


Eupatilin Attenuates Ethanol Withdrawal-Induced Anxiety-like Behavior in Rats by Improving Ventral Hippocampus GABAa Transmission
-
摘要: 目的:研究异泽兰黄素(Eupatilin,Eptl)对大鼠酒精戒断(Ethanol withdrawal,EtOHWI)焦虑样行为的改善作用及其与腹侧海马(Ventral hippocampus,vHippo)相关机制。方法:将32只成年雄性Sprague-Dawley大鼠随机分为4组,分别为生理盐水对照组、EtOHWI模型组、Eptl低剂量治疗组和Eptl高剂量治疗组,每组各8只。EtOHWI模型组、Eptl低、高剂量治疗组每天1次腹腔注射3 g/kg乙醇(Ethanol,EtOH),连续28 d,戒断3 d制备大鼠EtOHWI模型,生理盐水对照组腹腔注射等容积生理盐水。在3 d戒断期间,Eptl低、高剂量治疗组分别每天1次灌胃10和30 mg/kg Eptl,第3次给药30 min后,利用旷场(Open filed,OF)和高架十字迷宫(Elevated plus maze,EPM)实验对各组大鼠进行焦虑样行为学检测。利用ELISA检测血清皮质酮(Coritosterone,CORT)浓度、vHippo组织中GABA水平;利用实时荧光定量PCR检测vHippo组织中谷氨酸脱羧酶67(GAD 67)mRNA相对表达量;利用Western blot(WB)检测vHippo组织中GABAaRα1(GABAa receptor α1,GABAaRα1)、GABAaRα2、核因子E2相关因子2(Nrf2)、血红素加氧酶1(HO-1)蛋白表达;利用试剂盒检测vHippo组织中MDA、T-SOD、CAT、GSH以及IL-6和TNF-α的含量。利用免疫荧光检测技术观察HT22细胞的核中Nrf2蛋白水平。结果:与EtOHWI模型组相比,Eptl低、高剂量治疗组大鼠在OF中央区活动距离极显著升高(P<0.01),分别为70.62%和124.21%,活动时间显著升高(P<0.05或P<0.01),分别为251.75%和371.62%,EPM的开放臂进入次数百分比显著升高(P<0.05或P<0.01),分别为110.33%和207.32%,滞留时间百分比显著升高(P<0.05或P<0.01),分别为99.56%和184.18%;血清CORT含量极显著降低(P<0.01);vHippo组织中GABA含量和GAD67mRNA表达显著升高(P<0.05或P<0.01);GABAaRα1、GABAaRα2、Nrf2和HO-1蛋白表达显著增多(P<0.05或P<0.01);MDA水平极显著降低(P<0.01),而T-SOD、CAT和GSH的活性或水平显著升高(P<0.05或P<0.01);IL-6、TNF-α含量显著降低(P<0.05或P<0.01)。体外实验结果显示,与空白对照组相比,200 μmol/L H2O2刺激的HT22细胞核中Nrf2的水平极显著增多(P<0.01),但30 μmol/L Eptl预处理显著抑制了Nrf2水平的增多(P<0.05)。结论:Eptl具有改善大鼠EtOHWI焦虑样行为的作用,其机制可能通过Eptl的抗氧化和抗炎作用而调节EtOHWI大鼠vHippo的GABAaR传递紊乱所介导。Abstract: Objective: To study the improving effect of Eupatilin (Eptl) on ethanol withdrawal (EtOHWI)-induced anxiety-like behavior in rats and probe the mechanisms related to ventral hippocampus (vHippo). Methods: Thirty-two adult male Sprague-Dawley rats were randomly divided into four groups, 8 rats per group: Saline-treated control group, EtOHWI model group, low-dose Eptl treatment group and high-dose Eptl treatment group. The EtOHWI was established by intraperitoneal injection of 3 g/kg of ethanol (20% volume/volume, dissolved in saline) once a day for 28 days followed by 3 days of withdrawal, during the withdrawal period, the low-and the high-dose Eptl treatment groups were respectively given 10 and 30 mg/kg Eptl through oral route once a day, while the saline control group was administered with an equal volume of saline. Thirty minutes after the third Eptl, all the rats were subjected to open filed (OF) and elevated plus maze (EPM) tests to detect anxiety-like behaviors. The serum coritosterone (CORT) concentration and vHippo γ-aminobutyric acid (GABA) secretion were measured by enzyme linked immunosorbent assay (ELISA), and vHippo glutamic acid decarboxylase 67 (GAD 67) mRNA relative expression was assayed by real-time quantitative polymerase chain reaction. The protein expression of GABAa receptor α1 (GABAaRα1), GABAaRα2, nuclear factor E2-related factor 2 (Nrf2), heme oxygense-1 (HO-1) in the vHippo were analyzed by Western blot. The levels of MDA, T-SOD, CAT and GSH, IL-6 and TNF-α were measured by commercial kits. Meanwhile, in the in vitro experiment, the nuclear levels of Nrf2 in HT22 cells were detected via immunofluorescent technique. Results: Compared with the rats in the EtOHWI group, the rats in low and high-dose Eptl treatment groups moving distance increased significantly (P<0.01) in the central region of OF which was 70.62% and 124.21% respectively, and moving time increased significantly (P<0.05 or P<0.01) which was 251.75% and 371.62% respectively in the central zone of the OF. Visited more frequently (P<0.05 or P<0.01) which was 110.33% and 207.32% respectively, and stayed time increased significantly (P<0.05 or P<0.01) which was 99.56% and 184.18% respectively in the open arms of the EPM1. In biochemical assays, compared with those in EtOHWI rats, in the rats of low-dose and high-dose Eptl treatment groups, the serum CORT concentrations decreased significantly (P<0.01). The vHippo GABA and GAD67 mRNA levels increased significantly (P<0.05 or P<0.01). The protein expression of GABAaRα1, GABAaRα2, Nrf2, HO-1 in the vHippo increased significantly (P<0.05 or P<0.01). The level of MDA decreased significantly (P<0.01), while the activities of T-SOD and CAT, as well as the level of GSH increased respectively significantly (P<0.05 or P<0.01). The levels of IL-6 and TNF-α decreased significantly (P<0.05 or P<0.01). In the in vitro experiment, the immunofluorescent assay showed that compared with blank control group, Nrf2 level in nucleus of HT22 stimulated by 200 μmol/L H2O2 increased significantly (P<0.01), whereas pretreatment with 30 μmol/L Eptl inhibited the increase of Nrf2 level (P<0.05). Conclusion: Eptl attenuates EtOHWI-induced anxiety-like behavior in rats, which may be mediated by regulating the vHippo GABAaR transmissional disorder of EtOHWI rats via its antioxidant and anti-inflammatory activities.
-
酒精中毒(Alcoholism)的危害极大,但其治疗成功率极低[1]。酒精中毒患者戒酒一段时间后复饮是其治疗的最大难点,而酒精戒断(EtOH Withdrawal,EtOHWI)引起的情感障碍是导致复饮的根源之一[2]。临床研究和实验动物研究表明,EtOHWI后酒精中毒患者和实验动物均表现出显著的焦虑障碍(症),为了缓解此症状的痛苦,酒精中毒患者和实验动物会觅酒和重新饮酒[3−4]。目前临床上应用的治疗酒精中毒的药物地西泮或劳拉西泮等苯二氮卓类药物主要通过缓解患者的焦虑症而实现其治疗效果[5]。虽然此类药物表现出一定的疗效,但其本身具有较强的成瘾性[6],继而很大限制其进一步使用。很显然,研发具有良好的抗焦虑效应,并且无成瘾性的新药是目前酒精中毒治疗所面临的重大课题。
异泽兰黄素(Eupatilin,Eptl),中药艾叶中的重要黄酮类成份,也广泛存在于多种菊科蒿属类植物中[7],而艾叶在民间常被做食用原料或佐料使用[8]。已知Eptl具有良好的抗炎、抗氧化、抗肿瘤和神经保护等药理活性[9−10],并且发现Eptl通过海马(Hippocampus,Hippo)的神经传递调节和抗炎等机制抑制小鼠抑郁样行为[11−12]。海马是大脑边缘系统的重要结构,分布着大量的γ-氨基丁酸(Gamma-aminobutyric acid,GABA)能神经[13],而GABAaR是酒精的主要作用点,且GABA能神经功能紊乱是导致大量饮酒所致情感障碍的重要因素[14]。因此,本研究首先在行为和应激激素的角度观察Eptl对大鼠EtOHWI焦虑样行为的作用,以vHippo的GABA能神经传递功能变化、炎症和氧化应激等作为靶点探讨Eptl的作用机制,旨在为Eptl研发成新的抗EtOHWI焦虑症的药物提供基础研究依据。
1. 材料与方法
1.1 材料与仪器
32只雄性Sprague-Dawley(SD)大鼠 随机选取8周龄,体重均为200~220 g,由齐齐哈尔医学院动物实验中心提供,使用许可证号:SYXK(黑)2021-013。实验大鼠均在20~23 ℃,湿度40%~50%,具有自动空气清洁设备的专用IVC鼠笼饲养,自由饮水饮食。本实验已通过齐齐哈尔医学院实验动物伦理委员会批准(批号:QMU-AECC-2023-68),且所有操作均按照相关指南执行;小鼠海马神经细胞系HT22 以含10%胎牛血清、1%青霉素和链霉素的Dulbecco's modified Eagle's medium高糖培养液在37 ℃、5% CO2恒温培养箱中孵育,BeNa Culture Collection;异泽兰黄素(纯度≥98%) 成都德斯特生物科技有限公司;总蛋白测定试剂盒、MDA、CAT、T-SOD、GSH检测试剂盒、Nrf2抗体 碧云天生物技术公司;HO-1抗体 Santa Cruz Biotechnology;GABAaRα1、GABAaRα2抗体 索莱宝科技有限公司;大鼠CORT、大鼠GABA、大鼠TNF-α、大鼠IL-6 ELISA检测试剂盒 上海酶联生物科技有限公司。
RL20170223大鼠旷场(Open field,OF)实验箱、RL20170210高架十字迷宫(Elevated plus maze,EPM) 上海软隆科技发展有限公司;多功能酶标仪 奥地利Tecan公司;1658001小型垂直电泳槽、1703935电泳转印系统 美国BIO-RAD公司;UVP Chemstudio touch多功能成像仪、qTOWER3G荧光定量梯度基因扩增仪 德国Analytikjena公司。
1.2 实验方法
1.2.1 动物模型的分组、模型建立和给药
适应性饲养1周后,将实验大鼠随机分为4组,分别为生理盐水对照组、酒精戒断(EtOHWI)模型组、Eptl低剂量治疗组和Eptl高剂量治疗组,每组各8只。除生理盐水对照组外,其余各组每天1次腹腔注射3 g/kg EtOH(稀释于生理盐水,20%的容积比),共28 d。戒断3 d制备EtOHWI模型,生理盐水对照组以生理盐水代替EtOH[15]。在戒断3 d期间,每天对Eptl低剂量治疗组(10 mg/kg)和Eptl高剂量治疗组(30 mg/kg)分别灌胃1次[16](Eptl灌胃前先溶解于二甲基亚砜,后用生理盐水进行稀释)。生理盐水对照组和EtOHWI模型组灌胃生理盐水。在第3次给药30 min后,对各组大鼠进行OF和EPM检测,观察行为学变化。
1.2.2 行为学检测
旷场试验(OF):首先,将各组大鼠放置于旷场实验箱的正中央区域(50 cm×50 cm×50 cm),采用视频系统开始记录5 min内大鼠的活动情况。观察指标为:中央区域的活动距离(mm)及活动时间(s)[17];高架十字迷宫试验(EPM):EPM由从地面50 cm升高的十字形长宽(50 cm×10 cm)相同的开放臂(无侧壁)和闭合臂(有侧壁)垂直交叉构成。OF检测结束10 min后进行EPM实验,将各组大鼠放置于迷宫中央区域,头均朝向开放臂方向,采用视频系统开始记录5 min内大鼠的活动情况。观察指标为:开放臂进入次数百分比(%)和开放臂滞留时间百分比(%)。行为学检测期间实验室保持通风安静,每只大鼠检测后,用0.1%醋酸擦干仪器除味[18]。
开放臂进入次数百分比(%)=开放臂进入次数开放臂进入次数+闭合臂进入次数×100 开放臂滞留时间百分比(%)=开放臂滞留时间总时间×100 1.2.3 血清中CORT含量测定
行为学检测结束后,大鼠安乐死后收集血液至1.5 mL离心管中,在4 ℃、3000 r/min离心15 min后收集血清,严格按照ELISA试剂盒说明书检测CORT水平,结果以ng/mL表示。
1.2.4 vHippo中GABA含量测定
大鼠安乐死处理后取出大脑,根据大鼠脑定位图谱[19],分离出vHippo并称重,按5%的比例于PBS中充分匀浆,8000 r/min离心10 min,收集上清严格按照大鼠GABA测定试剂盒说明书检测GABA水平,结果以μmol/L表示。
1.2.5 Western blot检测vHippo中GABAaRα1、GABAaRα2、Nrf2、HO-1蛋白表达
将收集的vHippo于RIPA蛋白裂解液(含有蛋白酶抑制剂)中充分研磨,12000 r/min离心5 min取上清液,用BCA试剂盒进行蛋白定量,并按照常规操作方法进行电泳、转膜、封闭、Ⅰ抗4 ℃孵育过夜,再加入Ⅱ抗室温摇床孵育2 h后,用ECL显色,利用多功能凝胶成像系统扫描并用ImageJ软件进行灰度值分析,以GAPDH作为内参对照。
1.2.6 vHippo中氧化应激指标和炎症因子的测定
将收集的vHippo按10%的比例于PBS中充分匀浆,3000 r/min离心20 min,收集上清严格按试剂盒说明书分别检测vHippo中氧化应激指标MDA、T-SOD、CAT和GSH水平以及炎症因子IL-6和TNF-α。
1.2.7 qPCR检测vHippo中GAD67 mRNA相对表达量
将收集的vHippo采用Trizol Reagent法提取RNA并检测其浓度和纯度,严格按照试剂盒说明书进行逆转录合成cDNA。基因引物序列如下:GAD67,Forward:CATCATGGCGGCTGGTACACAG;Reverse:GTGACTGTGTTCTGAGGTGAAGAGG。GAPDH,Forward:AATCCTGGGCGGTACAACTC;Reverse:GTTCACACCGACCTTCACCA。qPCR反应条件为预变性95 ℃、3 min,变性95 ℃、15 S,退火55 ℃、30 S,延伸72 ℃、20 S,共进行43个循环,然后计算△Ct=Ct目的基因−Ct内参基因,△△Ct=处理组△Ct−对照组△Ct,最后计算2−△△Ct值,然后进行统计分析。
1.2.8 MTT法检测HT22细胞活力
根据文献[20]的方法,收集生长状态良好的HT22细胞,制备细胞悬液,以每孔4×103个接种于96孔板中,每孔100 μL,加入不同浓度(50、100、200、400、800、1000 μmol/L)的H2O2培养24 h。准备相同细胞加入不同浓度(3、10、30、60 μmol/L)的Eptl培养24 h。接种相同细胞,预处理组加入30 μmol/L的Eptl 预处理2 h,与H2O2模型组同时给予200 μmol/L H2O2刺激,培养24 h,每组均设3个复孔。然后加入10 μL MTT染色液,于37 ℃、5% CO2恒温培养箱中继续孵育4 h,每孔加100 μL Formanzan溶解液继续孵育5 h待结晶完全溶解后用酶标仪检测570 nm处吸光度(OD)值,并计算细胞存活力,筛选最佳H2O2和Eptl处理浓度。
1.2.9 HT22细胞分组、给药以及免疫荧光实验
收集生长状态良好的HT22细胞,爬片到经组织培养物处理的圆形玻片上,分为Control组(空白组)、H2O2模型组(200 μmol/L H2O2)和Eptl预处理组(30 μmol/L Eptl+200 μmol/L H2O2)。Eptl预处理组以30 μmol/L Eptl预处理2 h后,与模型组同时给予200 μmol/L H2O2培养24 h,空白组用正常培养基培养。造模完成后经PBS润洗3 min,10%中性甲醛固定40 min,0.1% Triton-X穿透10 min,5% BSA封闭0.5 h,加入Nrf2 Ⅰ抗4 ℃孵育过夜,Ⅱ抗室温孵育2 h,DAPI核染色10 min,抗荧光淬灭剂封片后于倒置荧光显微镜观察。
1.3 数据处理
采用GraphPad Prism 8.0统计软件进行数据分析,数据均采用均数±标准差(Mean±SD)表示,多组间比较采用单因素方差分析(One-way,ANOVA),组间两两比较采用Newman-Keuls法进行检验,P<0.05为差异具有统计学意义。
2. 结果与分析
2.1 Eptl对EtOHWI大鼠行为学变化的影响
OF是利用啮齿类动物对开阔的新异环境的恐惧和探究能力来反映动物的自发活动情况和焦虑心理。EPM是利用啮齿类动物对新异环境的探究能力和对高悬敞开臂的恐惧形成矛盾冲突行为来检测动物的焦虑状态[21−22]。由表1可知,在OF检测中,EtOHWI模型组大鼠在中央区域的活动距离和活动时间均极显著低于生理盐水对照组(P<0.01);与EtOHWI组比较,Eptl低、高剂量治疗组大鼠在OF中央区活动距离极显著增加(P<0.01),分别为70.62%和124.21%,活动时间显著增加(P<0.05或P<0.01),分别为251.75%和371.62%;在EPM检测中,EtOHWI模型组大鼠开放臂进入次数和开放臂滞留时间百分比均极显著低于(P<0.01)生理盐水对照组;与EtOHWI模型组比较,Eptl低、高剂量治疗组大鼠开放臂进入次数百分比显著升高(P<0.05或P<0.01),分别为110.33%和207.32%,滞留时间百分比显著增加(P<0.05或P<0.01),分别为99.56%和184.18%,表现出大鼠对开放臂的探索趋势显著升高;在组间两两比较中,Eptl高剂量治疗组大鼠开放臂进入次数和开放臂滞留时间百分比均显著高于(P<0.05)低剂量治疗组,说明Eptl对EtOHWI大鼠焦虑样行为的治疗具有剂量依赖性。上述检测指标提示,大鼠EtOHWI焦虑样模型制备成功。在EtOHWI期给予Eptl治疗后,焦虑样行为显著减轻。
表 1 Eptl对EtOHWI大鼠行为学变化的影响(n=8)Table 1. Effect of eupatilin on the changes of EtOHWI rats (n=8)组别 旷场试验 高架十字迷宫试验 中央区活动距离(mm) 中央区活动时间(s) 开放臂进入次数百分(%) 开放臂滞留时间百分比(%) 生理盐水对照组 232.07±22.52 16.6±7.09 27.37±4.69 29.51±6.69 EtOHWI模型组 101.73±19.71## 3.30±1.23## 8.97±2.97## 10.22±3.27## Eptl低剂量治疗组 173.57±12.03** 15.54±2.51* 18.87±6.95* 20.39±4.62* Eptl高剂量治疗组 228.09±23.32**& 16.74±4.86** 27.58±4.86**& 29.03±6.11**& 注:与生理盐水对照组比较,##表示有极显著差异P<0.01;与EtOHWI模型组比较,*表示有显著差异P<0.05,**表示有极显著差异P<0.01;与Eptl低剂量治疗组比较,&表示有显著差异P<0.05;表2~表3,图1~图2同。 2.2 Eptl对EtOHWI大鼠血清CORT水平、vHippo中GABA含量的影响
下丘脑-垂体-肾上腺轴(Hypothalamic pituitary adrenal axis,HPA)是机体内最为重要的神经内分泌系统之一。HPA轴中CORT含量变化可作为直接评价啮齿类动物焦虑程度的重要指标,两者呈正相关[23]。如表2所示,各组大鼠血清CORT检测结果如下。与生理盐水对照组比较,EtOHWI模型组大鼠血清CORT含量极显著升高(P<0.01);与EtOHWI模型组比较,Eptl低、高剂量治疗组血清CORT水平均极显著降低(P<0.01);组间两两比较显示,Eptl低、高剂量治疗组间CORT水平差异具有统计学意义(P<0.01),表明两者具有极显著的剂量依赖性,说明Eptl显著降低了EtOHWI大鼠血清CORT的上升趋势,减轻了对EtOH戒断焦虑样行为。GABA是大脑中广泛分布的抑制性神经递质,EtOH与GABA相互作用会增强EtOH的负性强化作用,在EtOH成瘾和依赖中起着重要作用。苯二氮卓类药物镇静、抗焦虑和抗惊厥作用与GABA系统的激活有关,本研究结果与之呼应[24]。ELISA检测结果显示,与生理盐水对照组比较,EtOHWI模型组大鼠vHippo中GABA含量极显著降低(P<0.01);与EtOHWI模型组比较,Eptl低、高剂量治疗组vHippo中GABA含量均显著升高(P<0.05或P<0.01)。此结果说明,在EtOHWI大鼠发病过程中vHippo中GABA的降低会导致焦虑样行为,而Eptl可通过上调vHippo中GABA水平改善EtOHWI大鼠焦虑样行为。
表 2 Eptl对EtOHWI大鼠血清CORT水平、vHippo中GABA含量的影响(n=8)Table 2. Effects of eupatilin on the level of serum CORT and the ventral hippocampal GABA content concentration of EtOHWI rats (n=8)组别 CORT(ng/mL) GABA(μmol/L) 生理盐水对照组 53.91±7.01 36.65±3.58 EtOHWI模型组 121.07±12.77## 19.68±3.53## Eptl低剂量治疗组 89.15±6.62** 29.78±7.36* Eptl高剂量治疗组 57.92±9.00**&& 34.48±3.92** 2.3 Eptl对EtOHWI大鼠vHippo中GABAaRα1、GABAaRα2、Nrf2、HO-1蛋白表达的影响
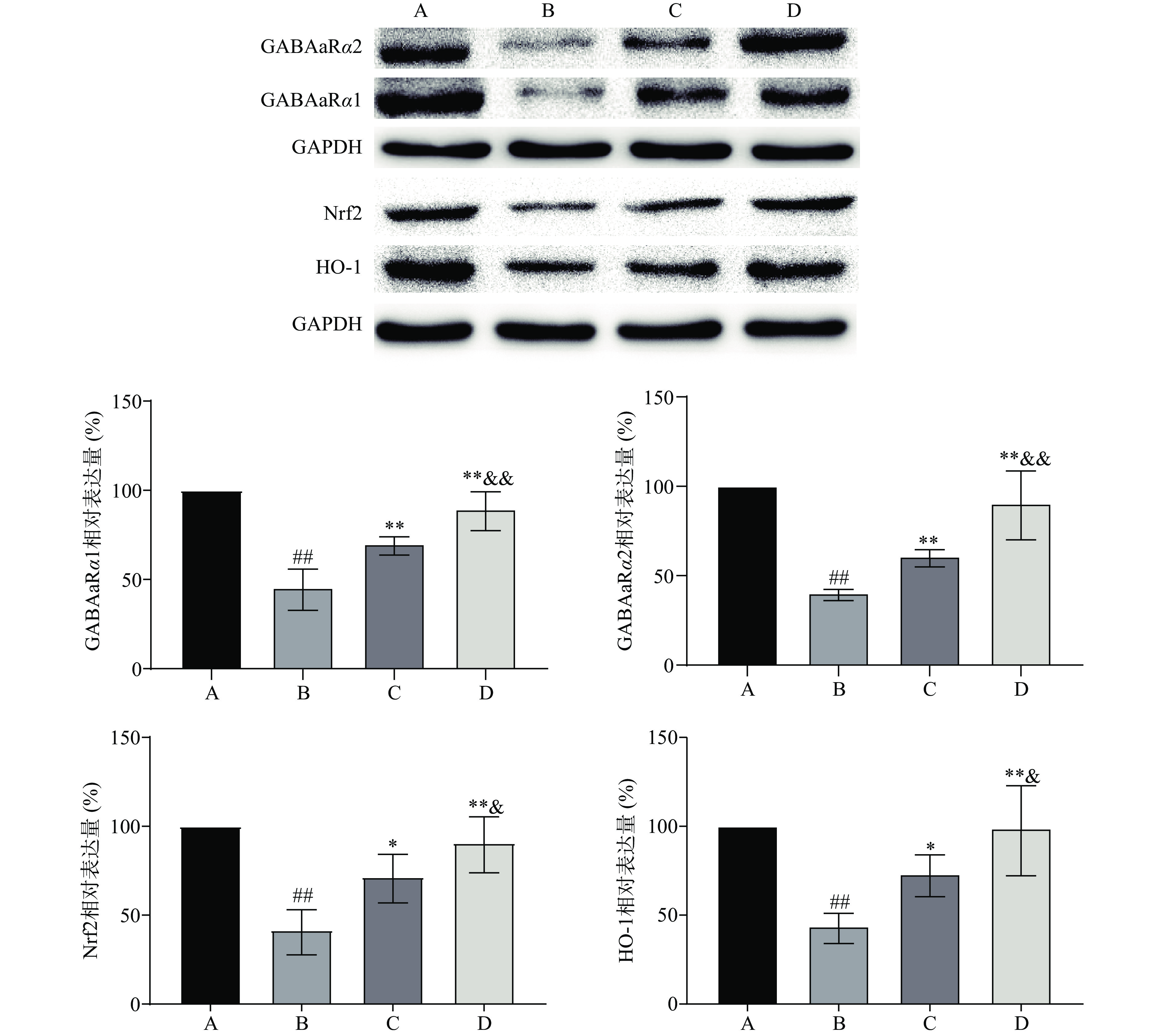
GABAaR是维持和控制大脑神经元兴奋的关键蛋白,GABA通过与其受体GABAaR结合发挥生理功能,焦虑症的发生与GABAaRα1、GABAaRα2的改变密切相关[25−26]。长期的EtOH依赖或戒断会导致机体处于氧化应激状态,其中Nrf2/HO-1是氧化应激反应重要的信号通路。Western blot检测结果如图1,与生理盐水对照组比较,EtOHWI模型组大鼠vHippo中GABAaRα1、GABAaRα2、Nrf2和HO-1蛋白表达极显著减少(P<0.01);与EtOHWI模型组比较,Eptl低、高剂量治疗组大鼠vHippo中GABAaRα1、GABAaRα2、Nrf2和HO-1蛋白表达显著升高(P<0.05或P<0.01);组间两两比较显示,Eptl低、高剂量治疗组间GABAaRα1、GABAaRα2、Nrf2和HO-1蛋白表达均有统计学意义(P<0.05或P<0.01),具有剂量依赖性。由此可推断,GABAaRα1、GABAaRα2蛋白下调可能是介导EtOHWI焦虑症的重要机制,而Eptl能纠正其紊乱发挥抑制EtOHWI焦虑样行为的效应。Nrf2/HO-1抗氧化系统可能参与了Eptl的抗氧化过程从而保护了vHippo氧化应激的损伤。
![]() 图 1 各组大鼠vHippo中GABAaRα1、GABAaRα2、Nrf2、HO-1蛋白表达(n=8)注:A:生理盐水对照组;B:EtOHWI模型组;C:Eptl低剂量治疗组;D:Eptl高剂量治疗组;图2同。Figure 1. Protein expression of ventral hippocampal GABAaRα1, GABAaRα2, Nrf2 and HO-1 of each group rats (n=8)
图 1 各组大鼠vHippo中GABAaRα1、GABAaRα2、Nrf2、HO-1蛋白表达(n=8)注:A:生理盐水对照组;B:EtOHWI模型组;C:Eptl低剂量治疗组;D:Eptl高剂量治疗组;图2同。Figure 1. Protein expression of ventral hippocampal GABAaRα1, GABAaRα2, Nrf2 and HO-1 of each group rats (n=8)2.4 Eptl对EtOHWI大鼠vHippo中氧化应激指标和炎症因子水平的影响
各组实验大鼠vHippo中氧化应激指标和炎症因子的变化如表3所示,与生理盐水对照组比较,EtOHWI模型组大鼠vHippo中MDA水平呈极显著升高(P<0.01),而T-SOD、CAT、GSH活性极显著降低(P<0.01));与EtOHWI模型组比较,Eptl低、高剂量治疗组大鼠vHippo中MDA水平极显著降低(P<0.01),T-SOD、CAT、GSH活性显著升高(P<0.05或P<0.01);在组间两两比较中,Eptl高剂量治疗组大鼠较Eptl低剂量治疗组vHippo中MDA水平显著降低(P<0.05),有剂量依赖性。在炎症因子的检测中,EtOHWI模型组大鼠vHippo中IL-6、TNF-α含量较生理盐水对照组极显著升高(P<0.01),而Eptl低、高剂量治疗组大鼠vHippo中IL-6、TNF-α含量较EtOHWI模型组大鼠显著降低(P<0.05或P<0.01);在组间两两比较中,Eptl高剂量治疗组大鼠较Eptl低剂量治疗组vHippo中TNF-α显著降低(P<0.05),表现出剂量依赖性。因此可以推断,EtOHWI期大鼠vHippo中发生了氧化应激和炎症反应,失去了内环境稳态的动态平衡,诱导了vHippo中GABA能神经传递的紊乱,进一步说明EtOHWI焦虑症发病机制中大脑vHippo中氧化应激和炎症反应的重要作用。Eptl治疗后vHippo中氧化应激指标和炎症因子水平紊乱得到显著改善,缓解了EtOHWI对大鼠vHippo的损伤。
表 3 Eptl对EtOHWI大鼠vHippo氧化应激指标和炎症因子水平的影响(n=8)Table 3. Effects of eupatilin on the oxidative stress and inflammatory factors in ventral hippocampal of EtOHWI rats (n=8)组别 氧化应激指标 炎症因子 MDA
[μmol/(g protein)]T-SOD
[×103 U/(g protein)]CAT
[×103 U/(g protein)]GSH
[μmol/(mg protein)]IL-6
(pg/mg)TNF-α
(pg/mg)生理盐水对照组 1.54±0.19 60.55±9.11 86.38±8.14 3.71±1.11 6.90±1.15 4.02±0.67 EtOHWI模型组 3.53±0.62## 31.90±6.11## 38.60±6.81## 1.62±0.51## 13.04±2.67## 8.22±1.29## Eptl低剂量治疗组 2.23±0.49** 48.21±4.44** 71.50±12.14** 3.02±0.76* 8.42±1.60** 5.85±1.44* Eptl高剂量治疗组 1.52±0.34**& 55.99±5.82** 82.12±12.57** 3.37±0.93* 6.92±0.43** 3.98±0.76**& 2.5 Eptl对EtOHWI大鼠vHippo中GAD67 mRNA相对表达量的影响
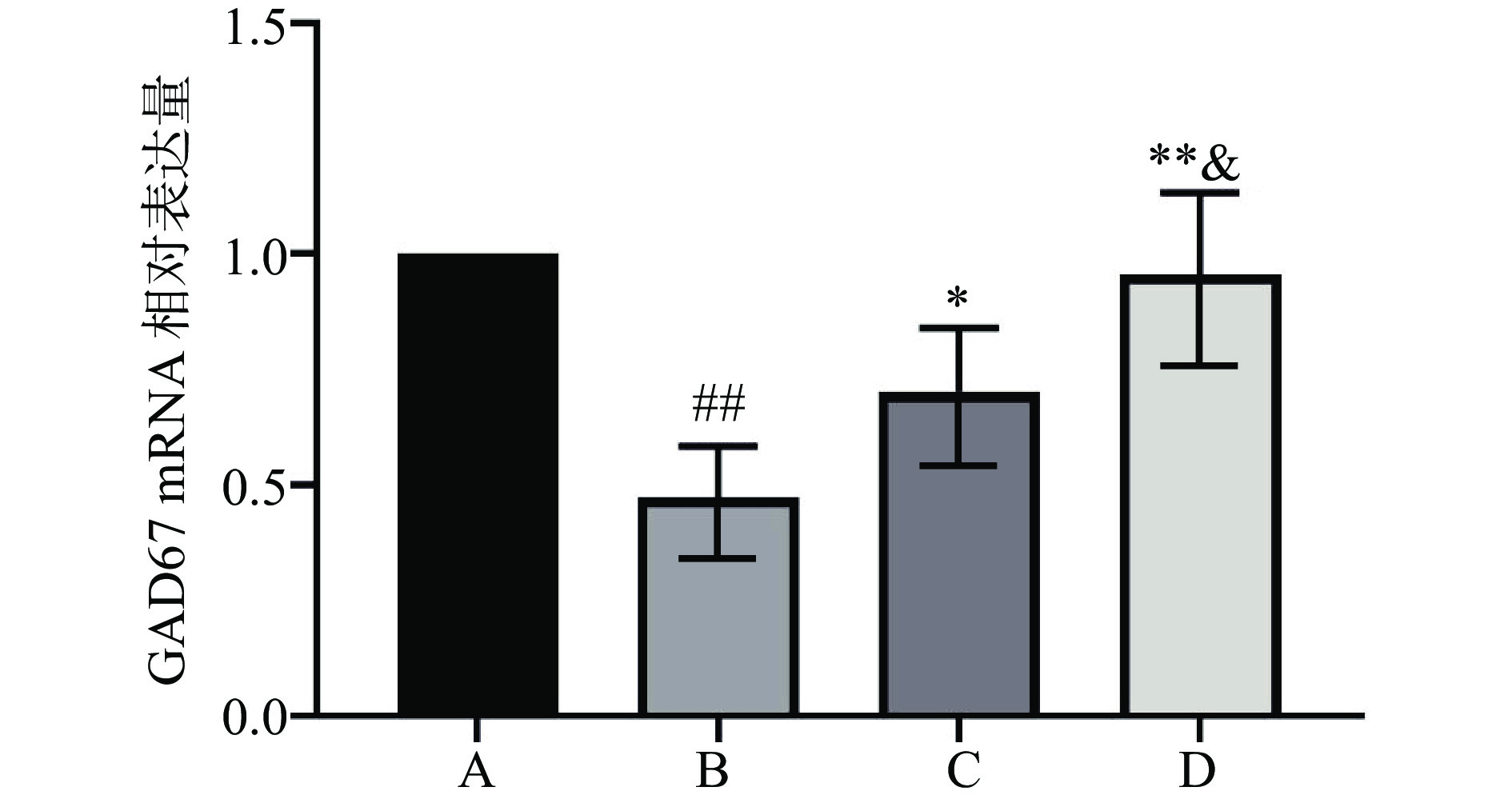
qPCR检测结果如图2所示,与生理盐水对照组比较,EtOHWI模型组大鼠vHippo中GAD67 mRNA相对表达量极显著减少(P<0.01);与EtOHWI模型组比较,Eptl低、高剂量治疗组大鼠vHippo中GAD67 mRNA相对表达量显著升高(P<0.05或P<0.01);组间两两比较显示,Eptl低、高剂量治疗组间GAD67 mRNA相对表达量差异具有统计学意义(P<0.05),具有剂量依赖性。此结果进一步说明了GABAa神经传递紊乱可能是介导EtOHWI焦虑症的重要机制,给予Eptl可以恢复GABAa神经的生理功能,减轻EtOHWI给机体带来的损伤,缓解焦虑症状。
![]() 图 2 各组大鼠vHippo中GAD67 mRNA相对表达量(n=8)Figure 2. Relative expression of ventral hippocampal GAD67 mRNA of each group rats (n=8)
图 2 各组大鼠vHippo中GAD67 mRNA相对表达量(n=8)Figure 2. Relative expression of ventral hippocampal GAD67 mRNA of each group rats (n=8)2.6 Eptl对H2O2诱导的HT22细胞活力的影响
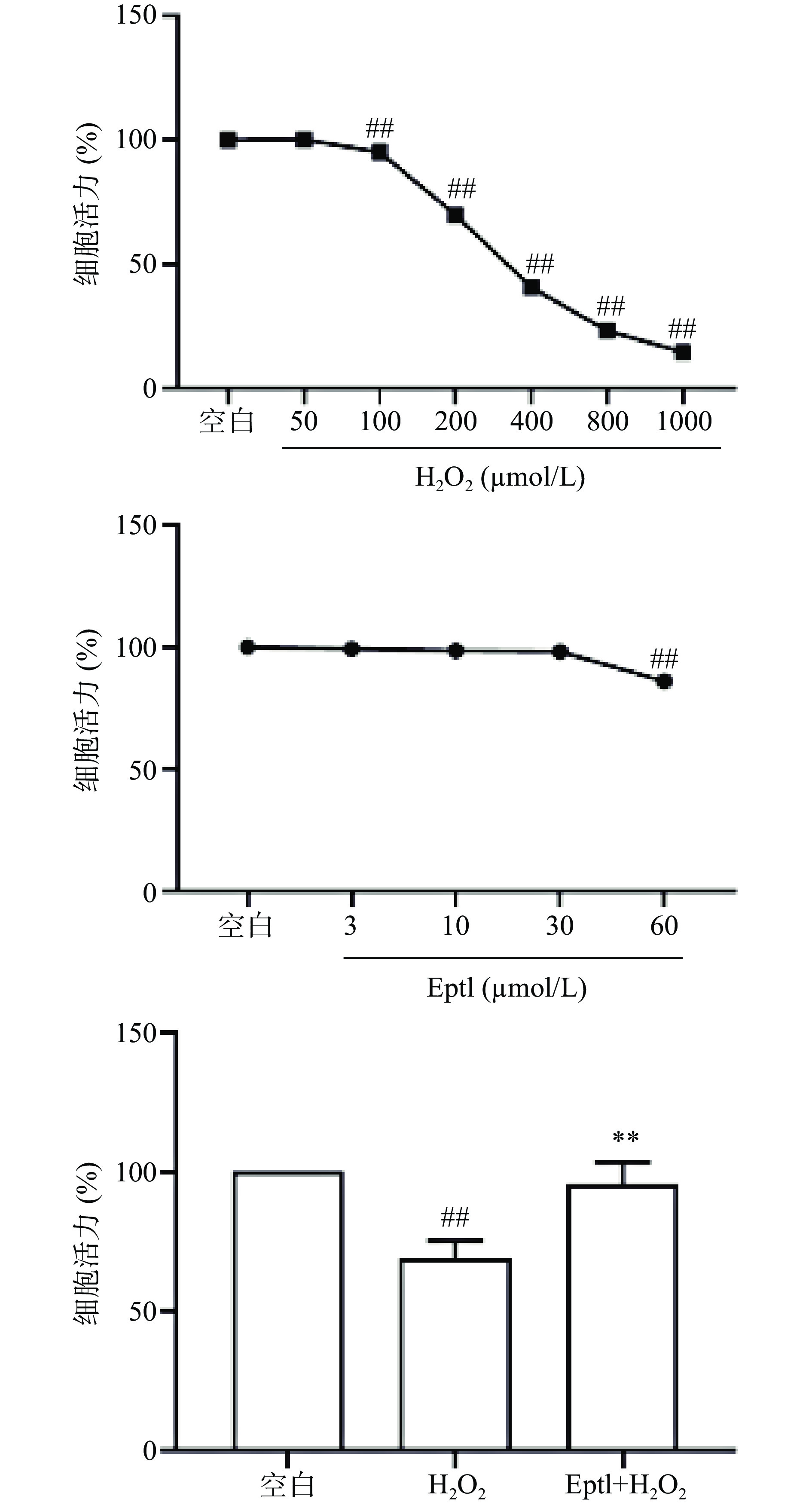
如图3所示,H2O2和Eptl单独处理后观察对HT22细胞活力的影响,H2O2最适刺激浓度为200 μmol/L,Eptl预处理浓度为30 μmol/L。与空白组比较,H2O2模型组细胞活力极显著降低(P<0.01);与H2O2模型组比较,30 μmol/L Eptl预处理组的细胞活力显著升高(P<0.01)。
![]() 图 3 Eptl对H2O2诱导的HT22细胞活力的影响(n=3)注:与空白组比较,##表示有极显著差异P<0.01;与H2O2模型组比较,**表示有极显著差异P<0.01。Figure 3. Effect of eupatilin on the viability of H2O2-induced HT22 cells (n=3)
图 3 Eptl对H2O2诱导的HT22细胞活力的影响(n=3)注:与空白组比较,##表示有极显著差异P<0.01;与H2O2模型组比较,**表示有极显著差异P<0.01。Figure 3. Effect of eupatilin on the viability of H2O2-induced HT22 cells (n=3)2.7 Eptl对H2O2诱导的HT22细胞中Nrf2表达的影响
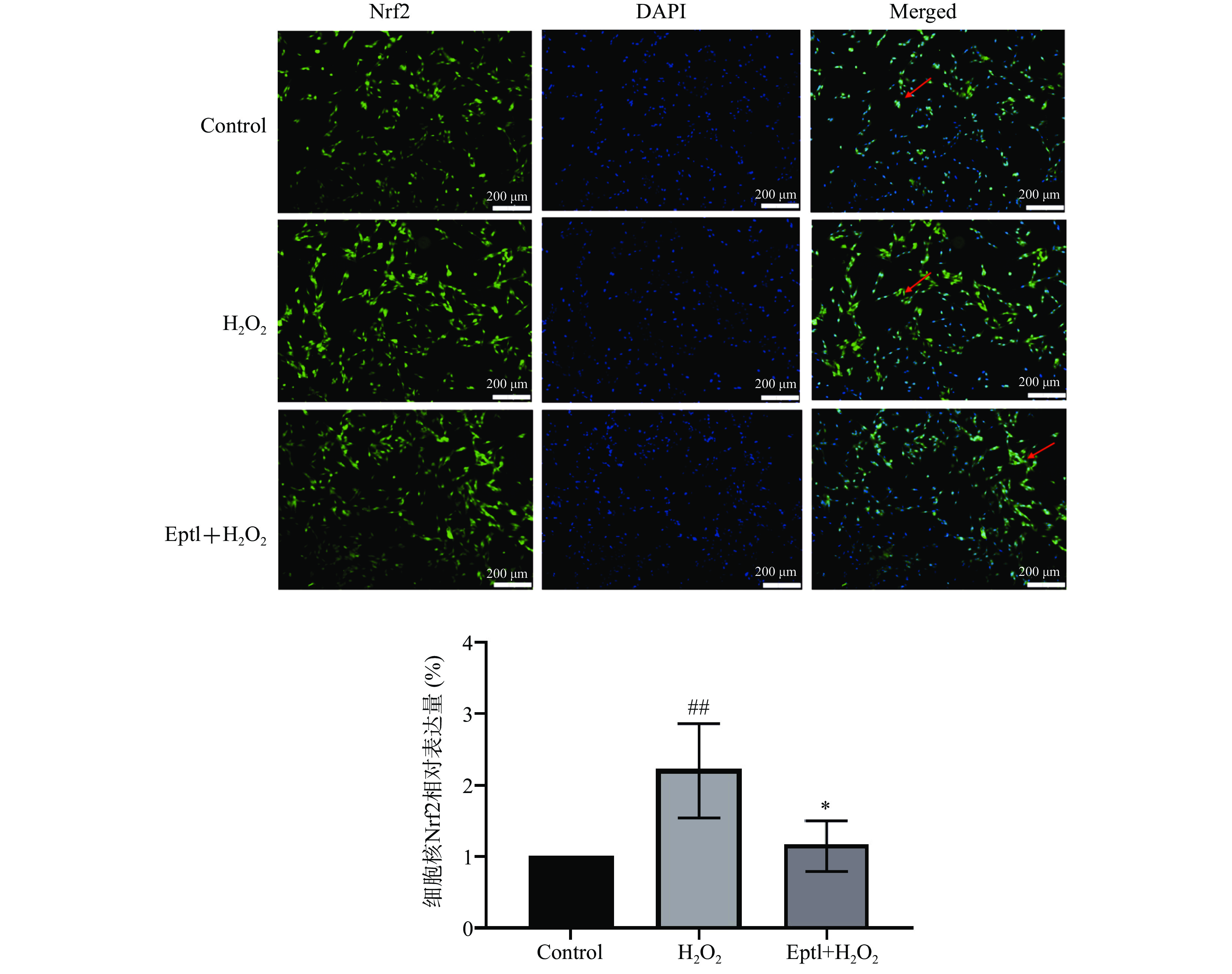
如图4所示,Control(空白)组中,Nrf2主要存在于胞质内,胞核中表达极少,而H2O2模型组胞核中Nrf2的表达较Control组极显著升高(P<0.01);与H2O2模型组比较,Eptl预处理组细胞核中Nrf2的表达显著减少(P<0.05),且与Control(空白)组比较,并无显著性差异(P>0.05)。
![]() 图 4 Eptl对H2O2诱导的HT22细胞中Nrf2表达的影响(200×,n=3)注:与Control组比较,##表示有极显著差异P<0.01;与H2O2模型组比较,*表示有显著差异P<0.05。Figure 4. Effect of eupatilin on the expression of H2O2-induced HT22 cells (200×, n=3)
图 4 Eptl对H2O2诱导的HT22细胞中Nrf2表达的影响(200×,n=3)注:与Control组比较,##表示有极显著差异P<0.01;与H2O2模型组比较,*表示有显著差异P<0.05。Figure 4. Effect of eupatilin on the expression of H2O2-induced HT22 cells (200×, n=3)3. 讨论与结论
酒精中毒会引起酒精依赖,戒断会导致睡眠障碍、焦虑和抑郁等精神症状,给社会和个人的生活带来严重的危害[27]。本研究发现EtOHWI期给予Eptl对抑制大鼠的EtOHWI焦虑样行为有积极作用。EtOHWI 期血液中 CORT 水平增高被认为是大鼠焦虑的激素学基础[27]。本研究中观察到EtOHWI大鼠血清CORT水平的极显著升高(P<0.01) ,且两个剂量的Eptl均可有效地抑制其增高,此结果不仅从激素学角度支持了Eptl抗EtOHWI焦虑的行为学的作用,而且与行为学检测结果相印证初步确定了Eptl对EtOHWI焦虑样行为的改善作用。
本研究在不同角度探讨Eptl的抗焦虑作用机制,其中vHippo作为EtOHWI焦虑症的解剖组织学靶点,解释了Eptl作用机制。Hippo是学习记忆等认知功能的关键部位,也是介导焦虑等情感障碍的重要结构,分布有大量的GABAaR[28−29]。在中枢神经系统(Central nervous system,CNS),GABA从突触前膜被释放后,与突触后膜的GABAaR结合而发挥抑制神经传递作用,因此,GABA合成和释放以及GABAaR表达直接影响GABAaR传递效应[30−31]。GAD65和GAD67是中枢GABA合成的限速酶,后者一般以功能饱和状态存在,主要负责胞浆内的GABA合成,但前者多数受到一些刺激后才被激活发挥作用。因此,组织的GABA水平主要由GAD67表达量所决定[32−33]。研究发现,情感障碍患者的Hippo存在GAD67的显著下调并伴有GABAaR传递异常[34],表明GABA合成能力下降是其发病机制之一。在Hippo,GABAaR和GABAbR是介导GABA神经传递的主要受体,其中GABAaR作为抑制性离子通道受体的代表,是介导EtOH中枢作用的重要物质基础。GABAaR是由5个亚基围绕一个中心氯离子通道的五聚体,有19种亚型,分布于不同的中枢神经系统,具有特定的生理功能[35]。苯二氮卓类药物正是通过增强GABA 与其受体的结合发挥镇静催眠和抗焦虑作用,尤其在海马中体现[36−37]。在本研究中,EtOHWI大鼠模型vHippo的GABA水平极显著降低(P<0.01), 并伴有GAD67 mRNA相对表达量的减少,表明在EtOHWI期,GABA合成与释放减少,同时,EtOHWI大鼠vHippo中GABAaR的α1和α2亚基蛋白表达量均极显著减少(P<0.01) ,这些结果与上述其它研究者的结果一致。综上,vHippo的GABAa神经传递功能下降是介导EtOHWI焦虑症重要机制,而Eptl通过改善其发挥抗EtOHWI焦虑症的效应。
炎症反应和氧化应激存在相互作用,它们所致的损伤通常被认为是,长期饮用EtOH 或EtOHWI 导致的CNS(包括Hippo )功能紊乱的潜在因素。Jeong等[12]曾在研究中发现,Stillen(Eptl的别名)可降低Hippo的IL-6和TNF-α等炎症因子的表达以发挥抗小鼠抑郁症的效应。在本研究发现,EtOHWI大鼠模型vHippo的IL-6和TNF-α与生理盐水对照组相比,极显著升高(P<0.01) ,而戒断期低高剂量Eptl的治疗均显著抑制了这些现象,表明Eptl良好的抗炎作用可能是保护或逆转长期饮用EtOH或戒断期vHippo的GABAa神经传递损伤的基本机制之一。与此相似,Tiwari等[38]的研究也发现,长期EtOH引起Hippo的氧化应激损伤因子的表达增多,而白藜芦醇通过缓解氧化应激损伤等机制改善长期EtOH所致大鼠认知障碍。同样,在本研究中,检测到EtOHWI模型大鼠vHippo的MDA水平增高,而CAT和T-SOD的活性降低,且Eptl低、高剂量治疗显著改善氧化应激损伤,表明Eptl的抗氧化应激作用也可能是它保护vHippo的GABAa神经传递治疗EtOHWI焦虑症的重要机制之一。Nrf2/HO-1是细胞中重要的抗氧化应激系统,本研究检测了vHippo的HO-1和Nrf2的蛋白表达,结果发现EtOHWI模型大鼠vHippo的此两种蛋白表达有极显著降低(P<0.01),但Eptl治疗显著提高了它们的表达(P<0.05),表明Nrf2/HO-1抗氧化系统可能参与Eptl的抗氧化过程,此结果与课题组前期的研究相符合[20],也在本研究的细胞实验中进一步得到了证实。在利用啮齿类Hippo来源的HT22细胞进行的荧光免疫染色中发现,H2O2诱导组的HT22细胞中Nrf2向细胞核内的进入量极显著增多(P<0.01),通常Nrf2存在于胞浆内,与胞质接头蛋白(Kelch-like ECH-associated protein 1,Keap1)结合在一起。当氧化应激或一些信号因子存在的情况下,Nrf2与Keap1分离进入到胞核内与抗氧化反应元件(Antioxident responseelement,ARE)结合启动HO-1和NQO-1等抗氧化因子的转录过程[39]。因此,此结果显示HT22受到氧化应激攻击时的细胞保护反应,但在同样的免疫荧光染色中发现,30 μmol/L Eptl预处理的HT22细胞中Nrf2进入胞核的量反而减少,说明此浓度的Eptl预处理通过某种机制减弱了H2O2诱导的氧化应激的攻击,保护了HT22受到氧化应激的损伤。上述细胞实验结果和组织中 Nrf2、 HO-1蛋白表达的结果综合说明了,EtOHWI 期间存在 vHippo 的抗氧化应激保护通路紊乱,而Eptl通过纠正和改善这些紊乱缓解氧化应激对GABAa传递的损伤,进而表现出抑制EtOHWI焦虑症的疗效。
综上所述,本研究得出EtOHWI期给予Eptl能缓解EtOHWI大鼠焦虑样行为,这种效应可能通过 Eptl 的抗炎抗氧化作用所介导,从而改善大鼠 vHippo的 GABAaR 传递紊乱。本研究的实验结果为Eptl可能成为治疗EtOHWI焦虑症及酒精中毒的新药研发提供新的基础实验数据和依据,且考虑到艾叶是民间常用的食用原料和佐料,本研究的结果也为艾叶的药用和保健作用增添新的科学依据。
-
![]()
图 1 各组大鼠vHippo中GABAaRα1、GABAaRα2、Nrf2、HO-1蛋白表达(n=8)
注:A:生理盐水对照组;B:EtOHWI模型组;C:Eptl低剂量治疗组;D:Eptl高剂量治疗组;图2同。
Figure 1. Protein expression of ventral hippocampal GABAaRα1, GABAaRα2, Nrf2 and HO-1 of each group rats (n=8)
![]()
图 2 各组大鼠vHippo中GAD67 mRNA相对表达量(n=8)
Figure 2. Relative expression of ventral hippocampal GAD67 mRNA of each group rats (n=8)
![]()
图 3 Eptl对H2O2诱导的HT22细胞活力的影响(n=3)
注:与空白组比较,##表示有极显著差异P<0.01;与H2O2模型组比较,**表示有极显著差异P<0.01。
Figure 3. Effect of eupatilin on the viability of H2O2-induced HT22 cells (n=3)
![]()
图 4 Eptl对H2O2诱导的HT22细胞中Nrf2表达的影响(200×,n=3)
注:与Control组比较,##表示有极显著差异P<0.01;与H2O2模型组比较,*表示有显著差异P<0.05。
Figure 4. Effect of eupatilin on the expression of H2O2-induced HT22 cells (200×, n=3)
表 1 Eptl对EtOHWI大鼠行为学变化的影响(n=8)
Table 1 Effect of eupatilin on the changes of EtOHWI rats (n=8)
组别 旷场试验 高架十字迷宫试验 中央区活动距离(mm) 中央区活动时间(s) 开放臂进入次数百分(%) 开放臂滞留时间百分比(%) 生理盐水对照组 232.07±22.52 16.6±7.09 27.37±4.69 29.51±6.69 EtOHWI模型组 101.73±19.71## 3.30±1.23## 8.97±2.97## 10.22±3.27## Eptl低剂量治疗组 173.57±12.03** 15.54±2.51* 18.87±6.95* 20.39±4.62* Eptl高剂量治疗组 228.09±23.32**& 16.74±4.86** 27.58±4.86**& 29.03±6.11**& 注:与生理盐水对照组比较,##表示有极显著差异P<0.01;与EtOHWI模型组比较,*表示有显著差异P<0.05,**表示有极显著差异P<0.01;与Eptl低剂量治疗组比较,&表示有显著差异P<0.05;表2~表3,图1~图2同。  下载: 导出CSV
下载: 导出CSV
表 2 Eptl对EtOHWI大鼠血清CORT水平、vHippo中GABA含量的影响(n=8)
Table 2 Effects of eupatilin on the level of serum CORT and the ventral hippocampal GABA content concentration of EtOHWI rats (n=8)
组别 CORT(ng/mL) GABA(μmol/L) 生理盐水对照组 53.91±7.01 36.65±3.58 EtOHWI模型组 121.07±12.77## 19.68±3.53## Eptl低剂量治疗组 89.15±6.62** 29.78±7.36* Eptl高剂量治疗组 57.92±9.00**&& 34.48±3.92**
下载: 导出CSV
表 3 Eptl对EtOHWI大鼠vHippo氧化应激指标和炎症因子水平的影响(n=8)
Table 3 Effects of eupatilin on the oxidative stress and inflammatory factors in ventral hippocampal of EtOHWI rats (n=8)
组别 氧化应激指标 炎症因子 MDA
[μmol/(g protein)]T-SOD
[×103 U/(g protein)]CAT
[×103 U/(g protein)]GSH
[μmol/(mg protein)]IL-6
(pg/mg)TNF-α
(pg/mg)生理盐水对照组 1.54±0.19 60.55±9.11 86.38±8.14 3.71±1.11 6.90±1.15 4.02±0.67 EtOHWI模型组 3.53±0.62## 31.90±6.11## 38.60±6.81## 1.62±0.51## 13.04±2.67## 8.22±1.29## Eptl低剂量治疗组 2.23±0.49** 48.21±4.44** 71.50±12.14** 3.02±0.76* 8.42±1.60** 5.85±1.44* Eptl高剂量治疗组 1.52±0.34**& 55.99±5.82** 82.12±12.57** 3.37±0.93* 6.92±0.43** 3.98±0.76**&
下载: 导出CSV
-
[1] FREED C R. The natural history of alcoholism:Causes, patterns, and paths to recovery-the virtues of an interdisciplinary perspective of alcoholism and alcoholism recovery[J]. Addiction,2022,117(2):506−509. doi: 10.1111/add.15644
[2] KIM H J, LEE M Y, KIM G R, et al. Korean Red Ginseng extract attenuates alcohol-induced addictive responses and cognitive impairments by alleviating neuroinflammation[J]. Journal of Ginseng Research,2023,47(4):583−592. doi: 10.1016/j.jgr.2023.02.003
[3] MILIVOJEVIC V, SULLIVAN L, TIBER J. Pregnenolone effects on provoked alcohol craving, anxiety, HPA axis, and autonomic arousal in individuals with alcohol use disorder[J]. Psychopharmacology (Berl),2023,240(1):101−114. doi: 10.1007/s00213-022-06278-3
[4] WANG S C, CHEN Y C, CHEN S J, et al. Alcohol addiction, gut mcrobiota, and alcoholism treatment:A review[J]. International Journal of Molecular Sciences,2020,21(17):6413. doi: 10.3390/ijms21176413
[5] ROSENBAUM J F. Anxiety treatment with benzodiazepines[J]. Focus (American Psychiatric Publishing) ,2021,19(2):211.
[6] PENG L, MORFORD K L, LEVANDER X A. Benzodiazepines and related sedatives[J]. The Medical Clinics of North America,2022,106(1):113−129. doi: 10.1016/j.mcna.2021.08.012
[7] NAGEEN B, SARFRAZ I, RASUL A, et al. Eupatilin:A natural pharmacologically active flavone compound with its wide range applications[J]. J Asian Nat Prod Res,2020,22(1):1−16. doi: 10.1080/10286020.2018.1492565
[8] 游新侠, 李越. 艾叶果蔬复合小馒头的研制[J]. 粮食与油脂, 2020, 33(10):75−79. [YOU Xinxia, LI Yue. The development of crunchy rice ball with mugwort and fruit-vegetable compound[J]. Cereals & Oils, 2020, 33(10):75−79. YOU Xinxia, LI Yue. The development of crunchy rice ball with mugwort and fruit-vegetable compound[J]. Cereals & Oils, 2020, 33(10): 75−79.
[9] 徐海卫. 艾叶黄酮异泽兰黄素对运动损伤大鼠炎症和抗氧化系统的影响[J]. 分子植物育种,2023,21(13):4414−4420. [XU Haiwei. Effects of Artemisia argyi flavonoid eupatilin on inflammation and antioxidant system in rats with sports injury[J]. Molecular Plant Breeding,2023,21(13):4414−4420. XU Haiwei . Effects of Artemisia argyi flavonoid eupatilin on inflammation and antioxidant system in rats with sports injury[J]. Molecular Plant Breeding,2023 ,21 (13 ):4414 −4420 .[10] 程德刚, 李福昌, 任刚. 异泽兰黄素通过DCST1-AS1/miR-138-5p对胰腺癌细胞SW-1990的影响[J]. 河北医药,2022,44(19):2907−2910,2915. [CHENG Degang, LI Fuchang, REN Gang. Effects and molecular mechanism of eupatilin yellow on the biological behavior ofpancreatic cancer cells-SW-1990 through DCST1-AS1/miR-138-5p[J]. Hebei Medical Journal, 2022, 44(19):2907−2910,2915. CHENG Degang, LI Fuchang, REN Gang. Effects and molecular mechanism of eupatilin yellow on the biological behavior ofpancreatic cancer cells-SW-1990 through DCST1-AS1/miR-138-5p[J]. Hebei Medical Journal, 2022, 44(19): 2907−2910,2915.
[11] ZHANG Y, QIN L, XIE J, et al. Eupatilin prevents behavioral deficits and dopaminergic neuron degeneration in a Parkinson's disease mouse model[J]. Life Sciences,2020,253:117745. doi: 10.1016/j.lfs.2020.117745
[12] JEONG H J, KIM J H, KIM N R, et al. Antidepressant effect of stillen[J]. Arch Pharm Res,2015,38(6):1223−1231. doi: 10.1007/s12272-014-0472-8
[13] SAKIMOTO Y, OO P M, GOSHIMA M, et al. Significance of GABAa receptor for cognitive function and hippocampal pathology[J]. International Journal of Molecular Sciences,2021,22(22):12456. doi: 10.3390/ijms222212456
[14] CENTANNI S W, TEPPEN T, RISHER M L, et al. Adolescent alcohol exposure alters GABA α receptor subunit expression in adult hippocampus[J]. Alcoholism, Clinical and Experimental Research,2014,38(11):2800−2808. doi: 10.1111/acer.12562
[15] ZHAO Z, KIM Y W, WU Y, et al. Korean Red Ginseng attenuates anxiety-like behavior during ethanol withdrawal in rats[J]. J Ginseng Res,2014,38(4):256−263. doi: 10.1016/j.jgr.2014.05.010
[16] LEE H Y, NAM Y, CHOI W S, et al. The hepato-protective effect of eupatilin on an alcoholic liver disease model of rats[J]. Korean J Physiol Pharmacol,2020,24(5):385−394. doi: 10.4196/kjpp.2020.24.5.385
[17] 王小龙, 武爽, 吴琦, 等. 芍药苷对酒精戒断大鼠焦虑样行为、炎症因子及肠道菌群的影响[J]. 食品工业科技,2022,43(10):378−387. [WANG Xiaolong, WU Shuang, WU Qi, et al. Effects of paeoniflorin on anxiety-like behavior, inflammatory factors and intestinal microflora in alcohol withdrawal rats[J]. Science and Technology of Food Industry,2022,43(10):378−387. WANG Xiaolong, WU Shuang, WU Qi, et al . Effects of paeoniflorin on anxiety-like behavior, inflammatory factors and intestinal microflora in alcohol withdrawal rats[J]. Science and Technology of Food Industry,2022 ,43 (10 ):378 −387 .[18] JIAO Y, KIM S C, WANG Y, et al. Sauchinone blocks ethanol withdrawal-induced anxiety but spares locomotor sensitization:involvement of Nitric Oxide in the bed nucleus of the stria terminalis[J]. Evid Based Complement Alternat Med,2021,2021:6670212.
[19] KHAZIPOV R, ZAYNUTDINOVA D, OGIEVETSKY E, et al. Atlas of the postnatal rat brain in stereotaxic coordinates[J]. Front Neuroanat,2015,9:161.
[20] 姜敏, 李露露, 吴桐, 等. 三白草酮通过调控海马Nrf2/HO-1表达减少酒精戒断大鼠抑郁样行为[J]. 中国病理生理杂志,2022,38(10):1803−1811. [JING Min, LI Lulu, WU Tong, et al. Sauchinone attenuates alcohol withdrawal-induced depression-like behaviors in rats by modulating hippocampal Nrf2/HO-1 expression[J]. Chinese Journal of Pathophysiology,2022,38(10):1803−1811. JING Min, LI Lulu, WU Tong, et al . Sauchinone attenuates alcohol withdrawal-induced depression-like behaviors in rats by modulating hippocampal Nrf2/HO-1 expression[J]. Chinese Journal of Pathophysiology,2022 ,38 (10 ):1803 −1811 .[21] 李光美, 王朝群, 唐超, 等. 天麻多糖对阿尔茨海默症小鼠焦虑行为及记忆功能的影响[J]. 遵义医科大学学报,2023,46(3):227−233. [LI Guangmei, WANG Chaoxian, TANG Chao, et al. Effect of gastrodia elata polysaccharide on anxiety-like and learning memory behaviors in AD mouse models[J]. Journal of Zunyi Medical University,2023,46(3):227−233. LI Guangmei, WANG Chaoxian, TANG Chao, et al . Effect of gastrodia elata polysaccharide on anxiety-like and learning memory behaviors in AD mouse models[J]. Journal of Zunyi Medical University,2023 ,46 (3 ):227 −233 .[22] FIGUEIREDO CERQUEIRA M M, CASTRO M M L, VIEIRA A A, et al. Comparative analysis between open field and elevated plus maze tests as a method for evaluating anxiety-like behavior in mice[J]. Heliyon,2023,9(4):e14522. doi: 10.1016/j.heliyon.2023.e14522
[23] 谢晓燕. 基于HPA轴功能调节探讨调神针法抗焦虑的临床及机制研究[D]. 广州:广州中医药大学, 2021. [XIE Xiaoyan. Discussion on the anti-anxiety effect of regulating spirit acupuncture technique based on the HPA axis neuromodulation:A clinical and mechanism research[D]. Guangzhou:Guangzhou University of Chinese Medicine, 2021. XIE Xiaoyan. Discussion on the anti-anxiety effect of regulating spirit acupuncture technique based on the HPA axis neuromodulation: A clinical and mechanism research[D]. Guangzhou: Guangzhou University of Chinese Medicine, 2021.
[24] 李晨浩, 杨行, 张勇. 酒精成瘾对中枢神经系统的影响[J]. 神经解剖学杂志,2021,37(5):601−604. [LI Chenhao, YANG Hang, ZHANG Yong. The effects of alcohol addiction on the central nervous system J]. Chinese Journal of Neuroanatomy,2021,37(5):601−604.
[25] VANAVESKI T, MPLCHANOVA S, PHAM D D, et al. PGC-1alpha signaling increases GABA(A) receptor subunit alpha2 expression, GABAergic neurotransmission and anxiety-like behavior in mice[J]. Front Mol Neurosci,2021,14:588230. doi: 10.3389/fnmol.2021.588230
[26] 李冬雪. 丙泊酚/七氟醚配伍对MCI大鼠GABAa受体α1/α2亚基蛋白稳态及抑郁状态的影响[D]. 天津:天津医科大学, 2020. [LI Dongxue. Effect of the combination of propofol and sevoflurane on GABAa receptor α1/α2 subunits protein proteostasis anddepressive state in MCI rats[D]. Tianjin:Tianjin Medical University, 2020. LI Dongxue. Effect of the combination of propofol and sevoflurane on GABAa receptor α1/α2 subunits protein proteostasis anddepressive state in MCI rats[D]. Tianjin: Tianjin Medical University, 2020.
[27] SINHA R, WEMM S, FOGELMAN N, et al. Moderation of Prazosin's efficacy by alcohol with drawal symptoms[J]. Am J Psychiatry,2021,178(5):447−458. doi: 10.1176/appi.ajp.2020.20050609
[28] LI Y, SHEN M, STOCKTON M E, et al. Hippocampal deficits in neurodevelopmental disorders[J]. Neurobiol Learn Mem,2019,165:106945. doi: 10.1016/j.nlm.2018.10.001
[29] ZAKHAROVA E I, PROSHIN A T, MONAKOV M Y, et al. Cholinergic internal and projection systems of hippocampusand neocortex critical for early spatial memory consolidation in normal and chronic cerebral hypoperfusion conditions in rats with different abilities to consolidation:The role of cholinergic interneurons of the hippocampus[J]. Biomedicines,2022,10,1532:1−28.
[30] BIANE J S, LADOW M A, STEFANINI F, et al. Neural dynamics underlying associative learning in the dorsal and ventral hippocampus[J]. Nature Neuroscience,2023,26(5):798−809. doi: 10.1038/s41593-023-01296-6
[31] TURNER V S, O'SULLIVAN R O, KHEIRBEKM A. Linking external stimuli with internal drives:A role for the ventral hippocampus[J]. Current Opinion in Neurobiology,2022,76:102590. doi: 10.1016/j.conb.2022.102590
[32] LEE S E, LEE Y, LEE G H. The regulation of glutamic acid decarboxylases in GABA neurotransmission in the brain[J]. Arch Pharm Res,2019,42(12):1031−1039. doi: 10.1007/s12272-019-01196-z
[33] KANAO-KANDA M, KANDA H, LIU S, et al. Viral vector-mediated gene transfer of glutamic acid decarboxylase for chronic pain treatment:A literature review[J]. Hum Gene Ther,2020,31(7-8):405−414. doi: 10.1089/hum.2019.359
[34] KIMOTO S, ZAKI M M, BAZMI H H, et al. Altered markers of cortical gamma-aminobutyric acid neuronal activity in schizophrenia:Role of the NARP gene[J]. JAMA Psychiatry,2015,72(8):747−756. doi: 10.1001/jamapsychiatry.2015.0533
[35] AGRAWAL J, DWIVEDI Y. GABA(A) receptor subunit transcriptional regulation, expression organization, and mediated calmodulin signaling in prefrontal cortex of rats showing testosterone-mediated impulsive behavior[J]. Front Neurosci,2020,14:600099. doi: 10.3389/fnins.2020.600099
[36] ENGIN E, BENHAM R S, RUDOLPH U. An emerging circuit pharmacology of GABA(A) receptors[J]. Trends Pharmacol Sci,2018,39(8):710−732. doi: 10.1016/j.tips.2018.04.003
[37] BERNARDO A, LEE P, MARCOTTE M, et al. Symptomatic and neurotrophic effects of GABAA receptor positive allosteric modulation in a mouse model of chronic stress[J]. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology,2022,47(9):1608−1619. doi: 10.1038/s41386-022-01360-y
[38] TIWARI V, Chopra K. Resveratrol abrogates alcohol-induced cognitive deficits by attenuating oxidative-nitrosative stress and inflammatory cascade in the adult rat brain[J]. Neurochem Int,2013,62(6):861−869. doi: 10.1016/j.neuint.2013.02.012
[39] SONG X, LONG D. Nrf2 and Ferroptosis:A new research direction for neurodegenerative diseases[J]. Front Neurosci,2020,14:267. doi: 10.3389/fnins.2020.00267
-
期刊类型引用(1)
1. 吴朋烊,赵容杰,李露露,李秋月,张春晶,吴丽娜,赵正林. 孤啡肽改善尼古丁戒断大鼠的焦虑样行为及其对HPA轴与炎症因子的调控机制. 中国病理生理杂志. 2024(10): 1926-1933 .  百度学术
百度学术
其他类型引用(1)
 下载:
下载:




计量
- 文章访问数: 167
- HTML全文浏览量: 24
- PDF下载量: 55
- 被引次数: 2



