疫信号NO的产生,同时通过上调MAPKs信号通路上JNK、ERK、p38等转录因子来保护经环磷酰胺诱导的小鼠免于免疫抑制[76]。这些结果表明硒多糖可通过抑制促炎细胞因子的合成表现出强大的抗炎活性,且在未来可作为一类潜在的缓解脑组织炎症的新型治疗物质,也具备减轻由化疗引起的免疫抑制能力。
目前对于硒蛋白和硒多肽的免疫活性研究尚不全面。迄今为止已经纯化了15种硒蛋白,并进一步表征了它们的生物学功能,其中4种是谷胱甘肽过氧化物酶(经典的GPx1,胃肠道GPx2,血浆GPx3,磷脂氢过氧化物GPx4)。除了这一类抗氧化的硒蛋白,还有硒蛋白-P、硫氧还蛋白(Rdx)、碘甲腺原氨酸脱碘酶(DIO)等在食用菌和哺乳动物中均起到特别重要的作用[78]。硒蛋白主要是通过参与炎症反应相关基因mRNA的表达和翻译水平影响机体免疫系统。例如有学者试图通过转基因和敲除小鼠模型阐明单一硒蛋白在调节免疫细胞功能中的作用,发现硒蛋白MSRB1(亚砜还原酶B1)对机体产生先天免疫反应十分重要,该硒蛋白可通过促进巨噬细胞中抗炎细胞因子的表达来控制免疫反应[79](图2);而硒蛋白SELENOK(硒蛋白K)在小鼠免疫组织中的水平相对较高,该硒蛋白作为参与蛋白质翻译后修饰酶的关键辅助因子,在调节免疫中发挥重要作用[40]。此外,有研究发现在使用富硒蛋白饲喂小鼠时,由于谷氧还蛋白(Grx)和硫氧化还原蛋白(Trx)的应激反应,延迟了细胞的衰老,使得整个机体的免疫功能加强[80]。还有研究评估了富硒灰树花糖蛋白对As3+(砷被认为具有高度细胞毒性)诱导的免疫毒性的改善作用,结果显示该物质可通过上调由NF-κB和MAPK信号通路介导的细胞因子(IL-2和INF-γ)的分泌来减轻As3+的免疫毒性[81]。但关于硒蛋白如何影响不同类型的免疫反应以及更深入的机制还有待进一步研究。
硒多肽也能通过抑制NF-κB的活化来降低促炎细胞因子的水平(图2)。例如在富硒冬虫夏草对巨噬细胞的免疫调节效应的研究中发现,硒多肽能够促进巨噬细胞的活性和细胞因子的产生,并通过NF-κB信号通路介导免疫应答[82]。硒多肽还可以影响细胞因子和免疫细胞信号的产生来应对机体炎症,增强机体的免疫功能。例如Zhang等[83]使用富硒大豆肽降低了小鼠中促炎细胞因子(IL-1β、IL-6和TNF-α)水平,并通过抑制肺中TLR4、NF-κB、IκBα、NLRP3、ASC和caspase-1蛋白的表达,保护小鼠肺部免受由PM2.5引起的炎症。此外,富硒大豆肽还可以通过上调小鼠脾脏中细胞因子(IL-2、IFN-γ)和免疫细胞信号iNOS的mRNA表达,增强宿主的特异性和非特异性免疫,有效地抑制由环磷酰胺诱导的免疫抑制,且较富硒大豆蛋白表现出更强的免疫调节活性[84]。同时,硒多肽能通过增加机体内的含硒酶来增强机体免疫器官的免疫值。例如胡居吾[85]通过对小鼠腋下注射S180瘤株,发现天然富硒大豆肽还可以通过增加机体内的含硒酶,使得机体免疫器官加强,尤其会促进胸腺的发育。而从富硒大米中提取的含硒肽也具有很高的免疫调节活性[86]。这些结果表明富硒肽具有成为调节和增强免疫的膳食补充剂的巨大潜力。
4.2 抗肿瘤活性
食用菌硒多糖具有抗肿瘤活性。有动物试验结果表明,姬松茸硒多糖在抗肿瘤过程中能对肿瘤细胞的凋亡起到重要作用[87]。Ma等[88]通过细胞实验发现,富硒平菇多糖对正常细胞无明显影响,但能通过降低A549(人肺癌细胞)、SKOV3(人卵巢癌细胞)、HepG2(人肝癌细胞)和MCF-7(人乳腺癌细胞)的活力诱导A549细胞凋亡。富硒平菇多糖抑制了癌细胞从上皮细胞到间质细胞的转化,具有成为天然抗肿瘤药物的巨大潜力。陈彻[89]首次在体内和体外研究了富硒茶多糖对肉瘤180(源于叶间组织的恶性肿瘤)的抗肿瘤活性,发现在硒和茶多糖这两种物质的协同作用下的抗肿瘤效果优于常规的富硒酵母,这说明硒和多糖协同作用能表现出更强大的抗肿瘤活性。再如,使用经姬松茸硒多糖处理过的血清可以上调细胞凋亡蛋白Caspase-3的活性,诱导人慢性髓原白血病K562细胞(在自然杀伤分析中广泛作为高敏感的体外受体)的凋亡[89]。从富硒梭柄松苞菇(Catathelasma ventricosum)中提取的硒多糖能够显著抑制肿瘤细胞的生长,并且能够增强小鼠的免疫功能[90]。此外,通过体外细胞培养并给予硒化的白沙蒿多糖(硒含量1703 μg/g)处理,发现该物质能提高人肝癌HepG-2细胞、人肺腺癌A549细胞和人宫颈上皮Hela癌细胞的抗增殖活性[91]。
硒蛋白可降低患肿瘤概率。例如,在松茸硒硒蛋白(SPP)的体内体外抗肿瘤活性实验研究中发现,SPP在体内可抑制移植肉瘤180的生长,促进荷瘤小鼠淋巴细胞转化和提高自然杀伤细胞(NK)活性。在体外实验中,经SPP处理的血清还能抑制K562细胞的增殖,引起细胞凋亡形态学改变和核间体DNA断裂[65]。此外,从苦荞中提取的硒蛋白能显著抑制人乳腺癌细胞Bcap37的增值作用,其抗肿瘤效果与时间和剂量相关[79]。
关于硒多肽的抗肿瘤活性,有研究发现天然富硒大豆肽的浓度与肿瘤细胞的大小呈现负相关性,当其浓度越高,肿瘤细胞逐渐缩小,细胞质中的颗粒增加致使大量细胞破碎,引发肿瘤细胞的凋亡[92]。此外,富硒香菇中硒多肽能够抑制血管生成并诱导肿瘤细胞凋亡,从而抑制肿瘤的生长和扩散[87]。目前有研究开发了一种自组装的硒肽纳米颗粒[92],该纳米颗粒通过硒肽的氧化代谢物激活NK细胞从而增强肿瘤化学免疫疗法的效果。预计在未来,硒肽或可作为抗肿瘤剂应用于多种生物医学中。
4.3 抗氧化活性
人体在外界因素影响下会不断产生自由基,自由基过量会加速机体衰老,甚至存在患癌风险。抗氧化则是具有抗氧化性的物质与自由基发生反应,能阻止自由基的进一步生成。硒多糖相较普通多糖具有更强的抗氧化能力。例如Gu等[93]对富硒茶叶粗多糖进行分级纯化并得到了SeTPS-1和SeTPS-2,发现这两种富硒茶多糖组分具有比茶多糖更强的羟基自由基清除能力、ABTS+自由基清除能力、还原力及DNA损伤保护活性。从富硒鸡腿菇菌丝体中分离出的硒多糖能够对四氧嘧啶诱导的小鼠氧化应激起到保护作用[94]。另外,通过在D-半乳糖诱导下建立亚急性衰老动物模型,发现富硒蛹虫草多糖也具有极显著的抗氧化活性[82]。目前在食用菌硒多糖中发挥抗氧化活性成分的构效关系尚不明确,但发现食用菌硒多糖能通过降低炎症因子mRNA表达,调节相关凋亡蛋白的表达等分子机制来减缓炎症的发生并增强机体的抗氧功能。例如Gu等[95]通过体外试验证明使用复合酶法提取的富硒香菇相对于普通香菇具有更强的抗氧化和抗炎症作用,能显著降低RAW264.7细胞系中由脂多糖(LPS)引起的mRNA表达和促炎因子(NO、IL-6/1β和TNF-α)水平,表明富硒香菇可作为一种良好的膳食硒补充剂。此外,Ma等[88]发现经过纯化的富硒平菇多糖可通过上调抗凋亡蛋白Bcl-2,下调促凋亡蛋白Bax、cleaved caspase 3和细胞色素c,进而对过氧化氢(H2O2)诱导的PC12细胞氧化损伤和凋亡起到明显的缓解作用。
硒蛋白相较于硒多糖更具抗氧化能力,其抗氧化机理主要包括增强抗氧化相关酶活性、清除自由基、抑制丙二醛(malondialdehyde,MDA)和脂质过氧化物的产生、螯合过渡金属离子进行解毒等。赵镭等[96]发现,在富硒灵芝子实体中硒与蛋白具有抗氧化协同作用,而硒与灵芝多糖、灵芝蛋白与灵芝多糖之间则无该协同作用但能彼此增强抗氧化活性。在双孢蘑菇的研究中发现,硒蛋白可以显著增强抗氧化酶活性,对细胞氧化损伤具有保护效应[39]。在牛肝菌中发现硒蛋白具有显著的自由基清除活性和抗氧化能力,能够抑制炎症反应[97]。此外,有研究表明纯化的恩施碎米荠硒蛋白对小鼠线粒体中MDA的生成有明显抑制作用,当其硒浓度大于0.95 μg/mL时,对小鼠体外Fe2+诱导的脂质过氧化有明显抑制作用;而在硒浓度处于0.15~5.96 μg/mL时,则能有效缓解由H2O2引起的氧化过程[98]。有研究表明硒蛋白-P能抵抗氧化损伤并将硒运输到大脑。此外,多数硒蛋白中硒代胱氨酸和硒代组氨酸含量的多少与金属的解毒功能密切有关[99]。
当前对于硒多肽抗氧化能力的研究较少,发现硒多肽可以通过促进内源性抗氧化酶的产生,调节在炎症相关通路上的蛋白表达,发挥机体抗氧化作用。Guo等[100]通过体外自由基清除和脂质过氧化抑制试验发现,富硒肽(SSeP)可以通过增加皮肤或血清中的GPx和过氧化氢酶活性以及谷胱甘肽含量来有效缓解UVB辐射引起的皮肤损伤和氧化应激。同时,富硒肽在培养的人表皮角质形成细胞(HaCaT)中表现出对H2O2诱导的细胞毒性的保护作用,这可能是由于SSeP通过增加水通道蛋白-3的表达和减弱p38 MAPK的磷酸化的作用。该研究表明富硒肽是一种很有前景的抗氧化营养素,可作为食品添加剂增强人体抗氧化能力,也可作为药妆产品减少皮肤氧化损伤。在D-半乳糖诱导的衰老小鼠模型中显示,含硒大豆肽能显著降低NF-κB水平,同时肝超氧化物酶歧化酶(SOD)、谷胱甘肽过氧化物酶-1(GPx-1)、天冬氨酸氨基转移酶(AST)及丙氨酸氨基转移酶(ALT)均发生了显著性下降。在该模型中含硒大豆肽还可通过调节MAPK/NF-κB通路抑制脑氧化应激。此外,与亚硒酸钠、硒代蛋氨酸及无硒肽相比,含硒大豆肽可以达到最高的抗氧化活性,且在抗氧化活性方面表现出硒和肽的协同作用[101]。
综上,食用菌的硒多糖、硒蛋白、硒多肽均具有强大的免疫活性、抗肿瘤活性和抗氧化活性。相对而言,硒多糖具有更强的免疫活性和抗肿瘤活性,硒蛋白具有更强的抗氧化能力,未来或可将硒多糖、硒蛋白和硒多肽作为高活性、高价值的膳食补充剂来开发。另外,肠道含有丰富的肠道菌群和免疫炎症蛋白,随着越来越多关于肠道菌群对人体健康全面影响的研究,以及免疫炎症蛋白对免疫状态的作用被大量报道,进一步探究硒多糖和硒蛋白(硒多肽)对肠道菌群多样性的调节和对免疫炎症蛋白的干预,必能使功能因子硒多糖和硒蛋白(硒多肽)的优势功效得到进一步的挖掘。
5. 富硒食用菌可能的发展方向
以香菇、木耳为主的食用菌产业是我国具有优势的战略资源之一,但目前产品过于单一、多元化加工不足,没有形成优势深加工产品,产品附加值低,市场竞争力严重不足,很难抵御国际市场激烈竞争所带来的风险。近年来,《“健康中国2030”规划纲要》《国民营养计划(2017~2030)》《健康中国行动(2019~2030)》等国家相关规划纲要的相继出台,以及精准营养科学研究的进步和人们生活水平的提高,为营养健康产业营造了良好的营商环境。目前我国人民普遍存在硒摄入水平低下的现象(成人硒摄入量仅有26.36 μg/d,中国营养学会推荐的每日硒摄入量是60 μg/d),因此科学补硒、安全补硒、日常补硒对国民的身体健康十分重要。基于食用菌是耐硒和富硒的优良载体,结合我国是食用菌产量大国(占全球食用菌总产量的71.29%)的国情,为了响应人们目前对营养健康的诉求,研发创新型富硒食用菌类的功能健康食品刻不容缓。然而就现有的富硒技术,供应与之匹配的优质富硒食用菌产品还存在一些问题有待完善。
5.1 高品质的富硒食用菌开发
基于硒的吸收转化机制,研发新型的精准富硒调控技术,有助于提高食用菌中多糖、蛋白、多肽等功能因子的生物量和生物活性,降低重金属超标的风险,是破解我国食用菌产业发展困境的有效途径。
食用菌产品稳占市场的关键在于质量与安全,目前市面上的食用菌多为普通食用菌,富硒食用菌中高生物功能活性的有机硒占比低且伴生危害重金属存在超标的风险,高品质的富硒食用菌匮乏。富硒食用菌含有大量的硒多糖、硒蛋白、硒多肽等天然活性成分以及丰富的矿质元素,且已被证明具有抗炎、抗癌、抗氧化、降血脂、增强免疫力、改善人体血液微循环、预防疾病等促进健康的作用。除此之外,高品质的富硒食用菌极符合当下消费者对科学补充硒和保持健康的需求。开发高生物功能活性且安全的富硒食用菌,是顺应消费趋势的选择,其发展前景十分可观。
5.2 食用菌天然活性硒多糖、硒蛋白(多肽)等食疗营养素开发
开发高纯度的食用菌天然活性营养素,可增加其效价、产品价值及应用范围,是延长食用菌产业链的基础。
木耳、香菇等食用菌一直以可以清肠胃、减肥、补气血、预防心脑血管疾病等而闻名,但其具有的药用和食疗价值被严重低估。最近大量文献数据已确证食用菌天然活性硒多糖、硒蛋白、硒多肽等功能因子兼具硒和多糖或蛋白(多肽)的生理活性,在身体保健和食疗方面具有特别重要的作用。世界卫生组织(WHO)指出亚健康人群对天然活性营养素摄入不足导致难以达到防病抗病的目的。目前食疗营养素市场主要被欧美国家长期垄断占据,欧美国家用于生产辅助降血脂的食用菌保健品占整个食用菌保健品市场的30%,其次为有助于增强免疫力的食用菌保健品,市场占比为26%。近年来免疫调节剂硒多糖、抗氧化剂硒多肽受到了欧美国家的青睐,如美国GNC公司的Selenium Max Plus在2019年的销售额就已达到约3亿美元;同年英国Holland & Barrett公司推出的产品Selenium Complex with Vitamin E的销售额达到了约1.5亿英镑;其中美国GNC硒产品系列、美国Confidence信心康乐麦芽硒和澳洲herbsofgold硒片等占据了我国补硒剂消费市场的主要地位。目前市场上的食疗营养素主要以欧美品牌为主,我国食疗营养素的开发尚处于起步阶段。随着经济社会发展,人们对食疗营养素的需求日益增加,深度开发利用食用菌营养素势在必行,且将产生巨大的经济价值和社会效益。
5.3 益生菌发酵的活菌型富硒食用菌酱菜开发
采用Lactobacillus plantarum等益生菌发酵富硒食用菌酱菜,可改善食用菌酱菜的风味、功能特性等,为食用菌多元化加工提供新的思路。
随着人类对益生菌及其发酵产物与健康关系研究的不断深入,益生菌发酵食品受到了越来越多的关注。乳制品是益生菌目前应用最多、最为成熟的食品领域,但由于该类产品的高胆固醇和乳糖不耐受等限制性,开发新的非乳益生菌发酵产品是目前的研究热点。富硒食用菌富含生物功能活性物质,具有较高的生物效价,是益生菌发酵的理想基质和载体。近年来许多常见的食用菌原料如香菇、蘑菇等都被用于发酵产品的开发。通过多种益生菌混合发酵,可增加食用菌酱菜中利于人体吸收的活性和营养物质,可改善食用菌酱菜的口感和风味,还可补充人体的有益菌,从而对其感官与促进人体健康等功能特性产生积极影响。经益生菌发酵的食用菌酱菜营养和商业价值更高,能迎合素食主义者和乳糖不耐受等人群(该人群体内乳糖酶不足,而益生菌通过代谢产生乳糖酶能降低乳糖积累的程度,促进肠道的吸收和消化)的需求。目前,市面上已有发酵食用菌酱菜制品,但活菌型食用菌制品相对较少。益生菌发酵的活菌型富硒食用菌酱菜市场将进一步规范扩大。
6. 展望
近年来,在富硒食用菌对硒的吸收转化机制、富硒食用菌中硒的不同转化形态的生理活性、功能食品的开发等方面的研究取得了较多进展,但不管在基础研究上还是在产业化应用上都需要继续挖掘探索,主要可以集中在以下几方面:首先,有机硒在富硒食用菌中进行转化利用的机制尚不清晰,需要更深入地探究不同硒转化形态的代谢途径和调节机制。其次,虽然富硒食用菌中硒的不同转化形态的生物功能研究取得了进展,并且得到了许多潜在的免疫调节剂硒多糖和硒多肽以及抗氧化剂硒蛋白,但由于分离和提取高纯度的硒蛋白和硒多肽的技术限制性和复杂性,目前若要将其高效化、工业化应用于生产功能性食品、食疗营养素和辅助药物仍需要更多的探索和研究。最后,不同硒转化形态的生物活性与结构之间的构效关系仍不清楚,若能进一步阐明特定硒转化形态结构的功效,则能为机体吸收不同硒转化形态的代谢过程和机制提供更基础的理论依据。
-
![]()
图 1 硒在微生物中的吸收转化过程(食用菌为大型真菌)
注:APSe:5’-磷酸硒腺苷;GSH:谷胱甘肽;APS:5’-磷硫酸腺苷还原酶;H2Se:硒化氢。
Figure 1. Absorption and transformation process of selenium in microorganism (edible fungi are macrofungi)
![]()
图 2 硒转化形态(功能组分)的免疫活性
注:富硒多糖/富硒肽可通过刺激免疫细胞(分为发挥固有免疫和适应性免疫的细胞)释放细胞因子。其中,硒蛋白MSRB1是通过促进巨噬细胞中抗炎细胞因子的表达来发挥机体固有免疫功能;促炎细胞因子的大量合成(如IL-6、IL-1β)或抑制抗炎细胞因子释放(如IL-13、IL-4),会激活细胞内MAPK/NF-κB/STAT-3信号通路上相关蛋白的表达,使细胞产生病变,形成癌细胞,最终正常细胞被癌细胞格式化;释放抗炎细胞因子(如IL-13、IL-4)或抑制促炎细胞因子(如IL-6、IL-1β)的合成能表现出强大的抗炎活性,从而避免免疫反应发生以及癌细胞的形成,维持正常细胞的健康;抑制炎症自由基NO的合成能表现出强大的抗炎活性,而炎症自由基NO的大量释放会激活体内RAGE信号通路上相关蛋白的表达,使机体产生炎症反应(但当NO作为一种免疫信号时其效果反之)。
Figure 2. Immunoactivity of the transformed form of selenium
表 1 不同国家/地区人群的硒日摄入量(μg/d)
Table 1 Daily intake of selenium by population in different countries/regions (μg/d)
国家/地区 适宜人群 平均需要量
(Estimated average
requirement,EAR)推荐摄入量
(Recommend nutrient
intake,RNI)适宜摄入量
(Adequate intake,AI)可耐受最高摄入量
(Tolerable upper intake level,UL)参考文献 中国 婴儿 0~6月 / / 15 55 [10,12] 7~12月 / / 20 80 婴幼儿 1~6周岁 20~25 25~30 / 100~150 儿童 7~13周岁 35~45 40~55 / 200~300 青少年 14~17周岁 50 60 / 350 成人 正常人群 50 60~250 / 400 孕妇 / 50 / 400 乳母 / 65 / 400 苏联 成人 / 50 / 400 [13] 美国 成人 / 60~150 / 400 [14] 加拿大 成人 / 98~224 / 400 [15] 孟加拉 成人 / 63~122 / 400 [16] 注:1岁以下婴儿没有需要量研究资料,只能提出AI值。  下载: 导出CSV
下载: 导出CSV
表 2 食用菌中硒的主要形态
Table 2 Main forms of selenium in edible fungi
类别 名称 分子式 缩写 无机硒 元素硒 Se Se(0) 亚硒酸盐 SeO32− Se(IV) 硒酸盐 SeO42− Se(VI) 硒化物 Se2− Se(-II) 有机硒 低分子量 硒代蛋氨酸 C5H11NO2Se SeMet 硒代胱氨酸 C7H12O4Se2 SeCys2 硒代半胱氨酸 C3H7NO2Se SeCys 甲基硒代半胱氨酸 C4H9NO2Se MeSeCys 二甲基硒 C2H6Se DMSe 二甲基二硒 C2H6Se2 DMDSe 高分子量 含硒酶 / / 含硒蛋白 / /
下载: 导出CSV
表 3 代表性食用菌对硒的吸收和转化
Table 3 Absorption and transformation of selenium by representative edible fungi
食用菌种类 施硒方式 硒源 培养基中硒浓度 子实体硒含量(mg/kg) 主要硒形态组成/有机硒含量(mg/kg)/
有机硒转化率(%)参考文献 猴头菌 / 亚硒酸钠 12 mg/L 菌丝139.1 98.37% [61] 秀珍菇 / 亚硒酸钠 55 mg/kg 2.812 / [62] 大球盖菇 / 亚硒酸钠 150 mg/L 4727. 68 96. 27% [63] 黑木耳 / 生物纳米硒 0-250 mg/kg 0.03-85.70 / [64] / 亚硒酸钠 250 mg/kg >100 / 灵芝 / 亚硒酸钠 0~1.5 mmol/L 28~73 100% [47] 茶树菇 / 亚硒酸钠 0~1.5 mmol/L 12~34 84% [47] 巴西菇 / / / 9.15 SeCys2 5.734 mg/kg;
MeSeCys 1.84 mg/kg;SeMet 0.22 mg/kg[65] 平菇 / 富硒酵母 / 3 / [66] 拌料 亚硒酸钠 / 267.6 SeMet,SeCys2,Se(IV),Se(VI) [67] 香菇 浸泡 亚硒酸钠 / / Se(IV),SeMet,SeCys2 [52] 灌溉 亚硒酸钠 / / 无机硒,MeSeCys,SeCys,SeMet [53] 浸泡 硒酸钠 / 174.5 Se(VI),SeMet [67] 金针菇 拌料 亚硒酸钠 5 mg/kg 菌盖86.7
菌柄45.3/ [68] 双孢菇 拌料 富硒酵母 / 160 SeMet,Se(IV),SeMet,SeCys2, [50] 拌料 亚硒酸钠 / / Se(IV),SeCys2 [51] 喷施 亚硒酸钠 10~40 mg/kg 12.2~415 MeSeCys,SeCys2,SeMet [54] 灌溉 亚硒酸钠 10~40 mg/L 菌盖347
菌柄415无机硒,SeCys,MeSeCys,SeMet [54−55]
下载: 导出CSV
-
[1] SCHIAVON M, LIMA L W, JIANG Y, et al. Effects of selenium on plant metabolism and implications for crops and consumers[M]. Germany : Selenium in Plants, 2017:257−275.
[2] OROPEZA-MOE M, WISLØFF H, BERNHOFT A. Selenium deficiency associated porcine and human cardiomyopathies[J]. Journal of Trace Elements in Medicine and Biology,2015,31:148−156. doi: 10.1016/j.jtemb.2014.09.011
[3] ARTHUR J R, BROWN K M, FAIRWEATHER-TAIT S J, et al. Dietary selenium:Why do we need it and how much is enough?[J]. Nutrition and Food Science,1997,97(6):225−228. doi: 10.1108/00346659710180343
[4] ZHANG L, ZENG H, CHENG W H. Beneficial and paradoxical roles of selenium at nutritional levels of intake in healthspan and longevity[J]. Free Radical Biology and Medicine,2018,127:3−13. doi: 10.1016/j.freeradbiomed.2018.05.067
[5] RAYMAN M P. The importance of selenium to human health[J]. The Lancet,2000,356(9225):233−241. doi: 10.1016/S0140-6736(00)02490-9
[6] LENZ M, LENS P N L. The essential toxin:The changing perception of selenium in environmental sciences[J]. Science of the Total Environment,2009,407(12):3620−3633. doi: 10.1016/j.scitotenv.2008.07.056
[7] 黄冰霞, 支添添 , 赵志刚, 等. 硒元素与人类健康[J]. 宜春学院学报, 2019, 41(9):95−101. [HUANG Bingxia, ZHI Tiantian, ZHAO Zhigang, et al. Selenium and human health[J]. Journal of Yichun College, 2019, 41(9):95−101.] HUANG Bingxia, ZHI Tiantian, ZHAO Zhigang, et al. Selenium and human health[J]. Journal of Yichun College, 2019, 41(9): 95−101.
[8] NAVARRO-ALARCON M, CABRERA-VIQUE C. Selenium in food and the human body:A review[J]. Science of the Total Environment,2008,400(1-3):115−141. doi: 10.1016/j.scitotenv.2008.06.024
[9] TAMÁS M, MÁNDOKI Z, CSAPÓ J. The role of selenium content of wheat in the human nutrition. A literature review[J]. Agricultural and Food Sciences Acta Universitatis Sapientiae: Alimentaria,2010,3:5−34.
[10] MONSEN E R. Dietary reference intakes for the antioxidant nutrients:vitamin C, vitamin E, selenium, and carotenoids[J]. Journal of the Academy of Nutrition and Dietetics,2000,100(6):637.
[11] 黄峙. 食品硒源的生物学研究进展[J]. 食品科学,2001(5):90−94. [HUANG Z. Biological research progress of selenium sources in food[J]. Food Science,2001(5):90−94.] doi: 10.3321/j.issn:1002-6630.2001.05.030 HUANG Z. Biological research progress of selenium sources in food[J]. Food Science, 2001(5): 90−94. doi: 10.3321/j.issn:1002-6630.2001.05.030
[12] 中国营养学会. 中国居民膳食营养素参考摄入量2023版[M]. 北京:人民卫生出版社, 2023. [Chinese Nutrition Society. Reference Intake of Dietary Nutrients for Chinese residents 2023 edition[M]. Beijing:People's Medical Publishing House, 2023.] Chinese Nutrition Society. Reference Intake of Dietary Nutrients for Chinese residents 2023 edition[M]. Beijing: People's Medical Publishing House, 2023.
[13] GOLUBKINA N A, ALFTHAN G. Selenium status of pregnant women and newborns in the former Soviet Union[J]. Biological Trace Element Research,2002,89:13−23. doi: 10.1385/BTER:89:1:13
[14] HU X F, STRANGES S, CHAN H M. Abstract P220:Whole blood selenium concentration is inversely associated with prevalence of stroke:Results from the canadian health measures survey and the us national health and nutrition examine survey[J]. Circulation,2019,139(Suppl_1):AP220−AP220.
[15] Institute of Medicine (US) Panel on Dietary Antioxidants and Related Compounds. Dietary reference intakes for vitamin c, vitamin e, selenium, and carotenoids[M]. Washington (DC):National Academies Press (US), 2000.
[16] SPALLHOLZ J E, BOYLAN L M, RHAMAN M M. Environmental hypothesis:Is poor dietary selenium intake an underlying factor for arsenicosis and cancer in Bangladesh and West Bengal, India?[J]. Science of the Total Environment,2004,323(1−3):21−32. doi: 10.1016/j.scitotenv.2003.09.034
[17] 王岁楼, 胡巅, 李玉香. 活性干酵母对无机硒生物转化作用的研究[J]. 郑州工程学院学报,2001(2):55−58. [WANG Yalou, HU Dian, LI Yuxiang. Study on bio-transformation of inorganic selenium by active dry yeast[J]. Journal of Zhengzhou Institute of Engineering,2001(2):55−58.] WANG Yalou, HU Dian, LI Yuxiang. Study on bio-transformation of inorganic selenium by active dry yeast[J]. Journal of Zhengzhou Institute of Engineering, 2001(2): 55−58.
[18] 徐辉碧, 黄开勋. 硒的化学、生物化学及其在生命科学中的应用[M]. 武汉:华中理工大学出版社, 1994. [XU Huibi, HUANG Kaixun. Chemistry and biochemistry of Selenium and its application in Life science[M]. Wuhan:Huazhong University of Science and Technology Press, 1994.] XU Huibi, HUANG Kaixun. Chemistry and biochemistry of Selenium and its application in Life science[M]. Wuhan: Huazhong University of Science and Technology Press, 1994.
[19] 邹小智, 康德灿, 罗佳, 等. 食药用菌富硒食品的研发概况[J]. 食用菌,2022,44(3):1−3,8. [ZOU Xiaozhi, KANG Dechan, LUO Jia, et al. Overview of research and development of selenium-enriched foods from medicinal mushrooms[J]. Edible Mushroom,2022,44(3):1−3,8.] doi: 10.3969/j.issn.1000-8357.2022.03.001 ZOU Xiaozhi, KANG Dechan, LUO Jia, et al. Overview of research and development of selenium-enriched foods from medicinal mushrooms[J]. Edible Mushroom, 2022, 44(3): 1−3,8. doi: 10.3969/j.issn.1000-8357.2022.03.001
[20] 刘莉, 刘毅. 微量元素硒与人体健康[J]. 黑龙江科学,2014,5(6):16−17. [LIU Li, LIU Yi. Trace element selenium and human health[J]. Heilongjiang Science,2014,5(6):16−17.] doi: 10.3969/j.issn.1674-8646.2014.06.005 LIU Li, LIU Yi. Trace element selenium and human health[J]. Heilongjiang Science, 2014, 5(6): 16−17. doi: 10.3969/j.issn.1674-8646.2014.06.005
[21] ULLAH H, LIU G, YOUSAF B, et al. A comprehensive review on environmental transformation of selenium:Recent advances and research perspectives[J]. Environmental Geochemistry and Health,2019,41:1003−1035. doi: 10.1007/s10653-018-0195-8
[22] DUMONT E, VANHAECKE F, CORNELIS R. Selenium speciation from food source to metabolites:A critical review[J]. Analytical and Bioanalytical Chemistry,2006,385:1304−1323. doi: 10.1007/s00216-006-0529-8
[23] 刘培棣. 硒资源及其综合开发利用[M]. 北京:中国科学技术出版社, 1993:65−71. [LIU Peidi. Selenium resources and their comprehensive development and Utilization[M]. Beijing:Science and Technology Press of China, 1993:65−71.] LIU Peidi. Selenium resources and their comprehensive development and Utilization[M]. Beijing: Science and Technology Press of China, 1993: 65−71.
[24] KESSI J, HANSELMANN K W. Similarities between the abiotic reduction of selenite with glutathione and the dissimilatory reaction mediated by Rhodospirillum rubrum and Escherichia coli[J]. Journal of Biological Chemistry,2004,279(49):50662−50669. doi: 10.1074/jbc.M405887200
[25] KIELISZEK M, BłAżEJAK S, GIENTKA I, et al. Accumulation and metabolism of selenium by yeast cells[J]. Applied Microbiology and Biotechnology,2015,99:5373−5382. doi: 10.1007/s00253-015-6650-x
[26] WEI R, ZIMMERMANN W. Biocatalysis as a green route for recycling the recalcitrant plastic polyethylene terephthalate[J]. Microbial Biotechnology,2017,10(6):1302. doi: 10.1111/1751-7915.12714
[27] 朱燕云, 吴文良, 赵桂慎, 等. 硒在动植物及微生物体中的转化规律研究进展[J]. 农业资源与环境学报,2018,35(3):189−198. [ZHU Yanyun, WU Wenliang, ZHAO Guishen, et al. Research progress on the transformation of selenium in plants, animals and microorganisms[J]. Journal of Agricultural Resources and Environment,2018,35(3):189−198.] ZHU Yanyun, WU Wenliang, ZHAO Guishen, et al. Research progress on the transformation of selenium in plants, animals and microorganisms[J]. Journal of Agricultural Resources and Environment, 2018, 35(3): 189−198.
[28] 肖竞, 周德平. 酵母对微量元素硒的生物富集及应用[J]. 中国饲料,2004(19):10−11,26. [XIAO Jing, ZHOU Deping. Bioenrichment and application of trace element selenium by yeast[J]. China Feed,2004(19):10−11,26.] doi: 10.3969/j.issn.1004-3314.2004.19.007 XIAO Jing, ZHOU Deping. Bioenrichment and application of trace element selenium by yeast[J]. China Feed, 2004(19): 10−11,26. doi: 10.3969/j.issn.1004-3314.2004.19.007
[29] LAZARD M, BLANQUET S, FISICARO P, et al. Uptake of selenite by Saccharomyces cerevisiae involves the high and low affinity orthophosphate transporters[J]. Journal of Biological Chemistry,2010,285(42):32029−32037. doi: 10.1074/jbc.M110.139865
[30] MCDERMOTT J R, ROSEN B P, LIU Z. Jen1p:A high affinity selenite transporter in yeast[J]. Molecular Biology of the Cell,2010,21(22):3934−3941. doi: 10.1091/mbc.e10-06-0513
[31] TURNER R J, WEINER J H, TAYLOR D E. Selenium metabolism in Escherichia coli[J]. Biometals,1998,11:223−227. doi: 10.1023/A:1009290213301
[32] FALANDYSZ J. Selenium in edible mushrooms[J]. Journal of Environmental Science and Health Part C,2008,26(3):256−299. doi: 10.1080/10590500802350086
[33] SERAFIN MUÑOZ A H, KUBACHKA K, WROBEL K, et al. Se-enriched mycelia of Pleurotus ostreatus:Distribution of selenium in cell walls and cell membranes/cytosol[J]. Journal of Agricultural and Food Chemistry,2006,54(9):3440−3444. doi: 10.1021/jf052973u
[34] WANG J, WANG B, ZHANG D, et al. Selenium uptake, tolerance and reduction in Flammulina velutipes supplied with selenite[J]. Peer J,2016,4:e1993. doi: 10.7717/peerj.1993
[35] KAUR T, VASHISHT A, PRAKASH N T, et al. Role of selenium-tolerant fungi on plant growth promotion and selenium accumulation of maize plants grown in seleniferous soils[J]. Water, Air, and Soil Pollution,2022,233:1−12. doi: 10.1007/s11270-021-05470-z
[36] MILETIĆ D, PANTIĆ M, SKNEPNEK A, et al. Influence of selenium yeast on the growth, selenium uptake and mineral composition of Coriolus versicolor mushroom[J]. Journal of Basic Microbiology,2020,60(4):331−340. doi: 10.1002/jobm.201900520
[37] SILVA M C S, LUZ J M R, PAIVA A P S, et al. Growth and tolerance of Pleurotus ostreatus at different selenium forms[J]. Journal of Agricultural Science,2019,11(2):151. doi: 10.5539/jas.v11n2p151
[38] NIE X, YANG X, HE J, et al. Bioconversion of inorganic selenium to less toxic selenium forms by microbes:A review[J]. Frontiers in Bioengineering and Biotechnology,2023,11:1167123. doi: 10.3389/fbioe.2023.1167123
[39] ZHAO C, LI J, CHEN N, et al. Antioxidant activity and transcriptomic analysis of Se-enriched golden oyster mushroom Pleurotus citrinopileatus (agaricomycetes)[J]. International Journal of Medicinal Mushrooms, 2020, 22(8).
[40] XU M, ZHU S, LI Y, et al. Effect of selenium on mushroom growth and metabolism:A review[J]. Trends in Food Science and Technology,2021,118:328−340. doi: 10.1016/j.jpgs.2021.10.018
[41] WANG H, YANG S, CHEN Y, et al. Comprehensive distribution and species of selenium in Se-enriched Pichia kudriavzevii 1845[J]. Food Chemistry, 2023:137966.
[42] OGRA Y, SHIMIZU M, TAKAHASHI K, et al. Biotransformation of organic selenium compounds in budding yeast, Saccharomyces cerevisiae[J]. Metallomics,2018,10(9):1257−1263. doi: 10.1039/C8MT00176F
[43] 郑世学, 粟静, 王瑞, 等. 硒是双刃剑?—谈微生物中的硒代谢[J]. 华中农业大学学报, 2013, 32 (5):1−8. [ZHENG Shixue, SU Jing, WANG Rui, et al. Selenium is a double-edged sword? — Selenium metabolism in microorganisms[J]. Journal of Huazhong Agricultural University, 2013, 32(5):1−8.] ZHENG Shixue, SU Jing, WANG Rui, et al. Selenium is a double-edged sword? — Selenium metabolism in microorganisms[J]. Journal of Huazhong Agricultural University, 2013, 32(5): 1−8.
[44] WANG Q, KONG L, HUANG Q, et al. Uptake and translocation mechanisms of different forms of organic selenium in rice (Oryza satival.)[J]. Frontiers in Plant Science,2022,13:970480. doi: 10.3389/fpls.2022.970480
[45] ABRAMS M M, BURAU R G, ZASOSKI R J. Organic selenium distribution in selected California soils[J]. Soil Science Society of America Journal,1990,54(4):979−982. doi: 10.2136/sssaj1990.03615995005400040007x
[46] ZHANG L, SONG H, GUO Y, et al. Benefit–risk assessment of dietary selenium and its associated metals intake in China (2017-2019):Is current selenium-rich agro-food safe enough?[J]. Journal of Hazardous Materials,2020,398:123224. doi: 10.1016/j.jhazmat.2020.123224
[47] NIEDZIELSKI P, MLECZEK M, SIWULSKI M, et al. Efficacy of supplementation of selected medicinal mushrooms with inorganic selenium salts[J]. Journal of Environmental Science and Health, Part B,2014,49(12):929−937. doi: 10.1080/03601234.2014.951576
[48] LI X, YAN L, LI Q, et al. Transcriptional profiling of Auricularia cornea in selenium accumulation[J]. Scientific Reports,2019,9(1):1−13. doi: 10.1038/s41598-018-37186-2
[49] HU T, LI L, HUI G, et al. Selenium biofortification and its effect on multi-element change in Auricularia auricular[J]. Food Chemistry,2019,295:206−213. doi: 10.1016/j.foodchem.2019.05.101
[50] DERNOVICS M, STEFÁNKA Z, FODOR P. Improving selenium extraction by sequential enzymatic processes for Se-speciation of selenium-enriched Agaricus bisporus[J]. Analytical and Bioanalytical Chemistry,2002,372:473−480. doi: 10.1007/s00216-001-1215-5
[51] STEFÁNKA Z, IPOLYI I, DERNOVICS M, et al. Comparison of sample preparation methods based on proteolytic enzymatic processes for Se-speciation of edible mushroom (Agaricus bisporus) samples[J]. Talanta,2001,55(3):437−447. doi: 10.1016/S0039-9140(01)00398-8
[52] YOSHIDA M, SUGIHARA S, INOUE Y, et al. Composition of chemical species of selenium contained in selenium-enriched shiitake mushroom and vegetables determined by high performance liquid chromatography with inductively coupled plasma mass spectrometry[J]. Journal of Nutritional Science and Vitaminology,2005,51(3):194−199. doi: 10.3177/jnsv.51.194
[53] GERGELY V, KUBACHKA K M, MOUNICOU S, et al. Selenium speciation in Agaricus bisporus and Lentinula edodes mushroom proteins using multi-dimensional chromatography coupled to inductively coupled plasma mass spectrometry[J]. Journal of Chromatography A,2006,1101(1-2):94−102. doi: 10.1016/j.chroma.2005.09.061
[54] MASEKO T, CALLAHAN D L, DUNSHEA F R, et al. Chemical characterisation and speciation of organic selenium in cultivated selenium-enriched Agaricus bisporus[J]. Food Chemistry,2013,141(4):3681−3687. doi: 10.1016/j.foodchem.2013.06.027
[55] CREMADES O, DIAZ-HERRERO M M, CARBONERO-AGUILAR P, et al. Preparation and characterisation of selenium-enriched mushroom aqueous enzymatic extracts (MAEE) obtained from the white button mushroom (Agaricus bisporus)[J]. Food Chemistry,2012,133(4):1538−1543. doi: 10.1016/j.foodchem.2012.02.046
[56] ROS G H, VAN ROTTERDAM A M D, BUSSINK D W, et al. Selenium fertilization strategies for bio-fortification of food:An agro-ecosystem approach[J]. Plant and Soil,2016,404:99−112. doi: 10.1007/s11104-016-2830-4
[57] ZHANG H, ZHAO Z, ZHANG X, et al. Effects of foliar application of selenate and selenite at different growth stages on selenium accumulation and speciation in potato (Solanum tuberosum L.)[J]. Food Chemistry,2019,286:550−556. doi: 10.1016/j.foodchem.2019.01.185
[58] 戴志华. 水稻对硒的吸收转化及调控机理研究[D]. 武汉:华中农业大学, 2020. [DAI Zhihua. Study on the absorption, transformation and regulation mechanism of Selenium in rice[D]. Wuhan:Huazhong Agricultural University, 2020.] DAI Zhihua. Study on the absorption, transformation and regulation mechanism of Selenium in rice[D]. Wuhan: Huazhong Agricultural University, 2020.
[59] ARVY M P, THIERSAULT M, DOIREAU P. Relationships between selenium, micronutrients, carbohydrates, and alkaloid accumulation in Catharanthus roseus cells[J]. Journal of Plant Nutrition,1995,18(8):1535−1546. doi: 10.1080/01904169509365002
[60] DENG X, LIU K, LI M, et al. Difference of selenium uptake and distribution in the plant and selenium form in the grains of rice with foliar spray of selenite or selenate at different stages[J]. Field Crops Research,2017,211:165−171. doi: 10.1016/j.fcr.2017.06.008
[61] 姜宁, 朱淑彤, 苏浩雨, 等. 猴头菌菌丝富硒特性及富硒深层发酵的研究[J]. 天然产物研究与开发,2021,33(5):810−818,830. [JIANG Ning, Zhu Shutong, Su Haoyu, et al. Study on selenium-rich characteristics and deep fermentation of Hericium mycelium[J]. Natural Products Research and Development,2021,33(5):810−818,830.] JIANG Ning, Zhu Shutong, Su Haoyu, et al. Study on selenium-rich characteristics and deep fermentation of Hericium mycelium[J]. Natural Products Research and Development, 2021, 33(5): 810−818,830.
[62] 吕欣彧. 富硒食用菌最新研究进展[J]. 现代食品,2022,28(13):29−33. [LU Xinyu. The latest research progress of selenium-rich edible fungi[J]. Modern Food,2022,28(13):29−33.] LU Xinyu. The latest research progress of selenium-rich edible fungi[J]. Modern Food, 2022, 28(13): 29−33.
[63] 鲍晓天. 皱环球盖菇及其栽培[J]. 农技服务, 1997(8):27−28. [BAO Xiaotian. Mushroom rugosus and its cultivation, 1997(8):27−28.] BAO Xiaotian. Mushroom rugosus and its cultivation, 1997(8): 27−28.
[64] 郭慧阳, 王彤, 胡惠萍, 等. 富集微量元素食用菌的研究进展[J]. 中国食用菌,2022,41(4):1−8. [GUO Huiyang, WANG Tong, HU Huiping, et al. Research progress on enrichment of trace elements in edible fungi[J]. Chinese Edible Fungi,2022,41(4):1−8.] GUO Huiyang, WANG Tong, HU Huiping, et al. Research progress on enrichment of trace elements in edible fungi[J]. Chinese Edible Fungi, 2022, 41(4): 1−8.
[65] HU Z, YAO Y, LÜ M, et al. Isolation and identification of three water-soluble selenoproteins in Se-enriched Agaricus blazei Murrill[J]. Food Chemistry,2021,344:128691. doi: 10.1016/j.foodchem.2020.128691
[66] WANG D, WANG J, LIU H, et al. The main structural unit elucidation and immunomodulatory activity in vitro of a selenium-enriched polysaccharide produced by Pleurotus ostreatus[J]. Molecules,2022,27(8):2591. doi: 10.3390/molecules27082591
[67] ASSUNÇÃO L S, SILVA M C S, FERNANDEZ M G, et al. Speciation of selenium in Pleurotus ostreatus and Lentinula edodes mushrooms[J]. Journal of Biotechnology Letters,2014,5(1):79−86.
[68] TIE M, LI B, SUN T, et al. HPLC-ICP-MS speciation of selenium in Se-cultivated Flammulina velutipes[J]. Arabian Journal of Chemistry,2020,13(1):416−422. doi: 10.1016/j.arabjc.2017.05.012
[69] 兰吉玉, 潘承丹. 黔东南州食用菌重金属污染状况及质量安全评价[J]. 农技服务,2023,40(5):100−103. [LAN Jiyu, PAN Chengdan. Heavy metal pollution status and quality safety evaluation of edible fungi in Qiandongnan prefecture[J]. Agricultural Technology Service,2023,40(5):100−103.] doi: 10.3969/j.issn.1004-8421.2023.5.njfw202305027 LAN Jiyu, PAN Chengdan. Heavy metal pollution status and quality safety evaluation of edible fungi in Qiandongnan prefecture[J]. Agricultural Technology Service, 2023, 40(5): 100−103. doi: 10.3969/j.issn.1004-8421.2023.5.njfw202305027
[70] 陈明杰, 宋盼盼, 赵妍, 等. 食用菌重金属污染及防控措施研究进展[J]. 分子植物育种,2023,21(5):1733−1740. [CHEN Mingjie, SONG Panpan, ZHAO Yan, et al. Research progress of heavy metal pollution of edible fungi and its prevention and control measures[J]. Molecular Plant Breeding,2023,21(5):1733−1740.] CHEN Mingjie, SONG Panpan, ZHAO Yan, et al. Research progress of heavy metal pollution of edible fungi and its prevention and control measures[J]. Molecular Plant Breeding, 2023, 21(5): 1733−1740.
[71] 刘思洁, 姜楠, 郭金芝. 吉林省干制食用菌中4种重金属污染状况监测及暴露风险评估[J]. 中国地方病防治,2023,38(1):18−21,24. [LIU Sijie, JIANG Nan, GUO Jinzhi. Pollution monitoring and exposure risk assessment of 4 heavy metals in dried edible fungi in Jilin Province[J]. Chinese Endemic Disease Control,2023,38(1):18−21,24.] LIU Sijie, JIANG Nan, GUO Jinzhi. Pollution monitoring and exposure risk assessment of 4 heavy metals in dried edible fungi in Jilin Province[J]. Chinese Endemic Disease Control, 2023, 38(1): 18−21,24.
[72] PAK S J, CHEN F, MA L, et al. Functional perspective of black fungi (Auricularia auricula):Major bioactive components, health benefits and potential mechanisms[J]. Trends in Food Science and Technology,2021,114:245−261. doi: 10.1016/j.jpgs.2021.05.013
[73] LI Q, CHEN G, CHEN H, et al. Se-enriched G. frondosa polysaccharide protects against immunosuppression in cyclophosphamide-induced mice via MAPKs signal transduction pathway[J]. Carbohydrate Polymers,2018,196:445−456. doi: 10.1016/j.carbpol.2018.05.046
[74] ZHENG Y, WANG W, LI Y. Antitumor and immunomodulatory activity of polysaccharide isolated from Trametes orientalis[J]. Carbohydrate Polymers,2015,131:248−254. doi: 10.1016/j.carbpol.2015.05.074
[75] 连科迅. 硒化新疆乌拉尔甘草多糖抗炎免疫调节作用研究[D]. 石河子:石河子大学, 2018. [LIAN Kosen. Study on anti-inflammatory immunomodulatory effects of selenized Glycyrrhiza uralensis polysaccharide[D]. Shihezi:Shihezi University, 2018.] LIAN Kosen. Study on anti-inflammatory immunomodulatory effects of selenized Glycyrrhiza uralensis polysaccharide[D]. Shihezi: Shihezi University, 2018.
[76] LI Q, ZHAO T, MAO G, et al. A Se-enriched Grifola frondosa polysaccharide induces macrophage activation by TLR4-mediated MAPK signaling pathway[J]. International Journal of Biological Macromolecules,2023,238:124108. doi: 10.1016/j.ijbiomac.2023.124108
[77] WANG J, ZHANG T, LIU X, et al. Aqueous extracts of Se-enriched Auricularia auricular attenuates D-galactose-induced cognitive deficits, oxidative stress and neuroinflammation via suppressing RAGE/MAPK/NF-κB pathway[J]. Neuroscience Letters, 2019.
[78] INGOLD I, BERNDT C, SCHMITT S, et al. Selenium utilization by GPX4 is required to prevent hydroperoxide-induced ferroptosis[J]. Cell,2018,172(3):409−422. doi: 10.1016/j.cell.2017.11.048
[79] 孙纳新, 王成华. 硒蛋白及其对巨噬细胞功能调节作用的研究进展[J]. 食品科学,2012,33(19):363−366. [SUN Naxin, WANG Chenghua. Research progress of selenoproteins and their regulatory effects on macrophage function[J]. Food Science,2012,33(19):363−366.] SUN Naxin, WANG Chenghua. Research progress of selenoproteins and their regulatory effects on macrophage function[J]. Food Science, 2012, 33(19): 363−366.
[80] AVERY J C, HOFFMANN P R. Selenium, selenoproteins, and immunity[J]. Nutrients,2018,10(9):1203. doi: 10.3390/nu10091203
[81] ZHANG Z H, LIAO T T, DENG C M, et al. Purification and characterization of Se-enriched Grifola frondosa glycoprotein, and evaluating its amelioration effect on As3+-induced immune toxicity[J]. Journal of the Science of Food and Agriculture,2022,102(6):2526−2537. doi: 10.1002/jsfa.11594
[82] LIU D, LIU Y, HUANG P, et al. Polysaccharide from natural Cordyceps sinensis ameliorated intestinal injury and enhanced antioxidant activity in immunosuppressed mice[J]. Journal of Agricultural and Food Chemistry, 67(4):1138−1147.
[83] ZHANG J, GAO S, LI H, et al. Immunomodulatory effects of selenium-enriched peptides from soybean in cyclophosphamide-induced immunosuppressed mice[J]. Food Science and Nutrition,2021,9(11):6322−6334. doi: 10.1002/fsn3.2594
[84] HE T, YANG H, ZHANG L, et al. Antioxidant activity and inhibition of ultraviolet radiation-induced skin damage of Selenium-rich peptide fraction from selenium-rich yeast protein hydrolysate[J]. Food Chemistry, 2020, 178:2−7.
[85] 胡居吾. 硒在农作物生长过程的富集及大豆硒蛋白生物功效研究[D]. 南昌:南昌大学, 2017:68−88. [HU Juwu. Studies on the accumulation of selenium in the growing process of crops and the biological efficacy of soybean selenoprotein[D]. Nanchang:Nanchang University, 2017:68−88.] HU Juwu. Studies on the accumulation of selenium in the growing process of crops and the biological efficacy of soybean selenoprotein[D]. Nanchang: Nanchang University, 2017: 68−88.
[86] FANG Y, PAN X, ZHAO E, et al. Isolation and identification of immunomodulatory selenium-containing peptides from selenium-enriched rice protein hydrolysates[J]. Food Chemistry,2019,275:696−702. doi: 10.1016/j.foodchem.2018.09.115
[87] CHEN C, ZHANG Y, LI Y, et al. In vivo and in vitro anti-tumour response of selenium-protein polysaccharide extracted from rich selenium Agaricus blazei[J]. Food and Agricultural Immunology,2007,18(2):139−149. doi: 10.1080/09540100701616548
[88] MA L, LIU J, LIU A, et al. Cytoprotective effect of selenium polysaccharide from Pleurotus ostreatus against H2O2-induced oxidative stress and apoptosis in PC12 cells[J]. Arabian Journal of Chemistry,2022,15(4):103686. doi: 10.1016/j.arabjc.2022.103686
[89] 陈彻. 富硒姬松茸提取物硒蛋白多糖在体内与体外的抗肿瘤效果研究[D]. 兰州:兰州大学, 2008. [CHEN Che. Study on the antitumor effect of selenium proteoglycan from selenium-enriched Agaricus blazei Murrill extract in vivo and in vitro[D]. Lanzhou:Lanzhou University, 2008.] CHEN Che. Study on the antitumor effect of selenium proteoglycan from selenium-enriched Agaricus blazei Murrill extract in vivo and in vitro[D]. Lanzhou: Lanzhou University, 2008.
[90] LIU Y, YOU Y, LI Y, et al. The characterization, selenylation and antidiabetic activity of mycelial polysaccharides from Catathelasma ventricosum[J]. Carbohydrate Polymers,2017,174:72−81. doi: 10.1016/j.carbpol.2017.06.050
[91] WANG J, YANG X, BAO A, et al. Microwave-assisted synthesis, structure and anti-tumor activity of selenized Artemisia sphaerocephala polysaccharide[J]. International Journal of Biological Macromolecules,2017,95:1108−1118. doi: 10.1016/j.ijbiomac.2016.10.101
[92] WEI Z, YI Y, LUO Z, et al. Selenopeptide nanomedicine activates natural killer cells for enhanced tumor chemoimmunotherapy[J]. Advanced Materials,2022,34(17):2108167. doi: 10.1002/adma.202108167
[93] GU Y G, QIU Y, YUAN Y H, et al. Characterization of selenium-containing polysaccharides isolated from selenium-enriched tea and its bioactivities[J]. Food Chemistry,2020,316:126371. doi: 10.1016/j.foodchem.2020.126371
[94] CHEN S, SU T, WANG Z. Structural characterization, antioxidant activity, and immunological activity in vitro of polysaccharides from fruiting bodies of Suillus granulatus[J]. Journal of Food Biochemistry,2018,42(3):e12515. doi: 10.1111/jfbc.12515
[95] GU H, LIANG L, PENG Z X, et al. Optimization of enzymatic extraction, characterization and bioactivities of Se-polysaccharides from Se-enriched Lentinus edodes[J]. Food Bioscience,2023,51:102346. doi: 10.1016/j.fbio.2022.102346
[96] 赵镭. 灵芝生物富硒及富硒灵芝硒蛋白的分离纯化和抗氧化性研究[D]. 北京:中国农业大学, 2004. [ZHAO Lei. Study on Se-enriched Ganoderma lucidum and isolation, purification and antioxidant activity of selenium protein from Ganoderma lucidum [D]. Beijing:China Agricultural University, 2004.] ZHAO Lei. Study on Se-enriched Ganoderma lucidum and isolation, purification and antioxidant activity of selenium protein from Ganoderma lucidum [D]. Beijing: China Agricultural University, 2004.
[97] 张强, 王松华, 孙玉军, 等. 羊肚菌蛋白的硒化修饰及其体外抗氧化活性[J]. 精细化工,2017,34(11):1252−1259. [ZHANG Qiang, WANG Songhua, SUN Yujun, et al. Selenium modification and antioxidant activity of Morchella protein in vitro[J]. Fine Chemical Industry,2017,34(11):1252−1259.] ZHANG Qiang, WANG Songhua, SUN Yujun, et al. Selenium modification and antioxidant activity of Morchella protein in vitro[J]. Fine Chemical Industry, 2017, 34(11): 1252−1259.
[98] ZHAO J, ZHAO H, WANG X, et al. Characteristics and enhanced antioxidant activity of egg white protein selenized by dry-heating in the presence of selenite[J]. Journal of Agricultural and Food Chemistry,2013,61(12):3131−3139. doi: 10.1021/jf305166z
[99] SCHOMBURG L. Selenoprotein P–Selenium transport protein, enzyme and biomarker of selenium status[J]. Free Radical Biology and Medicine,2022,191:150−163. doi: 10.1016/j.freeradbiomed.2022.08.022
[100] GUO H, GUO S, LIU H. Antioxidant activity and inhibition of ultraviolet radiation-induced skin damage of Selenium-rich peptide fraction from selenium-rich yeast protein hydrolysate[J]. Bioorganic Chemistry,2020,105:104431. doi: 10.1016/j.bioorg.2020.104431
[101] AGRAWAL H, JOSHI R, GUPTA M. Isolation, purification and characterization of antioxidative peptide of pearl millet (Pennisetum glaucum) protein hydrolysate[J]. Food Chemistry,2016,204:365−372. doi: 10.1016/j.foodchem.2016.02.127
-
期刊类型引用(5)
1. 胡平国,李霞雪,杨巧慧,曹璐,陈志民,陈亚,曾艳. 甜樱桃中多菌灵等6种农药的残留检测方法建立及残留消解动态研究. 农产品质量与安全. 2025(01): 86-91 .  百度学术
百度学术
2. 黄敏君,黄靓,黄亚,朱小芳,关培培,王静. LC-MS/MS法检测棉花7种杀虫剂残留研究进展. 纺织报告. 2024(09): 17-19+33 . 百度学术
3. 罗雪燕,陈宝欣,黄柳霞,林洁娜,贝荣廷. 研究不同前处理方法对蔬菜中21种农药残留量的影响. 中外食品工业. 2024(14): 46-50 . 百度学术
4. 邓航,朱礼,张可欣,袁列江,李政,吴海智,上官佳,陈瑶,薛敏敏,王秀,李格,孙逸清. QuEChERS净化结合高效液相色谱串联质谱法快速测定果蔬中四唑虫酰胺残留. 农药. 2024(12): 904-910+924 . 百度学术
5. 李艳霞,皮大能. QuEChERS-气相色谱-三重四级杆串联质谱法同时测定青贮玉米中24种农药残留. 中国饲料. 2024(23): 315-322 . 百度学术
其他类型引用(1)
 下载:
下载:


计量
- 文章访问数: 186
- HTML全文浏览量: 70
- PDF下载量: 30
- 被引次数: 6



