


Progress on the Improvement of Quality and Functional Properties of Fermented Milk by Complex Strains of Bacteria
-
摘要: 发酵乳具有悠久的历史,是经乳酸菌发酵而成的,富含蛋白质、矿物质和维生素等多种营养成分。随着人们对生活品质追求的提高,发酵乳受到消费者越来越多的关注。目前市面上的发酵乳多数是以单菌株或传统乳酸菌(嗜热链球菌和保加利亚乳杆菌)作为发酵剂制备而成的,但是以此生产发酵乳会有发酵时间过长、口感略差和稳定性不佳等问题。近年来,复配菌株作为发酵乳的发酵剂成为发酵乳制备方向的研究热点,利用不同菌株间的互作协同作用,可以有效提高代谢产物的含量和丰富度,进而改善发酵乳的品质及功能特性,弥补传统发酵乳在产品形态以及感官体验等方面的不足。本文综述了复配菌株能够加快发酵乳pH下降速度,丰富产品感官以及改善其流变特性和持水能力,同时还可以增强发酵乳在降血脂、抗炎、抗氧化、抑菌等功能方面的能力。最后展望了发酵乳在未来可以进行创新的研究方向,为今后发酵乳的多元化、功能化、精准化生产提供指导。Abstract: Fermented milk has a long history of being fermented by lactic acid bacteria. Fermented milk contains many elements, including protein, minerals, and vitamins. Fermented milk is gaining more and more attention from customers as people’s desire for a high quality of life improves. Most of the fermented milk on the market today are prepared with single strains or traditional lactic acid bacteria (Streptococcus thermophilus and Lactobacillus bulgaricus) as fermenting agents. However, this production method results in issues like an excessively long fermentation time, a mildly inferior taste, and poor stability. Compound strains have recently gained attention in the field of fermented milk preparation. By utilizing interactions and synergies between various strains, it is possible to increase the quantity and diversity of metabolites, enhancing the quality and functional properties of fermented milk and compensating for some of the shortcomings of conventional fermented milk in terms of product morphology and sensory experience. This study examines how complexing strains have accelerated pH reduction, improved the product’s sensory qualities, rheological characteristics, and water-holding capacity, as well as increased their capacity for lipid-lowering, anti-inflammatory, antioxidant, and bacteriostatic effects. Finally, the future research paths for fermented milk innovation are intended to offer suggestions for the diverse, functionalized, and precise production of fermented milk.
-
Keywords:
- strain /
- fermented milk /
- physicochemical properties /
- functional properties
-
世界卫生组织、联合国粮农组织和国际乳品联合会将发酵乳定义为:在含有(或不含)乳粉(或脱脂乳粉)的乳中(杀菌乳或浓缩乳),添加乳酸菌(Lactic acid bacteria,LAB)进行乳酸发酵制成的凝乳状产品[1]。我国也于2010年发布相关国家标准,将以生牛(羊)乳或乳粉为原料,经杀菌、发酵后制成的pH降低的产品定义为发酵乳[2]。根据保藏温度可以将发酵乳分为常温和低温发酵乳,根据发酵乳的口味,还可以分为原味发酵乳和风味发酵乳。风味发酵乳的出现主要是因为消费者追求不断改变,目前市售的风味发酵乳中有添加苹果、枸杞、芒果、草莓、凤梨等原料,口味众多,能够满足不同人群的喜好,逐渐成为最受欢迎的发酵乳品类。表1介绍了这四种发酵乳的特点、优势及不足。
表 1 不同类型发酵乳的特点、优势及不足Table 1. Characteristics, advantages and disadvantages of different types of fermented milk分类标准 发酵乳种类 特点 优势 不足 保藏温度 常温发酵乳[3] 生产过程中进行二次巴氏杀菌 可以在4~25 ℃保存5~6个月,便于长途运输;携带方便;保质期长 营养创新被限制,目前只能向口味创新、饮品和甜品化方向发展[4] 低温发酵乳[5] 需要在2~6 ℃储存,生产过程中只进行一次巴氏杀菌 存有活性乳酸菌,可以促进人体肠道消化、调节肠道菌群健康 保存时间有限;储存要求高;
销售成本高口感 原味发酵乳 保留传统发酵乳最醇正的口感风味 无其它物质添加,较风味发酵乳更加安全 口味单一,不能满足消费者对于商品多元化的需求 风味发酵乳 除乳或乳粉外,同时可以添加其它原料,选择性添加食品添加剂、营养强化剂、果蔬、谷物等 口味丰富,满足不同人群的感官要求 产品质量参差不一;额外添加成分
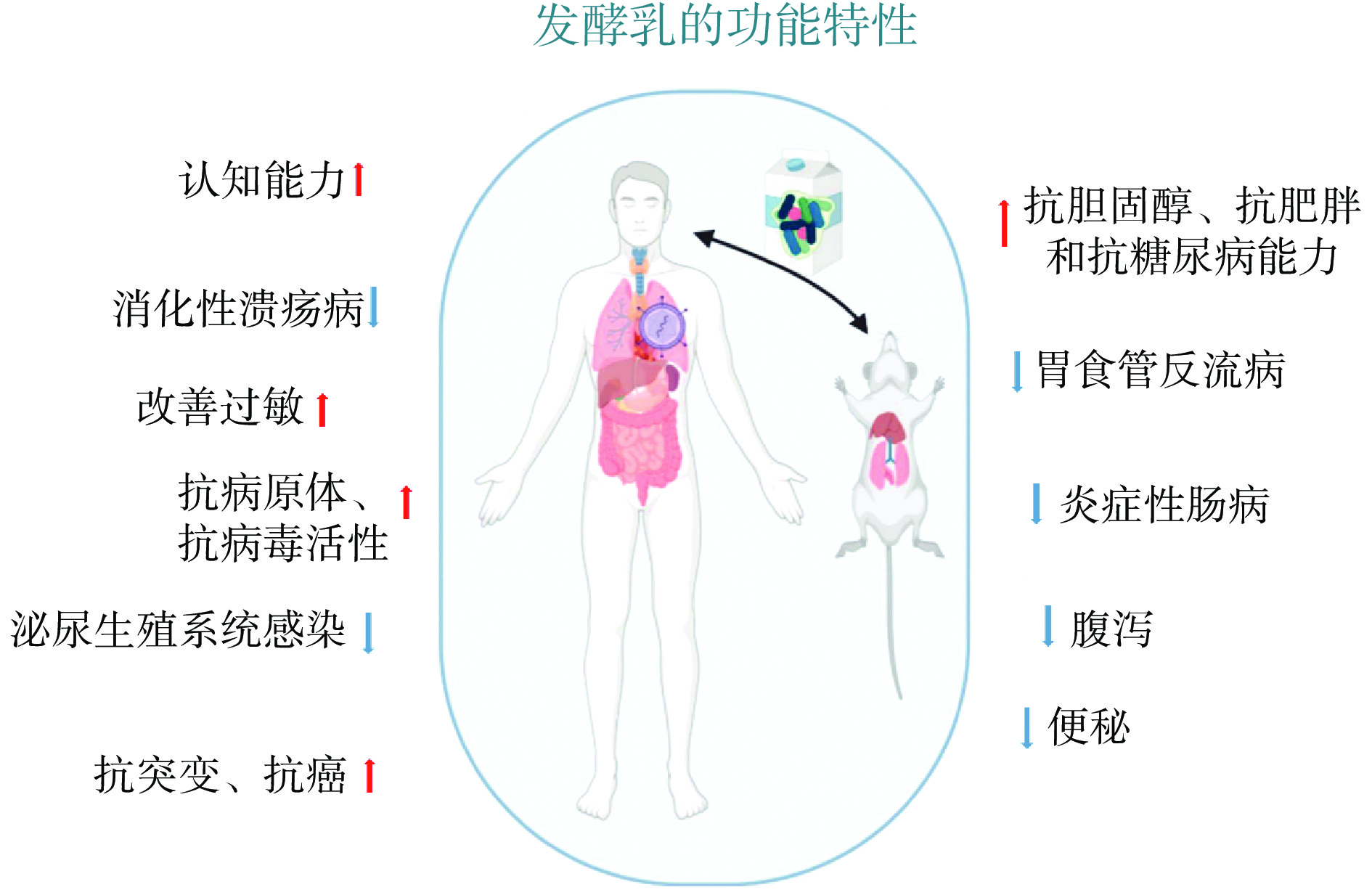
较多发酵乳作为一种功能性食品,越来越多的研究证实了发酵乳对人体的健康益处,如降低血清胆固醇[6]、改善肠道健康[7]、预防各种癌症[8]、促进免疫反应[9]和改善认知障碍[10]。图1列举了发酵乳的一些功能特性。由于消费者逐渐倾向于健康食品[11],并意识到发酵乳的益处,从而促进了对发酵乳的需求。同时,乳糖不耐症患病率的增加是发酵乳市场的另一个驱动力。虽然乳糖在发酵乳中不会完全降解,但引发乳糖不耐症的可能性会大大降低。此外,与未发酵的乳制品相比,发酵乳相对更容易消化[12]。随着人口的不断增长和高蛋白饮食的日益普及,消费者对发酵乳的认可度与接受度,也是全球发酵乳的消费量急剧增加的原因之一[13]。我国乳制品市场在未来五年仍会稳步发展,预测在2024年或将突破5500亿元。其中发酵乳占比35%,是乳制品市场的第二大品类。从发展形势上看,得益于常温发酵乳的出现,发酵乳品类的市场还在逐步增大,在2024年预计可以达到42%[14],发展潜力巨大。
市场上的许多发酵乳制品是用单一乳酸菌菌株或传统发酵剂(嗜热链球菌和保加利亚乳杆菌)生产的[16],如嗜酸乳杆菌酸乳、干酪乳杆菌酸乳、酸奶等。然而,这类发酵乳通常存在生产发酵时间长、口感不佳等缺点。而通过研究具有不同发酵特性的一些菌株,进行复配可中和或弥补单菌株发酵乳的一些缺陷,复配菌株共同发挥作用进而改善发酵乳品质。本文拟对近年来国内外复配发酵改善发酵乳品质及功能特性等方面的最新研究进展进行综述,以期为深入探究复配发酵乳在宿主健康调控功能中的作用机制、产品研发、实际生产应用提供依据。
1. 复配菌株发酵改善理化特性
1.1 pH与酸度
pH4.5通常被用作为制造发酵乳时的发酵终点。酸度是感知发酵乳风味的重要参数[17]。适当的酸度使产品具有独特的风味,并抑制腐败细菌和食源性病原体的生长。
经过复配后的菌株能够更快地利用糖酵解、蛋白水解和脂解等途径,将乳基中的碳水化合物、蛋白质和脂肪转化为乳酸、游离氨基酸和游离脂肪酸等物质,从而加快发酵乳的酸化速度[18]。Zhou等[19]将瑞士乳杆菌H9与商业发酵剂复配,结果表明,在相同接种量的情况下,添加瑞士乳杆菌H9可以显著加快发酵乳pH的下降速度,使发酵乳pH的下降速率从0.4(无H9添加)增加到0.53,因此通过复配具有高蛋白水解能力的菌株,可以达到增加pH下降速率的目的。Zhao等[20]将凝结芽孢杆菌13002作为新的潜在发酵剂掺入发酵乳中进行发酵,比较三组发酵乳包括a.嗜热链球菌:保加利亚乳杆菌=1:1(w/w)、b.嗜热链球菌:保加利亚乳杆菌:凝结芽孢杆菌13002=1:1:3(w/w/w)、c.凝结芽孢杆菌13002,发现复配组更快达到发酵终点,同时可滴定酸度的变化速率最大,可在2 h内从初始的14.67°T增长到61.64°T,而嗜热链球菌和保加利亚乳杆菌、凝结芽孢杆菌13002两组只能从14.67°T分别增长到44.74°T和37.15°T,与复配组有显著差异。可见凝结乳杆菌可以与嗜热链球菌和保加利亚乳杆菌协同作用,加速蛋白质的分解。
1.2 持水能力
持水能力是指发酵乳保留全部或部分自身水分的能力,持水能力低的发酵乳会导致乳清析出,从而导致发酵乳质量变差[21]。
复配菌株与乳中的蛋白质相互作用,以增强发酵乳的水合作用和持水能力,并且复配菌株发酵乳的凝胶网络相对致密,同时在复配菌株代谢过程中生成的水要多于消耗的水,所以发酵乳的持水力有所改善[22]。Yang等[23]制作山羊发酵乳除使用传统发酵剂外,还分别复配了四种菌株进行发酵,包括嗜酸乳杆菌、动物双歧杆菌、干酪乳杆菌和植物乳杆菌,山羊发酵乳的持水能力随着贮藏时间的延长而显著下降,复配发酵剂提高了山羊发酵乳贮藏28 d后的持水能力,贮藏结束时,含有复配发酵剂的山羊发酵乳持水能力在53%左右波动;值得注意的是,由于菌株特异性,嗜酸乳杆菌和干酪乳杆菌在山羊奶发酵过程中会产生更多的胞外多糖(exopoly saccharides,EPS),因此含有嗜酸乳杆菌和干酪乳杆菌的山羊发酵乳比含有动物双歧杆菌和植物乳杆菌的山羊发酵乳具有更高的持水能力。Zhang等[24]将发酵乳杆菌HY01与商业发酵剂MY105(包含嗜热链球菌和保加利亚乳杆菌)组合发酵牦牛奶,与只使用HY01或MY105发酵的进行对比发现,因为复配菌株制备的发酵乳提高了其凝胶强度并减小了孔径,从而锁定了更多的水进一步减少了脱水收缩,所以复配菌株制备的发酵乳持水力显著高于其它组;此外还观察到,其在贮藏过程中的黏度和凝胶强度优于单独添加HY01或商业发酵剂的发酵乳。
1.3 流变特性
发酵乳的流变特性主要是pH变化以及发酵剂对存在的大分子和颗粒的影响而决定[25]。乳酸菌在生长过程中产生乳酸,乳酸积累会导致乳pH降低,复配酵母和乳酸菌可以促进酪蛋白胶束的结合,导致发酵过程中乳的表观黏度增加。同时,还促进EPS的产生,EPS可以改善发酵乳的质地和感官特征。Huang等[26]采用含乳酸菌(嗜热链球菌:保加利亚乳杆菌=1:1)和马克思克鲁维酵母Y51-6的多发酵剂发酵系统,获得山羊奶发酵乳。进行流变学分析得出结论:发酵可以提高羊奶的表观黏度;在整个剪切速率范围内,乳酸菌和酵母共发酵的羊奶黏度高于单独乳酸菌发酵的羊奶黏度;在频率扫描测试中,由于马克思克鲁维酵母刺激了乳酸菌的生长,使其产生了更多的EPS,所以共培养发酵乳的弹性模量和黏性模量最高。Wang等[27]将从母乳中分离、筛选出的植物乳杆菌MWLp-12和发酵粘液乳杆菌MWLf-4复配到商业发酵剂中,用于发酵普通牛乳和A2牛乳;在两种不同的牛乳基质中,复配菌株发酵乳的弹性模量和黏性模量始终大于商业发酵剂的发酵乳,复配菌株发酵乳的表观黏度也分别高于商业发酵剂组,表观黏度反映了发酵乳的结构和稳定性[28],这意味着两种乳酸菌菌株的添加有助于发酵乳获得更优的流变特性。
2. 复配菌株发酵改善挥发性风味物质
发酵食品的味道和滋味可能会随着新物质的产生和发酵过程中风味成分的浓度而变化[29]。发酵乳中的挥发性风味化合物可以分为醛类、酮类、酸类、醇类、酯类和其它。表2介绍了目前发酵乳中常见的挥发性风味物质[30]。
类别 关键物质 风味形容 醛类 乙醛、丁醛、3-甲基丁醛、2-戊烯醛、己醛、2-辛烯醛、壬醛等 青草和水果香气 酮类 2,3-丁二酮、苯乙酮、2-庚酮、3-辛酮、2,3-戊二酮等 黄油、奶油香气 酸类 乙酸、己酸、辛酸、3-甲基丁酸、2-甲基丙酸 坚果香、奶酪味 醇类 2-乙基己醇、3-甲基丁醇、1-戊醇、乙醇等 酒花香气 酯类 丙酸乙酯、醋酸甲酯、甲酸乙烯酯 苹果和菠萝果香味 其它 甲硫醚、月桂烯 蔬菜香气、油脂香 在发酵过程中,挥发性化合物主要通过微生物转化形成,发酵乳制品的挥发性特征的差异可以直接归因于不同菌株的差异性[31]。在与传统发酵剂的共同发酵中,复配菌株不会影响发酵乳中关键的挥发性代谢物,但在经过规定的发酵时间和温度后会产生不同数量的代谢产物[32]。例如,添加乳杆菌属的菌株将有助于在发酵过程中增加挥发性有机酸和醇类物质的产量,并在冷藏过程中增加非挥发性有机酸和游离氨基酸的形成。Gu等[33]采用代谢组学分析评估了与传统发酵剂共发酵的副干酪乳杆菌IMC502对发酵乳代谢物的影响,采用顶空固相微萃取和气相色谱-质谱法测定了196种挥发性代谢物,副干酪乳杆菌IMC502不影响乙醛、双乙酰和乙酰脲的含量,但是复配菌株后可以促进丙酮(甜味、果味)和丁酸(黄油味)的形成。Papaioannou等[34]将双歧杆菌BB-12与市售发酵剂共培养用于牛奶发酵、将嗜酸乳杆菌LA-5与市售发酵剂共培养用于山羊奶发酵,检测乳制品的挥发性风味物质;乙醛形成的最重要途径是苏氨酸由苏氨酸醛缩酶催化分解成乙醛和甘氨酸,这种酶存在于嗜热链球菌和保加利亚乳杆菌中;不添加双歧杆菌BB-12的发酵乳,仅检测到乙醛;当复配双歧杆菌BB-12时,可以在发酵乳中检测到乙醛、戊醛、己醛和壬醛。另外,复配了嗜酸乳杆菌LA-5的山羊发酵乳与未复配相比,酮含量高出约2.6倍;这进一步验证了针对性地复配菌株是可以改善发酵乳风味的一种策略。
除了复配乳酸菌外,添加酵母菌株也是改善发酵乳风味的一种方法。Wang等[35]研究发现酵母代谢物可以中和瑞士乳杆菌SNA12发酵产生的酸。并且,共培养发酵乳的浓郁度和回味高于单一发酵乳。Korma等[36]研究了用酵母和五种乳酸菌混合共同发酵豆浆的情况,结果表明,在共同发酵过程中产生了大量的醇、酮、酸和酯,显著改善了发酵豆浆的风味质量,且共同培养的发酵乳的咸味、鲜味和涩味与单一发酵的相似。
3. 复配菌株发酵改善功能特性
3.1 降血脂
随着饮食结构的变化,人们的饮食逐渐高脂化,导致体重增加同时还会导致代谢和慢性疾病,对人类健康构成严重威胁。有研究表明,食用发酵乳可以促进肠道蠕动,并降低血脂、改善非酒精性脂肪肝、减少心脑血管疾病[37]。
菌株复配后使过氧化物酶体增殖物激活受体γ(peroxisome proliferator-activated receptor-γ,PPAR-γ)基因在机体肝脏和脂肪组织中的表达下降程度显著大于未复配菌株的发酵乳;在血清生化方面,可以更好地抑制甘油三酯(triglyceride,TG)、碱性磷酸酶(alkaline phosphatase,AKP)、谷氨酸草酰乙酸转氨酶(glutamic oxaloacetic transaminase,GOT)的升高以及高密度脂蛋白胆固醇(high-density lipoprotein cholesterol,HDL-c)的降低[38]。Li等[39]使用从牦牛酸奶中分离的植物乳杆菌HFY01复配商业保加利亚乳杆菌发酵豆浆,以探讨复配菌株对高脂小鼠的作用效果,结果表明复配LP-HFY01发酵豆浆可降低肥胖小鼠体脂率和肝指数;对于PPAR-γ基因,复配菌株发酵调节基因的表达更有效,与保加利亚乳杆菌发酵组呈现显著差异;用复配菌株发酵还显著抑制CCAAT/增强子结合蛋白-α(CCAAT/enhancer-binding protein-α,C/EBP-α)基因在肝脏和脂肪组织中的表达,其效果与左旋肉碱药物处理相似。Cho等[40]探讨从蒙古传统乳制品中分离出的发酵乳杆菌TS1和S2混合发酵乳对高脂饮食诱导肥胖大鼠的代谢作用,相比于单菌株发酵,复配菌株发酵更加显著地减少了腹部脂肪,改善了血清高密度脂蛋白胆固醇水平和脂联素分泌。虽然两种菌株组合的协同作用尚不清楚,但可以将其作为改善血液中胆固醇和体内脂肪积累的潜在发酵剂。
3.2 抗炎
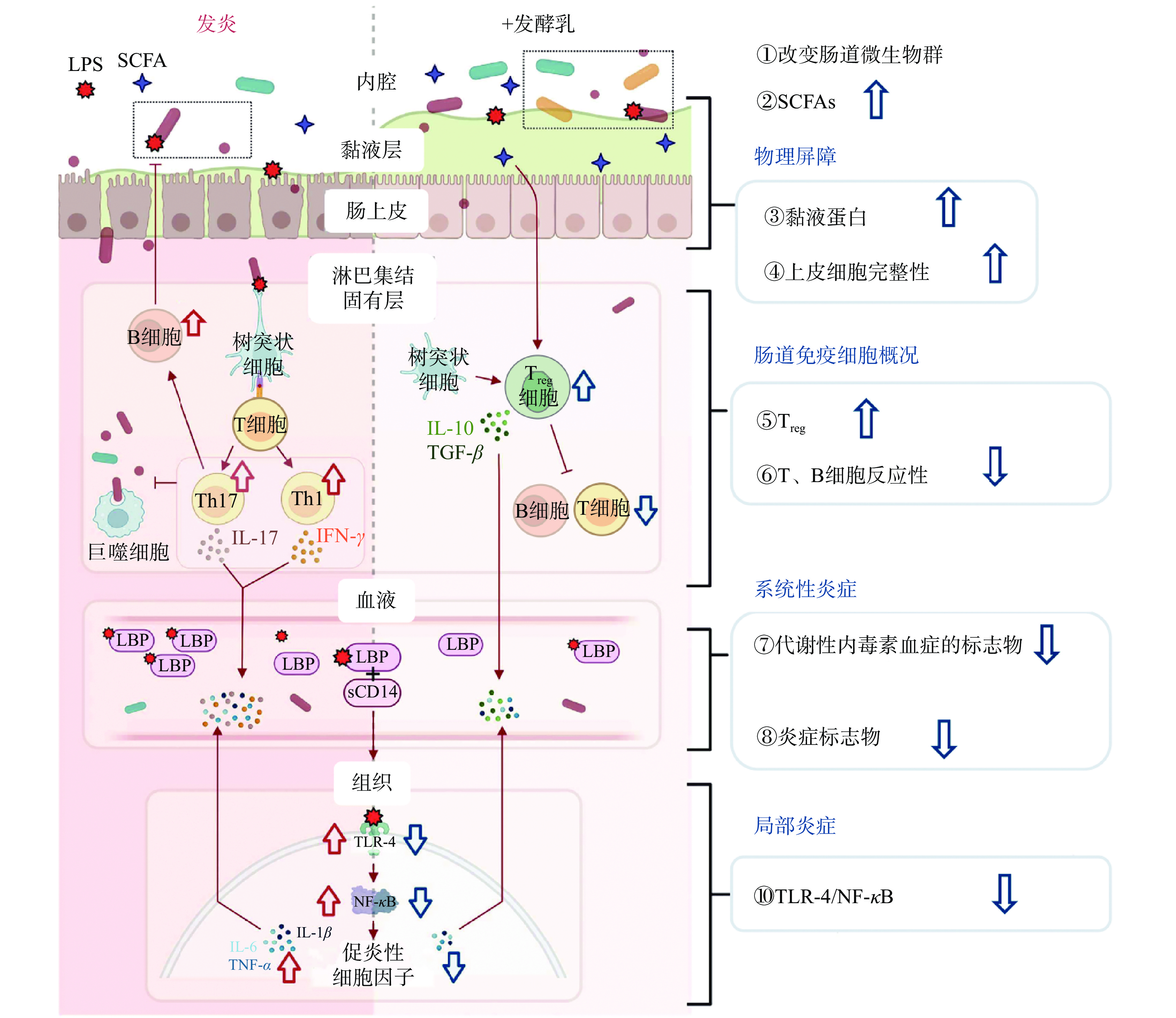
慢性、未解决的炎症是许多疾病风险的基础。有研究表明,增加发酵乳的摄入可以减少炎症的生物标志物并增加肠道微生物多样性[41]。图2表达了发酵乳的抗炎作用。
![]()
3.2.1 抗炎症性肠病
炎症性肠病(inflammatory bowel disease,IBD)是一种全身性疾病,主要包括溃疡性结肠炎和克罗恩病。它通常发生在免疫系统对常驻肠道微生物群产生过度的免疫反应时,引发一系列破坏肠壁的炎症事件[43]。IBD患者患神经退行性疾病的风险更高[44]。此外,IBD患者更容易患上胃肠道肿瘤[45]。研究表明发酵乳对炎症存在调节作用,摄入发酵乳可以将宿主肠道微生物群从紊乱恢复到正常状态[46]。
复配菌株发酵乳主要通过激活相应的信号通路,抑制促炎性细胞因子的表达,从而起到抗炎作用。Feng等[47]将商业发酵剂YF904与副干酪乳杆菌PC-01、植物乳杆菌Lp-6、瑞士乳杆菌H9、乳双歧乳杆菌M8复配制成巴氏杀菌发酵乳,胃内给予复配菌株发酵乳使大鼠体重恢复到与正常组相似的水平,也显著高于葡聚糖硫酸钠(dextran sulfate sodium,DSS)诱导组;食用只有YF904的发酵乳仅仅增加了DSS处理的大鼠的体重;给予复配菌株发酵乳可以显著降低结肠组织学评分,但在补充YF904发酵乳给药的情况下,这种保护效果较差;对于血清中促炎因子的水平,发现食用复配菌株发酵乳降低IL-6和TNF-α水平显著优于YF904发酵乳;但对于IL-1β、IL-10水平无显著差异,原因还有待深入探讨;与YF904发酵乳相比,复配菌株发酵乳的KEGG代谢途径(主要是与核苷酸和氨基酸生物合成相关的途径)显著富集,表明使用复配菌株发酵乳可以有效调节肠道微生物群及其潜在功能。Lao等[48]的研究也证明了将嗜热链球菌、保加利亚乳杆菌、植物乳杆菌以1:1:2复配菌株得到的发酵乳比单菌株发酵乳缓解炎症效果更加明显;乳酸菌使牛奶脂肪裂解产生的脂肪酸可以通过抑制炎症因子IL-6和TNF-α的表达,并通过促进抗炎因子IL-10的分泌阻断JNK-MAPK信号通路的磷酸化来缓解DSS诱导的结肠炎。
3.2.2 缓解过敏
牛奶是食品工业中广泛使用的加工原料[49]。然而,牛奶中含有抗原蛋白(酪蛋白、乳清蛋白),可诱发过敏反应,包括湿疹、肺部浸润、鼻炎和腹泻,被联合国粮食及农业组织和世界卫生组织认为是8种主要过敏原之一。在牛奶发酵过程中,蛋白质结构发生变化,降低了其致敏性。
由于不同的乳酸菌具有不同的蛋白水解系,所以菌株降低乳蛋白抗原性的能力不同。研究证实,Th2细胞和B细胞的发育和活化是产生IgE的主要因素[50]。另据报道,Th2细胞产生的细胞因子IL-4在IgE介导的牛乳过敏中起着至关重要的作用,它能诱导IgE类的转换及其产生[51]。此外,Th1细胞产生的IFN-γ可抑制IL-4的产生,从而减少IgE的产生,缓解过敏反应[52]。Zhao等[53]使用BALB/c小鼠评估瑞士乳杆菌KLDS 1.8701和植物乳杆菌KLDS 1.0386复配发酵对过敏小鼠的治疗效果,结果显示,与未发酵和单菌株发酵相比,复配组的Th2相关细胞因子IL-4、IL-5水平显著下降,Th1相关细胞因子IFN-γ水平显著升高。Wróblewska等[54]也发现相比于用商业发酵剂或其它菌株复配制成的发酵乳,用嗜热链球菌2K和保加利亚乳杆菌BK与植物乳杆菌W42和乳酸芽孢杆菌Bi30菌株一起发酵后喂养的过敏小鼠,其脾脏分泌细胞因子IL-10和TGF-β能力显著增强;Th2细胞的活性降低,脾细胞分泌的IL-4减少,血清中的IgE和抗IgG1抗体也减少;反过来,血清和粪便中的总IgA分泌量以及脾细胞中的总IgA和IgG也有增加;这种机制伴随着免疫反应恢复至Th2/Th1平衡状态。因此,共同发酵可以更好地调节Th1/Th2平衡并降低过敏标志物水平,从而减少乳制品的致敏。筛选具有协同作用的菌株复配成发酵剂,应用于抗过敏功能性乳制品中,可以成为未来食品的研究方向。
3.3 抑菌活性
致病菌导致的发病率和死亡率在逐年上升,并对全世界造成直接的经济损失。抗生素治疗被认为是针对致病菌的最常见治疗方法。然而,抗生素由于滥用而造成了致病菌耐药性等问题,降低了其效率。因此,开发有前途的抗生素替代品对于预防和治疗致病菌势在必行。Abdelghani等[55]采用盘扩散琼脂技术检测不同复配菌株组合发酵豆浆对致病菌菌株的拮抗作用,结果显示“植物乳杆菌ATCC14917+嗜热乳杆菌DSM20259+长双歧杆菌B41409”组合对大肠杆菌O157:H7、志贺链球菌As2、伤寒沙门氏菌As3、金黄色葡萄球菌As4、单核细胞增生李斯特菌、蜡样芽孢杆菌Dsmz345共培养的抑制直径最大,分别为22、17、16、15、13、12 mm;而即便是最有效的单菌株(植物乳杆菌)发酵豆乳对这些病原菌的抑制直径都要小于复配组。Hyun等[56]将单核细胞增生李斯特菌与不同菌株(商业发酵剂、商业发酵剂与不同乳酸片球菌复配)发酵的酸奶混合培养,测定抑菌能力,发现在初始和储藏1 d后,市售发酵乳中李斯特菌的菌落数分别为5.28和6.45 log CFU/mL,而复配乳酸片球菌的发酵乳中数量显著降低,尤其是与乳酸片球菌JM01复配,此发酵乳中李斯特菌的数量分别为3.57和5.28 log CFU/mL。复配菌株发酵乳比普通发酵乳具有更高的抑菌活性,可以考虑将其作为抗生素的辅助产品。
3.4 抗氧化
发酵乳的抗氧化活性在很大程度上取决于菌株产生的生物活性化合物,因为这些化合物作为电子供体,与自由基反应,将其转化为更稳定的产物并终止氧化反应[57],所以可以通过复配对应的菌株进一步增强发酵乳的抗氧化活性。发酵乳的抗氧化活性可以用DPPH·、ABTS+·、·OH的自由基清除活性(radical scavenging activity,RSA)和铁还原力(ferric iron-reducing power,FRP)来衡量。Zhao等[58]制备了四种发酵乳(植物乳杆菌MC5与商业发酵剂的比例分别为0:1、1:1、2:1和1:2),研究发酵乳在储存期间的抗氧化活性,在储存期间,对照组的DPPH、ABTS+和OH的RSA缓慢增加,而FRP保持不变;1:1、2:1和1:2组的DPPH、ABTS+和OH的RSA在储存期间以较高的速度增加,特别是1:1和2:1组;与对照组相比,1:1组的DPPH、ABTS+和OH的RSA在储存期间分别增加了13.18%、14.14%和27.81%,而2:1组的DPPH、ABTS+和OH的RSA分别增加了18.87%、23.87%和25.77%,这些发现表明,复配MC5发酵后的代谢产物可以提供电子或氢来清除自由基,起到抗氧化的效果。
除了测定DPPH、ABTS+、OH的RSA来评估乳制品的抗氧化活性,多酚作为抗氧化剂被广泛熟知,复配菌株可以提高发酵乳中多酚含量,进而增强其抗氧化活性。Shori等[59]将干酪乳杆菌ATCC393、植物乳杆菌ATCC14917、鼠李糖乳杆菌ATCC53103这三株菌种分别与嗜热链球菌St1342、保加利亚乳杆菌ATCC7830组合发酵腰果奶,测定不同处理的发酵乳样品在储存21 d内的总酚与类黄酮含量,结果表明,在整个储存期间,使用组合菌株发酵乳的总酚与黄酮含量都要显著高于对照组;尤其是植物乳杆菌组合的发酵乳总酚含量最高为83 mg/mL,鼠李糖乳杆菌组合的发酵乳黄酮含量最高为56.3 mg/g,在腰果奶发酵过程中额外添加菌株可减少植物化学物质的降解,从而增加发酵乳中抗氧化类化合物的含量,从而带来潜在的健康益处[60]。
3.5 缓解乳糖不耐症
传统上,发酵乳制作依赖于两种菌株:嗜热链球菌和保加利亚乳杆菌,但这两种菌株通常仅代谢乳糖中的葡萄糖部分并将未被代谢的半乳糖释放到胞外,导致发酵乳中残留较高含量的半乳糖和未发酵乳糖,引发乳糖不耐症。复配特定菌株可以分泌酶消化掉残留的乳糖和半乳糖,简而言之,就是菌株可以分泌由galM、galK、galT和galE编码的酶将引起过敏的半乳糖转化为葡萄糖[61]。Zhang等[62]研究发现植物乳杆菌可以利用Leloir途径有效地代谢乳糖和半乳糖,因此将植物乳杆菌WCFS1与嗜热链球菌及保加利亚乳杆菌共同培养制作发酵乳,结果显示,乳糖被完全消耗,半乳糖被有效代谢;与传统的发酵相比,添加植物乳杆菌WCFS1使得总糖含量降低;该研究发掘了植物乳杆菌与传统发酵剂共同培养制造低糖发酵乳的潜力。Masoumi等[63]评估了嗜酸乳杆菌和双歧杆菌共同作为发酵剂制备乳制品的健康效果,结果表明以这两种菌株复配得到的发酵乳可以促进乳糖酶分泌到消化系统中以提高乳糖的消化率,进而缓解乳糖不耐症。
3.6 其他
除了前文详细阐述的复配菌株发酵乳可以增强降血脂、抗炎(抗炎症性肠病、缓解过敏)、抑菌活性、抗氧化、缓解乳糖不耐症等功能特性,复配菌株发酵乳还对增长认知能力、调节情绪、缓解便秘和腹泻等有一定的改善效果。富含GABA的发酵乳具有作为辅助治疗轻度抑郁症的新膳食疗法的潜力。Wu等[64]筛选了高产GABA的菌株短乳杆菌15953,以保加利亚乳杆菌:嗜热链球菌:植物乳杆菌15953:短乳杆菌J1为1:1:3:1的比例制备发酵乳,此组合得到的发酵乳GABA含量最高为241.3 μg/mL。复配组中的小鼠社交能力得到显著改善,并且在强迫游泳和悬尾实验中面对恐惧的反应也要优于其他实验组。研究表明富硒可以缓解小鼠认知功能障碍,上调海马神经发生和p-CREB信号通路被认为是预防和治疗脑衰老的有效策略[65]。Wu等[66]在发酵剂选择方面,除了传统的发酵剂(保加利亚乳杆菌和嗜热链球菌)外,同时采用植物乳杆菌PDD-1和发酵乳杆菌PDD-4加入消毒纯乳中,比例为2:3:1,发酵后测得总硒含量为192.50 μg/L,高于对照组;同时在动物实验中观察到,食用复配菌株发酵乳可显著增加D-半乳糖模型小鼠海马神经发生,并上调ERK-CREB-BDNF信号通路中的蛋白表达。
4. 总结与展望
本文介绍了发酵乳种类、常见发酵菌种以及复配菌株改善发酵乳的品质及功能特性。品质方面,复配菌株制成的发酵乳可以提高pH的下降速率、提升酸度;还可以改变发酵乳中的风味物质,改善感官特性。复配不同的菌株促进代谢产物如胞外多糖的产生,进而提高发酵乳的持水能力,改变其流变学特性。对于功能特性方面,复配菌株发酵后的乳制品可以降低体脂率和肝指数,调节相关基因表达起到降脂的作用。通过调节促炎、抗炎细胞因子以及肠道菌群可以改善炎症性肠道疾病和过敏症状。不同特征的菌株复配协同可以产生更高水平的生物活性物质,从而增强发酵乳的抑菌及抗氧化活性等。但目前国内外的研究多数聚焦于菌株复配后的表层现象和代谢物质变化等方面,缺少一些关于菌株复配在机制方面的研究,因此未来的研究方向可以基于此对机制作用进行更深层次的研究和讨论。
随着社会的多元化以及大众对食品的高要求,发酵乳产品需要不断推陈出新,满足不同消费者的差异化要求,在复配方面具有创新可能性:a.不仅对菌株进行复配,还可以与功能性或天然性成分复配,例如后生元(多糖、短链脂肪酸)、益生元(低聚糖、菊粉、蛋白质水解物)、维生素等,从而达到免疫调节、缓解眼疲劳、减压助眠和预防、缓解部分疾病的作用;b.在菌株复配的基础上,同时对菌株辅以激光、微胶囊处理或对乳进行超声波、超滤处理等以延长发酵乳的保质期、提高发酵乳的稳定性等;c.针对不同的乳基质的特点,挖掘相应的菌株进行复配,优化菌株作用以弥补基质的不足或增强基质的功能特性,从而研发出营养价值丰富、生物活性突出和风味更好的发酵乳产品。
-
![]()
表 1 不同类型发酵乳的特点、优势及不足
Table 1 Characteristics, advantages and disadvantages of different types of fermented milk
分类标准 发酵乳种类 特点 优势 不足 保藏温度 常温发酵乳[3] 生产过程中进行二次巴氏杀菌 可以在4~25 ℃保存5~6个月,便于长途运输;携带方便;保质期长 营养创新被限制,目前只能向口味创新、饮品和甜品化方向发展[4] 低温发酵乳[5] 需要在2~6 ℃储存,生产过程中只进行一次巴氏杀菌 存有活性乳酸菌,可以促进人体肠道消化、调节肠道菌群健康 保存时间有限;储存要求高;
销售成本高口感 原味发酵乳 保留传统发酵乳最醇正的口感风味 无其它物质添加,较风味发酵乳更加安全 口味单一,不能满足消费者对于商品多元化的需求 风味发酵乳 除乳或乳粉外,同时可以添加其它原料,选择性添加食品添加剂、营养强化剂、果蔬、谷物等 口味丰富,满足不同人群的感官要求 产品质量参差不一;额外添加成分
较多 下载: 导出CSV
下载: 导出CSV
-
[1] COREY E J, GHOSH A K. Mn(III)-promoted annulation of enol ethers and esters to fused or 2-cyclopentenones[J]. Tetrahedron Letters,1987,28(2):175−178. doi: 10.1016/S0040-4039(00)95679-X
[2] 中华人民共和国卫生部. GB 19302-2010 食品安全国家标准 发酵乳[S]. 北京:中国标准出版社, 2010. [Ministry of Health of the People's Republic of China. GB 19302-2010 National food safety standard Fermented milk[S]. Beijing:Standards Press of China, 2010.] Ministry of Health of the People's Republic of China. GB 19302-2010 National food safety standard Fermented milk[S]. Beijing: Standards Press of China, 2010.
[3] CHUAH L O, MAO Y. Stability assessment and improvement of a Lactobacillus plantarum mutant with low post-fermentation acidification characteristics[J]. Journal of Dairy Science,2020,103(9):7898−7907. doi: 10.3168/jds.2020-18285
[4] 王弘烨. 常温酸奶来势汹汹, 低温酸奶能否重绽芳华[J]. 乳品与人类,2022,123(2):20−26. [WANG H Y. Low-temperature yoghurts are coming on strong, can low-temperature yoghurts make a comeback[J]. Dairy and Humankind,2022,123(2):20−26.] WANG H Y. Low-temperature yoghurts are coming on strong, can low-temperature yoghurts make a comeback[J]. Dairy and Humankind, 2022, 123(2): 20−26.
[5] 嵇海涛. 低温酸乳产品霉菌和酵母的控制[J]. 食品安全导刊,2022,344(15):130−133. [JI H T. Control of mold and yeast in low temperature yoghurt products[J]. China Food Safety Magazine,2022,344(15):130−133.] JI H T. Control of mold and yeast in low temperature yoghurt products[J]. China Food Safety Magazine, 2022, 344(15): 130−133.
[6] AYYASH M, AL-DHAHERI A S, AL MAHADIN S, et al. In vitro investigation of anticancer, antihypertensive, antidiabetic, and antioxidant activities of camel milk fermented with camel milk probiotic:A comparative study with fermented bovine milk[J]. Journal of Dairy Science,2018,101(2):900−911. doi: 10.3168/jds.2017-13400
[7] HOU Q, LI C, LIU Y, et al. Koumiss consumption modulates gut microbiota, increases plasma high density cholesterol, decreases immunoglobulin G and albumin[J]. Journal of Functional Foods,2019,52:469−478. doi: 10.1016/j.jff.2018.11.023
[8] 于洁, 张和平. 益生菌发酵乳的研究及产业化进展[J]. 中国食品学报,2020,20(10):1−7. [YU J, ZHANG H P. Research and industrialization progress on probiotics fermented dairy products[J]. Journal of Chinese Institute of Food Science and Technology,2020,20(10):1−7.] YU J, ZHANG H P. Research and industrialization progress on probiotics fermented dairy products[J]. Journal of Chinese Institute of Food Science and Technology, 2020, 20(10): 1−7.
[9] KECHAGIA M, BASOULIS D, KONSTANTOPOULOU S, et al. Health benefits of probiotics:A review[J]. ISRN Nutrition,2013,2013:481651.
[10] LI C, KWOK L Y, MI Z, et al. Characterization of the angiotensin-converting enzyme inhibitory activity of fermented milks produced with Lactobacillus casei[J]. Journal of Dairy Science,2017,100(12):9495−9507. doi: 10.3168/jds.2017-12970
[11] BOUKID F, LAMRI M, DAR B N, et al. Vegan alternatives to processed cheese and yogurt launched in the buropean market during 2020:A nutritional challenge?[J]. Foods,2021,10(11):2782. doi: 10.3390/foods10112782
[12] SAVAIANO D A, HUTKINS R W. Yogurt, cultured fermented milk, and health:A systematic review[J]. Nutrition Reviews,2021,79(5):599−614. doi: 10.1093/nutrit/nuaa013
[13] BARROS C P, GUIMARÃES J T, ESMERINO E A, et al. Paraprobiotics and postbiotics:Concepts and potential applications in dairy products[J]. Current Opinion in Food Science,2020,32:1−8.
[14] WANG F, WANG H, CHO J H. Consumer preference for yogurtpackaging design using conjoint analysis[J]. Sustainability,2022,14(6):3463. doi: 10.3390/su14063463
[15] SAKANDAR H A, ZHANG H. Trends in probiotic(s)-fermented milks and their in vivo functionality:A review[J]. Trends in Food Science & Technology,2021,110:55−65.
[16] MEYBODI N M, MORTAZAVIAN A M, ARAB M, et al. Probiotic viability in yoghurt:A review of influential factors[J]. International Dairy Journal,2020,109:104793. doi: 10.1016/j.idairyj.2020.104793
[17] LIU W, PU X, SUN J, et al. Effect of Lactobacillus plantarum on functional characteristics and flavor profile of fermented walnut milk[J]. LWT-Food Science and Technology,2022,160:113254. doi: 10.1016/j.lwt.2022.113254
[18] WANG X, KRISTO E, LAPOINTE G. Adding apple pomace as a functional ingredient in stirred-type yogurt and yogurt drinks[J]. Food Hydrocolloids,2020,100:105453. doi: 10.1016/j.foodhyd.2019.105453
[19] ZHOU T, HUO R, KWOK L Y, et al. Effects of applying Lactobacillus helveticus H9 as adjunct starter culture in yogurt fermentation and storage[J]. Journal of Dairy Science,2019,102(1):223−235. doi: 10.3168/jds.2018-14602
[20] ZHAO S, ZHOU Q Y, HUANG Y Y, et al. Comparative analysis of physicochemical, rheological, sensory and flavour properties of yoghurts using a new probiotic Bacillus coagulans 13002 with traditional yoghurt starter[J]. International Journal of Food Science & Technology,2020,56(4):1712−1723.
[21] GE X, TANG N, HUANG Y, et al. Fermentative and physicochemical properties of fermented milk supplemented with sea buckthorn (Hippophae eleagnaceae L.)[J]. LWT-Food Science and Technology,2022,153:112484. doi: 10.1016/j.lwt.2021.112484
[22] 张睿, 刘继超, 药璐, 等. 益生菌对发酵乳理化性质影响的研究[J]. 中国食品添加剂,2013(6):66−70. [ZHANG R, LIU J C, YAO L, et al. Study of probiotics on the impact of physical and chemical properties of fermented milk[J]. China Food Additives,2013(6):66−70.] ZHANG R, LIU J C, YAO L, et al. Study of probiotics on the impact of physical and chemical properties of fermented milk[J]. China Food Additives, 2013(6): 66−70.
[23] YANG Y, ZHANG R, ZHANG F, et al. Storage stability of texture, organoleptic, and biological properties of goat milk yogurt fermented with probiotic bacteria[J]. Frontiers in Nutrition,2022,9:1093654.
[24] ZHANG X, YANG J, ZHANG C, et al. Effects of Lactobacillus fermentum HY01 on the quality characteristics and storage stability of yak yogurt[J]. Journal of Dairy Science,2022,105(3):2025−2037. doi: 10.3168/jds.2021-20861
[25] BERNAT N, CHÁFER M, CHIRALT A, et al. Hazelnut milk fermentation using probiotic Lactobacillus rhamnosus GG and inulin[J]. International Journal of Food Science & Technology,2014,49(12):2553−2562.
[26] HUANG Z, HUANG L, XING G, et al. Effect of co-fermentation with lactic acid bacteria and K. marxianus on physicochemical and sensory properties of goat milk[J]. Foods,2020,9(3):299. doi: 10.3390/foods9030299
[27] WANG Y, FENG K, JIN J, et al. Comparison on properties between normal and A2 bovine milk fermented using commercial bacteria mixed with/without two probiotics from human milk[J]. International Journal of Biological Macromolecules,2022,216:105−113. doi: 10.1016/j.ijbiomac.2022.06.200
[28] AYALA-HERNANDEZ I, GOFF H D, CORREDIG M. Interactions between milk proteins and exopolysaccharides produced by Lactococcus lactis observed by scanning electron microscopy[J]. Journal of Dairy Science,2008,91(7):2583−2590. doi: 10.3168/jds.2007-0876
[29] LI H, GAO J, CHEN W, et al. Lactic acid bacteria isolated from Kazakh traditional fermented milk products affect the fermentation characteristics and sensory qualities of yogurt[J]. Food Science & Nutrition,2022,10(5):1451−1460.
[30] 牟泓羽, 陈橙, 匡崇航, 等. 高蛋白搅拌酸奶的研制及风味成分的分析[J]. 中国酿造,2023,42(1):160−167. [MO H Y, CHEN C, KUANG C H, et al. Development and flavor composition analysis of high-protein stir yogurt[J]. China Brewing,2023,42(1):160−167.] MO H Y, CHEN C, KUANG C H, et al. Development and flavor composition analysis of high-protein stir yogurt[J]. China Brewing, 2023, 42(1): 160−167.
[31] PAN D D, WU Z, PENG T, et al. Volatile organic compounds profile during milk fermentation by Lactobacillus pentosus and correlations between volatiles flavor and carbohydrate metabolism[J]. Journal of Dairy Science,2014,97(2):624−631. doi: 10.3168/jds.2013-7131
[32] CHEN C, ZHAO S, HAO G, et al. Role of lactic acid bacteria on the yogurt flavour:A review[J]. International Journal of Food Properties,2017,20(sup1):S316−S330. doi: 10.1080/10942912.2017.1295988
[33] GU Y, LI X, XIAO R, et al. Impact of Lactobacillus paracasei IMC502 in coculture with traditional starters on volatile and non-volatile metabolite profiles in yogurt[J]. Process Biochemistry,2020,99:61−69. doi: 10.1016/j.procbio.2020.07.003
[34] PAPAIOANNOU G, KOSMA I, BADEKA A V, et al. Profile of volatile compounds in dessert yogurts prepared from cow and goat milk, using different starter cultures and probiotics[J]. Foods,2021,10(12):3153. doi: 10.3390/foods10123153
[35] WANG X, LI W, MAHSA G C, et al. Co-cultivation effects of Lactobacillus helveticus SNA12 and Kluveromyces marxiensis GY1 on the probiotic properties, flavor, and digestion in fermented milk[J]. Food Research International,2023,169:112843. doi: 10.1016/j.foodres.2023.112843
[36] KORMA S A, LI L, GHAMRY M, et al. Effect of co-fermentation system with isolated new yeasts on soymilk:Microbiological, physicochemical, rheological, aromatic, and sensory characterizations[J]. Brazilian Journal of Microbiology,2022,53(3):1549−1564. doi: 10.1007/s42770-022-00773-7
[37] YADAV R, KHAN S H, MADA S B, et al. Consumption of probiotic Lactobacillus fermentum MTCC:5898-Fermented milk attenuates dyslipidemia, oxidative stress, and inflammation in male rats fed on cholesterol-enriched diet[J]. Probiotics and Antimicrobial Proteins,2019,11(2):509−518. doi: 10.1007/s12602-018-9429-4
[38] WENG B B C, YUAN H D, CHEN L G, et al. Soy yoghurts produced with efficient GABA (γ-aminobutyric acid)-producing Lactiplantibacillus plantarum ameliorate hyperglycaemia and re-establish gut microbiota in streptozotocin (STZ)-induced diabetic mice[J]. Food & Function,2023,14(3):1699−1709.
[39] LI C, LIU H, YANG J, et al. Effect of soybean milk fermented with Lactobacillus plantarum HFY01 isolated from yak yogurt on weight loss and lipid reduction in mice with obesity induced by a high-fat diet[J]. RSC Advances,2020,10(56):34276−34289. doi: 10.1039/D0RA06977A
[40] CHO W Y, HONG G E, LEE H J, et al. Effect of yogurt fermented by Lactobacillus fermentum TSI and L. fermentum S2 derived from a mongolian traditional dairy product on rats with high-fat-diet-induced obesity[J]. Foods,2020,9(5):594. doi: 10.3390/foods9050594
[41] WASTYK H C, FRAGIADAKIS G K, PERELMAN D, et al. Gut-microbiota-targeted diets modulate human immune status[J]. Cell,2021,184(16):4137−4153. doi: 10.1016/j.cell.2021.06.019
[42] HASEGAWA Y, BOLLING B W. Yogurt consumption for improving immune health[J]. Current Opinion in Food Science,2023,51:101017. doi: 10.1016/j.cofs.2023.101017
[43] ANANTHAKRISHNAN A N, KAPLAN G G, NG S C. Changing global epidemiology of inflammatory bowel diseases:Sustaining health care delivery into the 21st century[J]. Clinical Gastroenterology and Hepatology,2020,18(6):1252−1260. doi: 10.1016/j.cgh.2020.01.028
[44] KIM G H, LEE Y C, KIM T J, et al. Risk of neurodegenerative diseases in patients with inflammatory bowel disease:A nationwide population-based cohort study[J]. Journal of Crohn's and Colitis,2022,16(3):436−443. doi: 10.1093/ecco-jcc/jjab162
[45] LIU Y J, TANG B, WANG F C, et al. Parthenolide ameliorates colon inflammation through regulating Treg/Th17 balance in a gut microbiota-dependent manner[J]. Theranostics,2020,10(12):5225−5241. doi: 10.7150/thno.43716
[46] CHEN Y, JIN Y, STANTON C, et al. Alleviation effects of Bifidobacterium breve on DSS-induced colitis depends on intestinal tract barrier maintenance and gut microbiota modulation[J]. European Journal of Nutrition,2021,60(1):369−387. doi: 10.1007/s00394-020-02252-x
[47] FENG C, ZHANG W, ZHANG T, et al. Oral administration of pasteurized probiotic fermented milk alleviates dextran sulfate sodium-induced inflammatory bowel disease in rats[J]. Journal of Functional Foods,2022,94:105140. doi: 10.1016/j.jff.2022.105140
[48] LAO L, YANG G, ZHANG A, et al. Anti-inflammation and gut microbiota regulation properties of fatty acids derived from fermented milk in mice with dextran sulfate sodium-induced colitis[J]. Journal of Dairy Science,2022,105(10):7865−7877. doi: 10.3168/jds.2022-21877
[49] VILLA C, COSTA J, OLIVEIRA M B P P, et al. Bovine milk allergens:A comprehensive review[J]. Comprehensive Reviews in Food Science and Food Safety,2018,17(1):137−164. doi: 10.1111/1541-4337.12318
[50] FALLON P G, BALLANTYNE S J, MANGAN N E, et al. Identification of an interleukin (IL)-25-dependent cell population that provides IL-4, IL-5, and IL-13 at the onset of helminth expulsion[J]. Journal of Experimental Medicine,2006,203(4):1105−1116. doi: 10.1084/jem.20051615
[51] DOURADO L P, SALDANHA J C, GARGIULO D L, et al. Role of IL-4 in aversion induced by food allergy in mice[J]. Cell Immunol,2010,262(1):62−68. doi: 10.1016/j.cellimm.2009.12.010
[52] STOTT B, LAVENDER P, LEHMANN S, et al. Human IL-31 is induced by IL-4 and promotes TH2-driven inflammation[J]. Journal of Allergy and Clinical Immunology,2013,132(2):446−454. doi: 10.1016/j.jaci.2013.03.050
[53] ZHAO L, SHI F, XIE Q, et al. Co-fermented cow milk protein by Lactobacillus helveticus KLDS 1.8701 and Lactobacillus plantarum KLDS 1.0386 attenuates its allergic immune response in Balb/c mice[J]. Journal of Dairy Science,2022,105(9):7190−7202. doi: 10.3168/jds.2022-21844
[54] WROBLEWSKA B, KALISZEWSKA-SUCHODOLA A, FUC E, et al. Effect of low-immunogenic yogurt drinks and probiotic bacteria on immunoreactivity of cow's milk proteins and tolerance induction-in vitro and in vivo studies[J]. Nutrients,2020,12(11):3390. doi: 10.3390/nu12113390
[55] ABDELGHANI D Y, GAD A I, ORABI M M, et al. Bioactivity of organic fermented soymilk as next-generation prebiotic/probiotics mixture[J]. Fermentation,2022,8(10):513. doi: 10.3390/fermentation8100513
[56] HYUN I K, KIM M Y, KIM S, et al. Functional properties of yogurt fermented by bacteriocin-producing Pediococcus acidilactici[J]. Journal of Dairy Science and Biotechnology,2020,38(3):154−160. doi: 10.22424/jdsb.2020.38.3.154
[57] AHIRE J J, MOKASHE N U, PATIL H J, et al. Antioxidative potential of folate producing probiotic Lactobacillus helveticus CD6[J]. Journal of Food Science and Technology,2011,50(1):26−34.
[58] ZHAO X, LIANG Q. EPS-Producing Lactobacillus plantarum MC5 as a compound starter improves rheology, texture, and antioxidant activity of yogurt during storage[J]. Foods,2022,11(11):1160.
[59] SHORI A B, ALJOHANI G S, AL-ZAHRANI A J, et al. Viability of probiotics and antioxidant activity of cashew milk-based yogurt fermented with selected strains of probiotic Lactobacillus spp[J]. LWT-Food Science and Technology,2022,153:112482. doi: 10.1016/j.lwt.2021.112482
[60] TIWARI U, CUMMINS E. Factors influencing levels of phytochemicals in selected fruit and vegetables during pre- and post-harvest food processing operations[J]. Food Research International,2013,50(2):497−506. doi: 10.1016/j.foodres.2011.09.007
[61] CAO J, YU L, ZHAO J, et al. Genomic analysis of B. coagulans ATCC 7050T reveals its adaption to fermented milk as an adjunct starter culture for yogurt[J]. LWT-Food Science and Technology,2022,154:112721. doi: 10.1016/j.lwt.2021.112721
[62] ZHANG S S, XU Z S, QIN L H, et al. Low-sugar yogurt making by the co-cultivation of Lactobacillus plantarum WCFS1 with yogurt starter cultures[J]. Journal of Dairy Science,2020,103(4):3045−3054. doi: 10.3168/jds.2019-17347
[63] MASOUMI S J, MEHRABANI D, SABERIFIROOZI M, et al. The effect of yogurt fortified with Lactobacillus acidophilus and Bifidobacterium sp. probiotic in patients with lactose intolerance[J]. Food Science & Nutrition,2021,9(3):1704−1711.
[64] WU Z, WANG P, PAN D, et al. Effect of adzuki bean sprout fermented milk enriched in gamma-aminobutyric acid on mild depression in a mouse model[J]. Journal of Dairy Science,2021,104(1):78−91. doi: 10.3168/jds.2020-19154
[65] KOCOT A M, WRÓBLEWSKA B. Fermented products and bioactive food compounds as a tool to activate autophagy and promote the maintenance of the intestinal barrier function[J]. Trends in Food Science & Technology,2021,118:905−919.
[66] WU Z, CHEN T, PAN D, et al. Resveratrol and organic selenium-rich fermented milk reduces D-galactose-induced cognitive dysfunction in mice[J]. Food & Function,2021,12(3):1318−1326.
-
期刊类型引用(1)
1. 刘志忠,刘立伟,刘志刚,陈海丽,崔红,杨波,张佳琪,乌日更,姚凯. 唾液链球菌嗜热亚种与动物双歧杆菌乳亚种复合发酵对发酵乳品质的改善作用. 食品科学. 2025(02): 89-98 .  百度学术
百度学术
其他类型引用(2)
 下载:
下载:


计量
- 文章访问数: 196
- HTML全文浏览量: 33
- PDF下载量: 14
- 被引次数: 3



