


Effects of 1-MCP on the Mitochondrial Antioxidant Activity and Energy Metabolism of Zucchini Fruits
-
摘要: 为探究1-甲基环丙烯(1-methylcyclopropene,1-MCP)处理对西葫芦果实常温贮藏条件下线粒体活性氧代谢和能量代谢的影响,以‘泰山绿’西葫芦为试材,采用20 μL·L−1的1-MCP在室温下熏蒸24 h后将其置于常温(20±1)℃下贮藏12 d,贮藏期间每3 d测定西葫芦果实的生理品质、线粒体抗氧化特性和能量代谢变化。结果表明,贮藏结束时的第12 d,1-MCP处理西葫芦果实失重仅为对照的54.6%,总酚含量为对照的1.44倍;贮藏期间,果实的呼吸强度显著(P<0.05)低于同时期对照果实(>10 mg·kg−1·h−1),1-MCP处理还降低了西葫芦线粒体丙二醛和过氧化氢含量、抑制了超氧阴离子产生速率,在第6、9 d,处理果实的过氧化氢含量、超氧阴离子产生的速率分别比对照组低了17.45%、23.29%和10.51%、7.62%。1-MCP处理使西葫芦果实线粒体维持了较高的超氧化物歧化酶、抗坏血酸过氧化物酶活性,降低了脂氧合酶的活性,贮藏至第3 d和第6 d,处理组线粒体超氧化物歧化酶活性是对照组的1.23、1.15倍。此外1-MCP处理还延缓了琥珀酸脱氢酶、细胞色素C氧化酶、线粒体H+-ATPase、线粒体Ca2+-ATPase活性的下降。贮藏第6 d,1-MCP处理的西葫芦果实后四种酶的活性都显著高于对照(P<0.05),H+-ATPase、Ca2+-ATPase分别是对照的1.05和1.22倍。因此得出结论,1-MCP可以通过减轻西葫芦果实线粒体膜氧化损伤,保持线粒体的完整性,延缓成熟衰老进程,有利于延长西葫芦果实贮藏期。Abstract: In order to explore the effects of 1-methylcyclopropene (1-MCP) treatment on mitochondrial reactive oxygen metabolism and energy metabolism of zucchini fruits stored at room temperature, this study took 'Taishan Green' zucchini as test materials. The fruits were fumigated with 20 μL·L−1 1-MCP at room temperature for 24 h and then stored at room temperature (20±1) ℃ for 12 d. The physiological quality, mitochondrial antioxidant properties and energy metabolism change of zucchini fruits were determined every 3 d during storage. Results showed that: On the 12th day at the end of storage, the weight loss of 1-MCP treated zucchini fruit was only 54.6% of that of the control, and the total phenol content was 1.44 times of the control. During storage, the respiration intensity of fruits was significantly lower than that of control fruits (>10 mg·kg−1·h−1) (P<0.05), 1-MCP treatment also decreased the content of malondialdehyde and hydrogen peroxide in mitochondria of zucchini and inhibited the production rate of superoxide anion. The hydrogen peroxide content and superoxide anion production rate of the treated fruits were 17.45% and 23.29% lower and 10.51% and 7.62% lower than those of the control group, respectively. 1-MCP treatment kept the activities of superoxide dismutase and ascorbate peroxidase higher in mitochondria of zucchini fruits, decreased the activities of lipoxygenase. The mitochondrial superoxide dismutase activity in the treatment group was 1.23 and 1.15 times of that in the control group at the 3rd and 6th day of storage. In addition, 1-MCP treatment also delayed the decrease of the activities of succinate dehydrogenase, cytochrome C oxidase, mitochondrial H+-ATPase and mitochondrial Ca2+-ATPase. On the sixth day of storage, the activities of four enzymes of zucchini fruits treated with 1-MCP were significantly higher than those of the control (P<0.05), the activities of H+-ATPase and Ca2+-ATPase were 1.05 and 1.22 times than those of the control. Therefore, 1-MCP could reduce the oxidative damage of mitochondrial membrane of zucchini fruits, maintain the integrity of mitochondria, delay the process of maturation and senescence, and prolong the storage period of zucchini fruits.
-
Keywords:
- zucchini /
- 1-MCP /
- mitochondria /
- reactive oxygen species /
- antioxidant activity /
- energy
-
西葫芦(Cucurbita pepo L.)为葫芦科南瓜属植物,是葫芦科南瓜属西葫芦种[1]。西葫芦果实质地脆嫩,富含多种维生素、膳食纤维等物质。西葫芦果实采后生理代谢旺盛,表皮常出现褪色、组织变松软,甚至腐败变质,失去商品价值。
目前保持西葫芦果实品质的物理方法有低温、热空气、热水、高相对湿度等,化学方法有水杨酸、1-MCP、壳聚糖涂膜等,也有通过复合的方式进行处理,如冷激结合甜菜碱[2]。西葫芦的保鲜方法有许多,然而,在采后贮藏过程中,这些方法通常由于成本高、不环保等因素难以应用到实际生产中。1-MCP作为一种绿色、无污染、成本不高的化学保鲜剂,已经广泛应用到生产实践当中,1-MCP是一种新型的乙烯抑制剂,具有与乙烯相似的性质,能够吸引乙烯受体中金属离子的电子并配对,与乙烯受体牢固结合从而达到抑制内外源乙烯的合成作用,降低果蔬采后呼吸和乙烯强度,延缓其成熟和衰老进程,保持商品品质[3]。
活性氧(reactive oxygen species,ROS)是生物体在正常有氧呼吸过程中产生的一类代谢产物,主要包括过氧化氢(H2O2)、超氧阴离子(O2−·)、羟自由基(-OH)等,ROS与果蔬的成熟衰老有关,线粒体是ROS产生的主要来源,ROS也将线粒体作为靶部位[4]。机体内过多的ROS会产生不利的影响,如破坏线粒体膜内氧化还原平衡,而线粒体内同时也存在着一些抗氧化物酶对ROS起清除作用[4]。如超氧化物歧化酶(superoxide dismutase,SOD)、过氧化氢酶(catalase,CAT)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)。大量研究结果表明,1-MCP可以通过提高抗氧化物酶的活性或抗氧化物质的含量来清除ROS达到延长保质期的作用[5-6]。Song等[7]研究发现,小白菜叶绿体中ROS积累会引起叶绿体细胞膜受损,而1-MCP可能通过提高抗氧化物酶SOD、APX的活性从而延缓了叶片衰老。线粒体是有氧呼吸的主要场所,呼吸过程会产生大量能量(ATP),而ATP的生成与能量代谢相关酶密不可分,琥珀酸脱氢酶(succinate dehydrogenase,SDH)、细胞色素C氧化酶(cytochrome C oxidase,CCO)、H+-ATPase和Ca2+-ATPase是能量代谢水平的特征酶[8]。目前,已有研究表明‘金红’苹果采后果皮组织褐变与能量亏缺有密切关系[9],张正敏等[10]研究‘白凤’桃的桃腐病发现较高的能荷水平能抑制桃果实发病率的上升。综上,目前研究1-MCP延缓果实成熟衰老机理的系统较完备,但在更深层次方面研究1-MCP对西葫芦果实内部线粒体的调控机理尚未明确,本文通过对1-MCP处理在常温贮藏下西葫芦果实品质和生理生化的研究,探究1-MCP处理对西葫芦果实线粒体抗氧化能力和能量代谢水平的调控机制,为1-MCP应用于采后西葫芦贮藏保鲜技术提供理论支撑。
1. 材料与方法
1.1 材料与仪器
西葫芦 江苏康润农业科技发展有限公司提供,品种为‘泰山绿’,露地栽培。选取大小均匀、无机械损伤、且与实验所需成熟度相符(成熟期)的西葫芦;1-MCP 咸阳西秦生物科技有限公司;三氯乙酸(Trichloroacetic acid,TCA)、四氯化钛、α-萘胺、亚油酸钠、没食子酸、聚乙烯吡咯烷酮(Polyvinylpyrrolidone,PVP) 上海麦克林生物化学有限公司;乙酸钠、30% H2O2(分析纯)、愈创木酚(含量≥99%,化学纯) 国药集团化学试剂有限公司;硫酸、盐酸 南京化学试剂股份有限公司;盐酸羟胺、乙二胺四乙酸二钠 西陇化工股份有限公司;琥珀酸钠、5-甲基酚嗪硫酸甲酯(Phenazine methosulfate,PMS)、细胞色素C(含量≥95%,牛心)、牛血清白蛋白、N,N-二甲基对苯二胺 上海源叶生物科技有限公司;三羟甲基氨基甲烷(Trimethylol aminomethane,Tris)、考马斯亮蓝G-250、甘露醇 北京索莱宝科技有限公司。
CR-400 CHROMA METER色差仪 日本柯尼卡美能达有限公司;PAL-1G电子数显折光仪 ATAGO公司;Alpha-1860A紫外可见分光光度计 上海谱元仪器有限公司;H1750R台式高速冷冻离心机 湖南湘仪实验室仪器开发有限公司;HH-6数显恒温水浴锅 常州国华电器有限公司;A11分析用研磨机 德国IKA;FE30实验室电导率仪、FE20 Plus实验室pH计 梅特勒-托利多仪器有限公司。
1.2 实验方法
1.2.1 材料及处理方法
挑选600个西葫芦果实,随机分为两组置泡沫箱,分别进行如下处理:a.采用20 μL·L−1的1-MCP熏蒸处理24 h,打开泡沫箱通风1 h,将上述西葫芦果实套塑料袋装框后置于常温(20±1 ℃)、相对湿度90%左右的环境中贮藏12 d;b.对照组(CK):不用1-MCP处理但使用同样操作。取样点为−1(样品当天运回未做任何处理之前,作为初始参考)、0、3、6、9、12 d。每个采样点取30个果实。在每个采样时间点测定果实品质和生理指标后,将西葫芦果实去皮、去籽,果肉切块,立即放到液氮中速冻,并保存在−80 ℃冰箱中以供进一步分析。试验设置三个重复样品。
1.2.2 失重率、总可溶性固形物的测定
果实失重率测定方法为取10个1-MCP处理过的西葫芦果实,称取每个贮藏时间点果实的重量,对照处理保持一致,重量记为M1,初始重量记为M0,单位以g计。计算公式为:
失重率(%)=M0−M1M0×100 总可溶性固形物(TSS)测定方法为取新鲜西葫芦果肉组织榨汁,使用ATAGO手持式折射仪测定,测定结果以%表示。
1.2.3 可溶性蛋白、总酚、VC含量的测定
可溶性蛋白含量的测定参考曹建康等[11]的方法略有改动。将新鲜西葫芦果肉切碎并混合均匀,称取1 g样品,加入5 mL蒸馏水,涡旋混匀后在4 ℃、12000 g离心20 min,收集上清液4 ℃低温保存。吸取1 mL上清(视蛋白质含量适当稀释,计算时按照稀释倍数计算即可),加入5 mL考马斯亮蓝G-250溶液,混合均匀,2 min后在波长595 nm处比色,标准曲线的制作方法为取六支试管依次按比例加入100 μg/mL的标准蛋白质溶液和蒸馏水,共计1 mL,混匀后按照样品测定方法进行测定,重复测定三次。
总酚含量参考Qiu等[12]的方法略有改动。称取2 g样品,加入4 mL 1% HCl-乙醇溶液,4 ℃、12000 g离心20 min,收集上清液4 ℃低温保存。取0.1 mL上清加入0.4 mL蒸馏水、2.5 mL福林酚、2 mL 7.5%碳酸钠溶液,45 ℃水浴15 min,于765 nm下测定吸光值,重复测定三次。
VC含量的测定参考曹建康等[11]的方法,采用分光光度法,含量表示为mg·100 g−1,重复测定三次。
1.2.4 线粒体的提取
线粒体的提取参考Millar等[13]的方法略有改动,称取6 g样品,根据绿色植物组织的线粒体提取方法,按比例加入低温预冷过的pH7.5 50 mmol·L−1 Tris-HCl提取液(含1 mmol·L−1 EDTA、0.25 mol·L−1蔗糖、0.3 mol·L−1甘露醇、0.1%半胱氨酸、0.1%BSA、0.5%PVP),涡旋混合均匀后经四层纱布过滤,使用玻璃棒搅拌收集滤液于50 mL离心管中,于4 ℃ 6000 g离心5 min,弃去沉淀,取上清采用同样的步骤再离心一次。离心后得到的滤液以14000 g在4 ℃离心20 min,弃上清,收集沉淀,用20 mL pH7.2 10 mmol·L−1 Tris-HCl洗涤液(含1 mmol·L−1 EDTA、0.25 mol·L−1蔗糖、0.3 mol·L−1甘露醇、0.1%BSA)洗涤,于4 ℃ 14000 g离心20 min,洗涤过程重复三次,得到沉淀即为线粒体。将线粒体沉淀保存在1.5 mL pH7.2 10 mmol·L−1 Tris-HCl悬浮液(含1 mmol·L−1 EDTA、0.25 mol·L−1蔗糖、0.3 mol·L−1甘露醇)中,用于线粒体相关酶活性的测定,线粒体制备需现提现用,否则会破坏线粒体完整性,使实验结果不准确。整个操作过程在冰浴条件下进行。
1.2.5 呼吸强度、线粒体MDA、H2O2、O2−·含量的测定
1.2.5.1 呼吸强度的测定
每个处理取1.2 kg左右(约3个)西葫芦果实,放入密封的玻璃罐中1 h,采用CO2呼吸测定仪测呼吸速率,结果表示为CO2 mg·kg−1·h−1。计算公式如下:
呼吸强度 (CO2mg⋅kg−1⋅h−1)=1.96×1000×V×A%t×m 式中:1.96×103表示CO2密度,mg/L;V表示玻璃罐体积,L;A%为CO2测定仪数值;t为测定时间,h;m为样品重量,kg。
1.2.5.2 线粒体MDA含量的测定
MDA含量的测定参考Li等[14]的方法略有改动。将以“1.2.4”方法制备的线粒体悬浮液涡旋搅拌30 s,使线粒体悬浮液混合均匀,取1 mL线粒体悬浮液加入2 mL 0.67% TBA,混匀后于置于水浴锅中100 ℃煮沸30 min,待冷却后4 ℃、12000 g下离心5 min,弃去沉淀,取线粒体上清液分别测定其在波长532nm和600 nm波长处的吸光值。
MDA( μmol⋅g-1FW)=(A532−A600)×V0.155×Vs×m 式中:V表示提取样品液体积,mL;Vs表示测定液体积,mL;m表示样品质量,g。
1.2.5.3 线粒体H2O2含量的测定
H2O2含量的测定参考曹健康等[11]的方法略有改动。取1 mL线粒体悬浮液加入0.1 mL 20%硫酸钛和0.2 mL浓氨水,混匀后25 ℃保温10 min后加入2 mol·L−1的硫酸3 mL,摇动使沉淀完全溶解,取上清液在测定其在波长415 nm处的吸光值。按照同样的方法制作H2O2标准曲线,最终的H2O2含量表示为μmol·g−1 FW。
H2O2(μmol⋅g−1FW)=n×VVs×m 式中:n表示标准曲线查得样品中H2O2浓度,μmol;V表示提取样品液体积,mL;Vs表示测定液体积,mL;m表示样品质量,g。
1.2.5.4 线粒体O2−.产生速率的测定
O2−.含量的测定参考贾乐等[15]的方法略有改动。取1 mL线粒体悬浮液加入1 mL磷酸钠缓冲液(pH7.8,50 mmol/L)和1 mL 1 mmol·L−1盐酸羟胺溶液,混合均匀后在25 ℃下保温20 min。取出后依次加入1 mL 17 mmol·L−1对氨基苯磺酸和1 mL mmol·L−1 α-萘胺溶液,在25 ℃下再保温30 min后进行显色反应,测定在波长530 nm处的吸光值。最终的O2−.含量表示为nmol·min−1·g−1 FW。
O−2⋅[nmol⋅(g⋅min)-1FW]=n×VVs×t×m 式中:n表示标准曲线查得样品中O2−.物质的量,nmol;V表示提取样品液体积,mL;Vs表示测定液体积,mL;m表示样品质量,g;t表示反应时间,min。
1.2.6 线粒体SOD、APX、LOX活性的测定
1.2.6.1 线粒体SOD活性的测定
SOD活性测定的方法参考Zheng等[16]的方法略有改动。取7支规格一致的试管,测定管、光下对照管各三支,暗中对照管1支,每管按照顺序加入试剂,依次加入50 mmol·L−1磷酸缓冲液1.7 mL,130 mmol·L−1甲硫氨酸溶液0.3 mL,1 mmol·L−1 EDTA溶液0.3 mL,130 mmol·L−1氮蓝四唑溶液0.3 mL,1 mmol·L−1核黄素溶液0.3 mL,最后加入0.1 mL线粒体悬浮液,混匀后立刻将6支试管置于4000 lx灯下显色反应30 min,反应结束后立即用黑布遮盖终止反应。以暗中对照管作为空白参比调零,测定其它管在560 nm处的吸光度值。SOD活性以每分钟每克样品反应体系对NBT光化还原的抑制为50%为1个酶活力单位,结果用U·g−1 FW表示。
1.2.6.2 线粒体APX活性的测定
APX活性测定方法参考Sharma等[17]的方法略有改动,反应体系包含APX反应液2.5 mL、0.2 mL线粒体悬浮液和0.3 ml 2 mmol·L−1 H2O2溶液。立即测定在290 nm处的吸光值,每30 s记录一次数值,记录3 min内的吸光值变化。APX的活性以每分钟每克样品吸光度变化0.01为1个酶活力单位,结果用U·g−1 FW表示。
1.2.6.3 线粒体LOX活性的测定
LOX活性测定方法参考Mao等[18]的方法略有改动。3 mL反应体系中包含2.75 mL乙酸钠缓冲液(pH5.6,0.05 mol·L−1),0.15 mL亚油酸钠底物和0.1 mL线粒体悬浮液。在234 nm处连续反应3 min,每30 s计数一次,记录3 min内的吸光值变化。LOX的活性以每分钟每克样品吸光度变化0.01为一个酶活力单位,结果用U·g−1 FW表示。
1.2.7 线粒体SDH、CCO、H+-ATPase和Ca2+-ATPase活性的测定
1.2.7.1 线粒体SDH活性的测定
SDH活性的测定参考Ackrell等[19]的方法稍作修改。取0.2 mL线粒体悬浮液,反应体系包括2.6 mL磷酸钾缓冲液(pH7.4,0.2 mol·L−1)、0.1 mL 0.9 mmol·L−1 2,6-二氯酚靛酚钠(DCPIP)。混匀后在30 ℃条件下孵育5 min,测定前加0.1 mL 0.33% PMS混匀后,测定DCPIP在600 nm处的变化。
1.2.7.2 线粒体CCO活性的测定
CCO活性测定参照Errede等[20]的方法并稍作修改。反应体系包括(2 mL蒸馏水、0.2 mL质量分数为0.04%的细胞色素C、0.2 mL线粒体悬浮液),将混合液在经过37 ℃、2 min的加热之后,再加入0.4% N,N-二甲基对苯二胺溶液0.5 mL,混合均匀后再保温3 min,在波长510 nm处测定吸光值的变化。CCO活性以每分钟每克样品吸光度变化0.01为一个酶活力单位,结果用U·g−1 FW表示。
1.2.7.3 线粒体H+-ATPase和Ca2+-ATPase活性的测定
H+-ATPase和Ca2+-ATPase的活性测定参考Jin等[21]的方法。H+-ATPase反应体系中含有0.7 mL pH8.0 30 mmol·L−1 Tris-HCl缓冲液(含3 mmol·L−1 MgSO4、50 mmol/LNaNO3、0.1 mmol·L−1 Na3VO4、50 mmol·L−1 KCl、0.1 mmol·L−1钼酸铵)、0.1 mL线粒体悬浮液,待混匀后加入0.1 mL ATP-Tris(pH7.5,30 mmol·L−1)迅速启动反应,反应在37 ℃保温20 min,最后加入0.2 mL 55% TCA终止反应。最终结果用无机磷含量表示,以每克样品每小时释放1 μmol无机磷所需要的酶量为1个酶活力单位。Ca2+-ATPase活性测定方法类同H+-ATPase。
1.3 数据处理
所有数据为三个生物学重复样品的平均值。采用Excel 2021和SPSS 25.0进行数据的分析处理,并将P<0.05定义为具有显著性差异,采用Origin 2021软件进行作图。
2. 结果与分析
2.1 1-MCP处理对西葫芦果实常温贮藏中失重率、TSS、可溶性蛋白、总酚、VC含量的影响
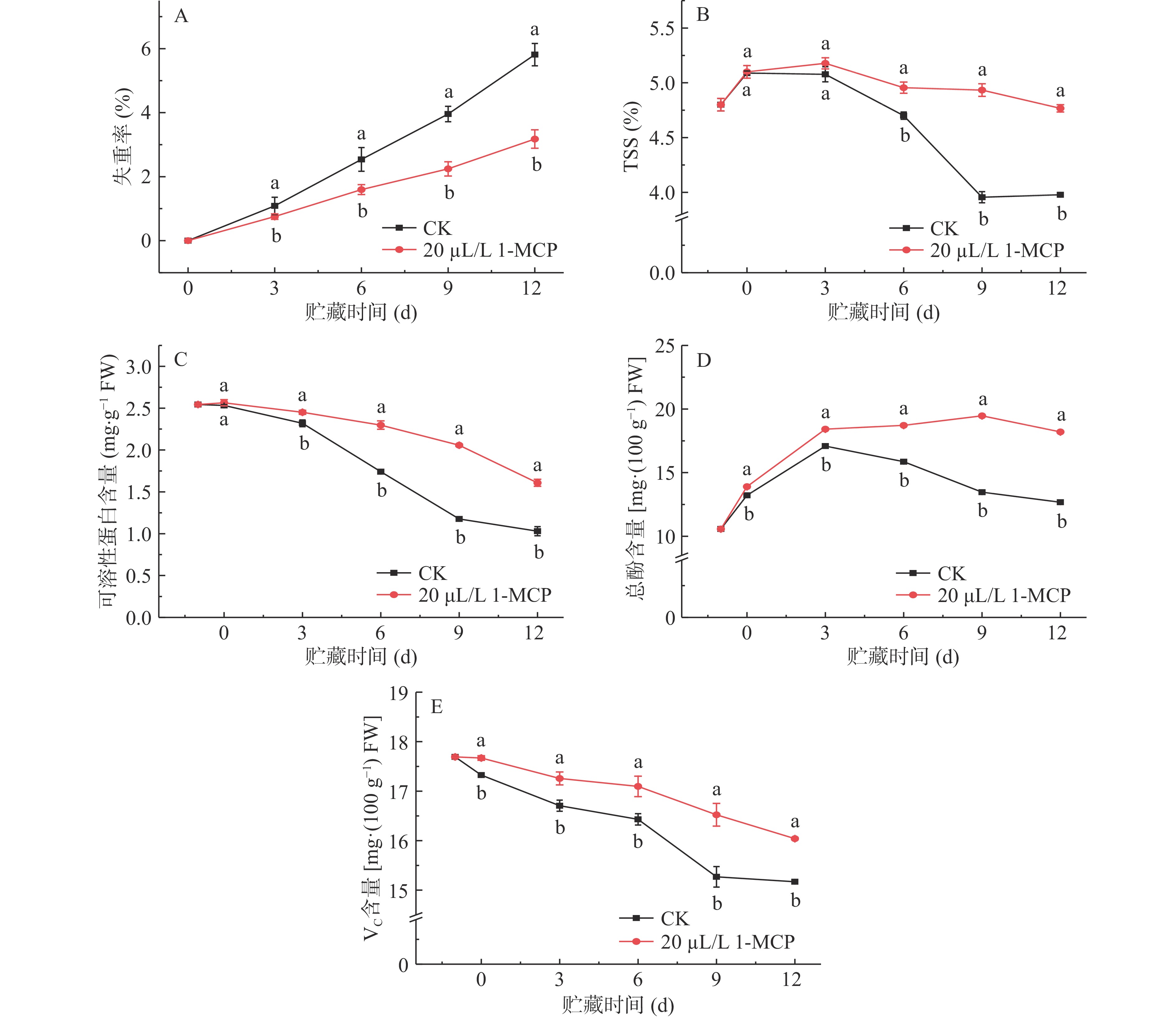
1-MCP处理对西葫芦果实失重率、TSS、可溶性蛋白、总酚和VC含量变化的影响见图1。图1A显示,西葫芦果实的失重率随着贮藏时间持续上升,但1-MCP处理可显著减少果实的失重(P<0.05)。贮藏到第12 d时,对照西葫芦果实的失重率为5.81%,处理仅为3.18%,对照组失重率是1-MCP处理组的1.83倍。因此,1-MCP处理对于降低西葫芦果实的失重有效。
![]()
果实中的TSS可以在一定程度上反应果实在贮藏期间品质和成熟度的变化趋势。由图1B可知,西葫芦果实TSS并不表现出持续下降的现象,而是呈现先上升后下降的趋势,1-MCP处理果实在第3 d后与对照TSS质量分数产生显著差异(P<0.05),第9 d和第12 d 处理组TSS分别比对照组高了19.8%、16.5%,因此,1-MCP处理可延缓TSS的下降,保持果实品质。
图1C表明,对照组西葫芦果实可溶性蛋白含量在0~3 d内缓慢下降,在3 d后迅速下降,而1-MCP处理的果实可溶性蛋白含量下降趋势较平缓,且在3~12 d内的同一贮藏期内,处理组含量显著高于对照组(P<0.05),在第3 d,对照可溶性蛋白含量2.32 mg·g−1,在12 d为1.03 mg·g−1,期间下降了55.58%,而处理可溶性蛋白含量仅下降了34.39%,因此,1-MCP处理抑制了西葫芦果实可溶性蛋白的降解过程。
果蔬中酚类物质的含量可以作为衡量抗氧化活性的指标之一。图1D表明,西葫芦果实在贮藏期间总酚含量呈先上升后下降的趋势。在贮藏第3 d和第9 d,对照组和处理组的总酚含量分别达到顶峰17.09、19.47 mg·100 g−1,而后开始下降,但在下降期间1-MCP处理的果实总酚含量依然显著高于对照(P<0.05),在贮藏12 d时处理组总酚含量是对照的1.44倍,故1-MCP处理延缓了西葫芦果实总酚含量的下降。
VC作为重要的抗氧化物质,在防止果实氧化方面起重要作用。如图1E所示,VC含量随着贮藏时间的延长不断下降,且在贮藏期内20 μL·L−1 1-MCP处理过的西葫芦果实与对照组变化趋势相同,在贮藏初期0~6 d缓慢下降,6~12 d下降速度明显加快,在贮藏期间,1-MCP处理的西葫芦果实VC含量显著高于对照组西葫芦,在贮藏的第9 d,处理果实VC含量分别为16.52 mg·100 g−1,比处理组高1.25 mg·100 g−1。因此,1-MCP处理减少了西葫芦果实中VC的氧化。
2.2 1-MCP处理对西葫芦果实常温贮藏中呼吸强度、线粒体MDA、H2O2含量、O2−·产生速率的影响
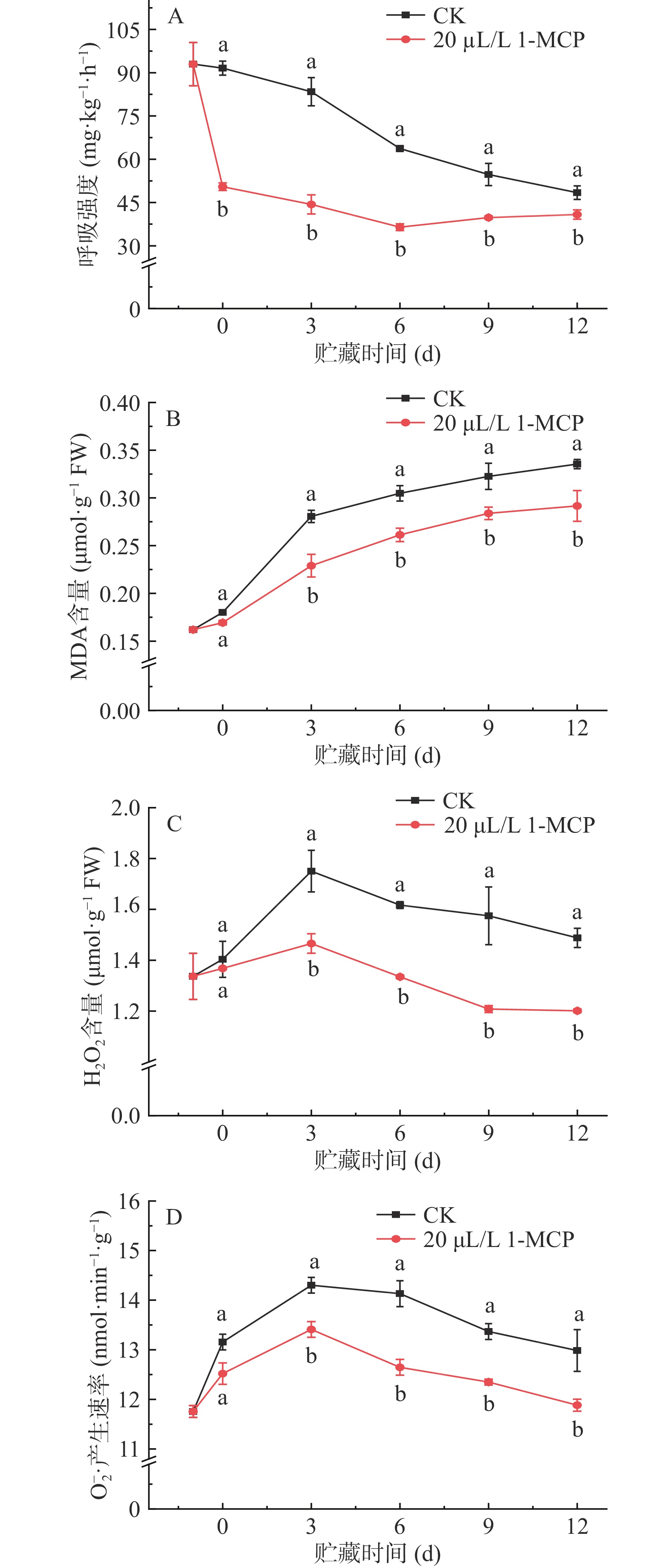
呼吸是新陈代谢的主导过程,是采后最主要的生理活动,也是生命存在的重要标志。贮藏期间1-MCP处理对西葫芦果实呼吸影响见图2A。由图2A可知,贮藏期间西葫芦果实的呼吸强度呈下降趋势,且经过1-MCP处理的西葫芦果实其呼吸强度显著低于对照果实(P<0.05),且在每个取样点对照组西葫芦的呼吸强度均比处理组强10 mg·kg−1·h−1以上。如第3 d,处理组与对照组西葫芦的呼吸强度分别为44.34、83.41 mg·kg−1·h−1,前者仅为后者的53.2%。这一结果表明,1-MCP对西葫芦果实的呼吸作用影响巨大,抑制呼吸有利于减少呼吸消耗,进而有利于果实品质的保持。
![]() 图 2 1-MCP处理对西葫芦果实呼吸强度(A)、线粒体MDA(B)、H2O2含量(C)、O2−·产生速率变化(D)的影响Figure 2. Effects of 1-MCP treatment on respiration intensity (A), mitochondrial MDA (B), H2O2 content (C) and O2−· (D) production rate of zucchini fruits
图 2 1-MCP处理对西葫芦果实呼吸强度(A)、线粒体MDA(B)、H2O2含量(C)、O2−·产生速率变化(D)的影响Figure 2. Effects of 1-MCP treatment on respiration intensity (A), mitochondrial MDA (B), H2O2 content (C) and O2−· (D) production rate of zucchini fruits线粒体MDA积累是产生线粒体膜脂过氧化的主要原因,积累过多会影响果实生理代谢。1-MCP对西葫芦果实线粒体MDA的影响如图2B。由图2B可知,贮藏期间西葫芦果实线粒体MDA含量呈持续上升趋势,在0~3 d内上升速度较快,在贮藏3~12 d内对照组果实的线粒体MDA含量显著高于处理组(P<0.05),如第6、9、12 d对照组的线粒体MDA含量分别是处理组的1.17、1.14、1.15倍,说明1-MCP处理能显著抑制西葫芦果实的线粒体MDA含量,减少膜脂过氧化程度,降低线粒体膜结构损伤。
H2O2和O2−.是衡量ROS的重要指标,由图2C、图2D可见,在同一贮藏期内,线粒体H2O2和O2−.产生速率的变化趋势都表现为先上升后下降,进一步研究发现,从第3 d开始,1-MCP处理的西葫芦果实显著抑制了线粒体内H2O2和O2−.含量的升高,在贮藏第9 d,处理组线粒体H2O2的含量为1.21 μmol∙g−1,比对照组低23.29%。对于O2−.来说,处理组O2−.的产生速率分别为12.65 nmol·g−1∙min−1,低于对照西葫芦果实10.51%。因此,1-MCP处理可以显著抑制西葫芦果实线粒体H2O2含量和O2−.产生速率,减少了果实组织内ROS含量,对保持果蔬品质具有重要作用。
2.3 1-MCP处理对西葫芦果实常温贮藏中线粒体SOD、APX、LOX活性的影响
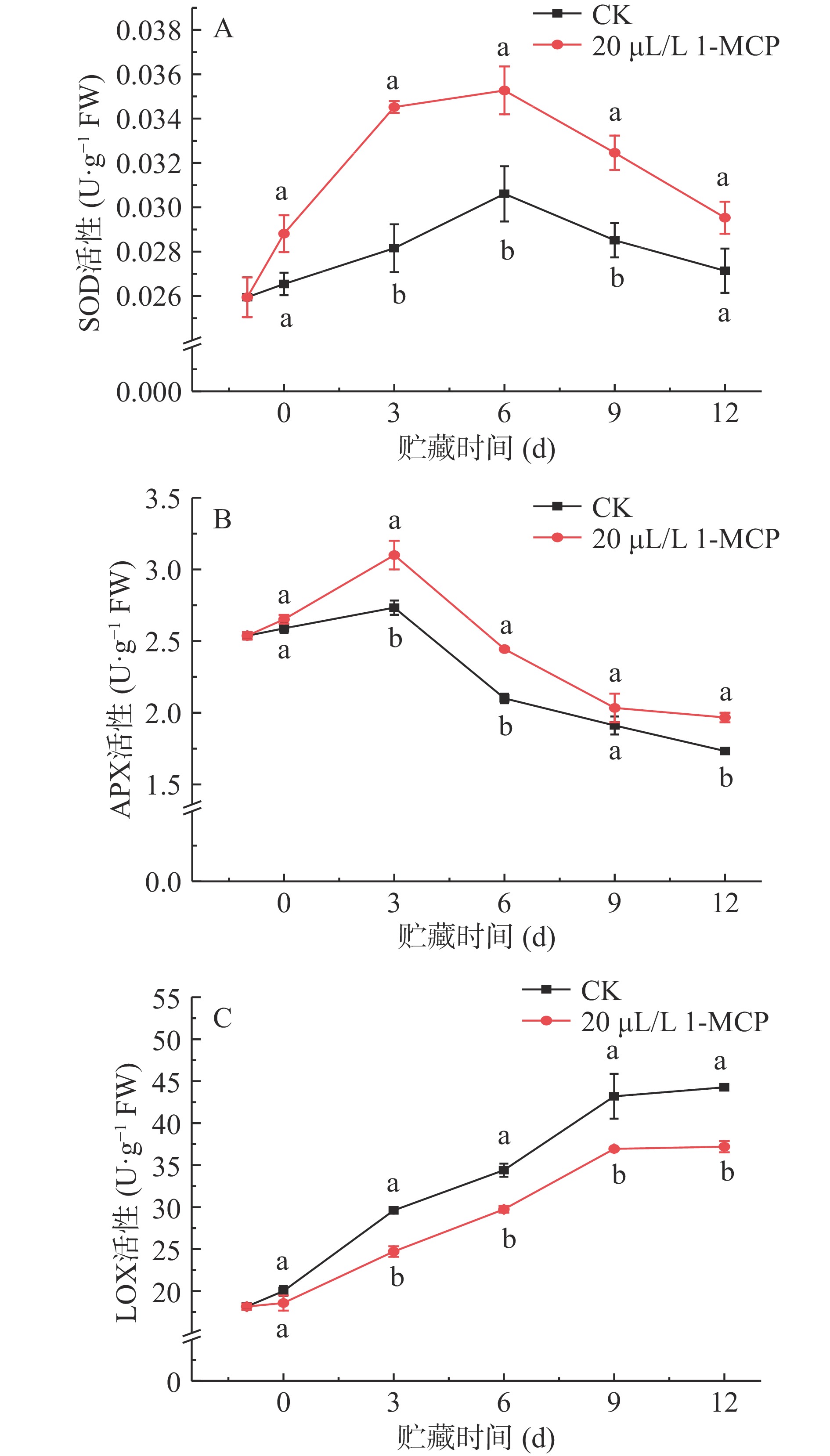
SOD、APX是抗氧化物酶,是清除ROS自由基的主要酶,在保持ROS代谢平衡中发挥着重要作用。如图3A所示,西葫芦果实线粒体SOD活性呈现先上升后下降的趋势,在贮藏中期(3~9 d),1-MCP处理的西葫芦果实线粒体SOD活性显著高于对照组(P<0.05),而在前期和末期没有显著差异(P<0.05),推测可能SOD在贮藏中期发挥的清除ROS的能力较大。贮藏至第3 d和第6 d,处理组线粒体SOD活性是对照组的1.23、1.15倍。1-MCP处理对西葫芦果实线粒体APX的影响如图3B所示,APX活性变化趋势表现为先上升后下降,且处理组和对照组一致,在第3 d达到高峰,在贮藏中期(3~6 d)和末期第12d处理组与对照组呈现显著性差异(P<0.05),第3 d、第6 d的处理组APX活性分别为3.1、2.44 U·g−1,比对照高了0.37、0.34 U·g−1。因此,1-MCP处理对采后西葫芦果实线粒体SOD、APX活性的下降有延缓作用,增强系统抗氧化能力,减少氧化损伤。
![]() 图 3 1-MCP对西葫芦果实线粒体SOD(A)、APX(B)和LOX(C)活性的影响Figure 3. Effects of 1-MCP on the SOD (A), APX (B) and LOX (C) activities in mitochondria of zucchini fruits
图 3 1-MCP对西葫芦果实线粒体SOD(A)、APX(B)和LOX(C)活性的影响Figure 3. Effects of 1-MCP on the SOD (A), APX (B) and LOX (C) activities in mitochondria of zucchini fruits图3C表明,西葫芦果实线粒体LOX活性在贮藏期间持续上升,在第9 d和第12 d,对照组果实线粒体LOX活性为43.2、44.27 U·g−1,较第3 d上升了13.6、14.67 U·g−1。1-MCP处理的西葫芦果实线粒体LOX活性在贮藏期间始终低于对照组,且从第3 d开始,对照组线粒体LOX活性与处理组出现显著性差异(P<0.05)。因此,1-MCP处理抑制了西葫芦果实线粒体LOX活性的上升。
2.4 1-MCP处理对西葫芦果实常温贮藏中SDH、CCO、H+-ATPase和Ca2+-ATPase活性的影响
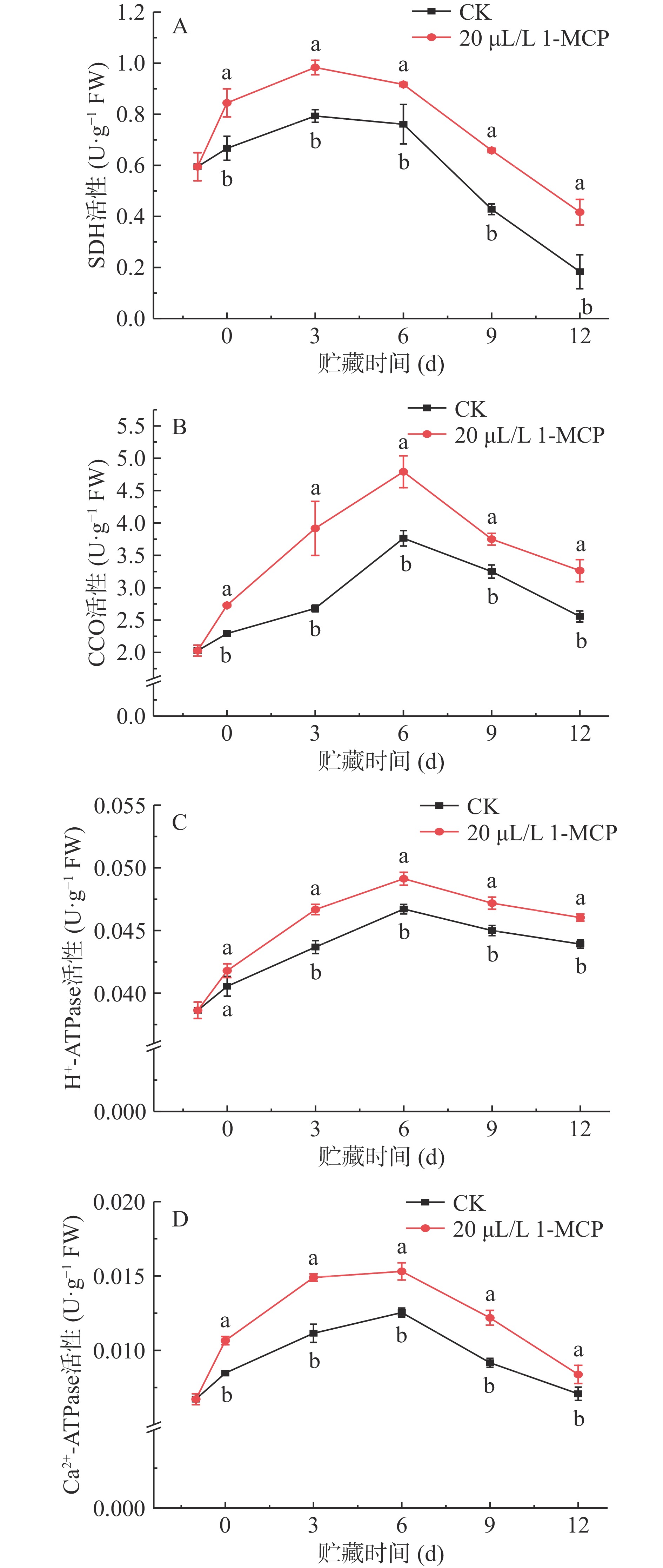
SDH是线粒体能量代谢的关键酶,是三羧酸循环(TCA)的关键酶。如图4A所示,贮藏期间SDH活性呈波动状态,且1-MCP处理西葫芦果实的SDH活性始终显著高于同时间的对照(P<0.05)。SDH活性在第3 d达到最高峰,处理组和对照组分别为0.98、0.79 U·g−1,是贮藏末期(12 d)活性的2.36和4.33倍。CCO是电子传递链末端的氧化酶,也是能量代谢中的关键酶。如图4B所示,其变化趋势与SDH相似,其中对照组果实CCO活性下降较明显,值得注意的是,处理组果实CCO活性在贮藏期间都显著高于对照组(P<0.05)。如在贮藏第6 d,对照组CCO活性为3.76 U·g−1,仅为处理组的21.45%。1-MCP处理能够保持西葫芦果实SDH、CCO活性在贮藏期间处于较高水平。
![]() 图 4 1-MCP处理对西葫芦线粒体SDH(A)、CCO(B)、H+-ATPase(C)和Ca2+-ATPase(D)活性的影响Figure 4. Effects of 1-MCP on SDH (A), CCO (B), H+-ATPase (C) and Ca2+-ATPase (D) activity of zucchini fruits
图 4 1-MCP处理对西葫芦线粒体SDH(A)、CCO(B)、H+-ATPase(C)和Ca2+-ATPase(D)活性的影响Figure 4. Effects of 1-MCP on SDH (A), CCO (B), H+-ATPase (C) and Ca2+-ATPase (D) activity of zucchini fruits1-MCP处理对西葫芦果实的H+-ATPase和Ca2+-ATPase活性影响如图4C、图4D所示,在贮藏期间活性变化趋势呈波动状态,这与前面提到的SDH与CCO活性的变化趋势是保持一致的,且与CCO活性变化趋势相同,在贮藏的第6 d达到最高点。进一步研究发现,第6 d处理组果实的H+-ATPase和Ca2+-ATPase分别是对照组的1.05和1.22倍。H+-ATPase在贮藏期第3 d到贮藏末期(12 d),对照组活性变化显著低于处理组(P<0.05),而对照组Ca2+-ATPase活性在整个贮藏期内都显著低于处理组(P<0.05)。
2.5 相关性分析
如表1相关性数据分析可知,采后西葫芦果实的呼吸强度与总酚呈显著性负相关(P<0.05),与SOD呈极显著性负相关(P<0.01);TSS与可溶性蛋白、VC、总酚、SOD、APX、SDH都呈现显著性正相关(P<0.05),与线粒体MDA、LOX呈极显著性负相关(P<0.01),VC与MDA、LOX呈极显著性负相关(P<0.01);MDA与LOX呈极显著正相关(P<0.01),与APX、SDH呈显著负相关(P<0.05);实验结果表明,1-MCP处理西葫芦果实采后品质与线粒体抗氧化和能量代谢的关系极为密切。
表 1 1-MCP处理西葫芦果实采后各项生理指标的相关性Table 1. Correlation of physiological indexes of postharvest zucchini fruits treated by 1-MCP项目 TSS 可溶性蛋白 VC 总酚 MDA SOD APX LOX SDH 呼吸强度 0.233 0.316 0.268 −0.398* −0.352 −0.524** 0.339 −0.383* −0.188 TSS 0.944** 0.803** 0.519** −0.702** 0.383* 0.709** −0.831** 0.670** 可溶性蛋白 0.924** 0.326 −0.837** 0.312 0.797** −0.916** 0.684** VC 0.066 −0.954** 0.158 0.686** −0.940** 0.605** 总酚 0.112 0.776** 0.19 −0.019 0.527** MDA 0.056 −0.618** 0.906** −0.423* SOD 0.233 −0.097 0.705** APX −0.743** 0.616** LOX −0.556** 注:**表示极显著相关(P<0.01);*表示显著相关(P<0.05)。 3. 讨论
3.1 1-MCP处理对西葫芦果实采后生理品质的影响
常温贮藏期间,营养物质会随着贮藏时间的延长不断损失,果实出现萎缩、组织松软、内容物流出等现象,严重影响商品价值。本实验研究表明,1-MCP处理可以降低西葫芦果实呼吸强度,减少失水,这与前人的研究结果一致[22]。西葫芦果实的TSS变化趋势为先上升后下降,表明可能是西葫芦果实在贮藏初期过程中呼吸作用强消耗能量,导致失水,可溶性固形物含量出现短暂上升趋势。可溶性蛋白也是衡量果实成熟衰老的重要指标,是绝大多数果蔬主要的底物和能量物质[23],本实验结果表明1-MCP处理保持了西葫芦果实可溶性蛋白的含量,这与李君兰等[24]的研究结果一致。许多研究结果表明[25-26],酚类物质和VC是果实中重要的抗氧化物质,通过非酶促代谢参与着机体内抗氧化过程,在清除ROS自由基方面具有重要作用。相关性分析的结果显示:VC与MDA呈极显著性负相关、与APX呈极显著性正相关,总酚与SOD呈极显著性正相关(P<0.01),表明VC抑制MDA的积累,并且和总酚一同作为抗氧化物质与SOD、APX协同作用参与机体内抗氧化过程。本研究结果表明,1-MCP处理能够保持西葫芦较高的TSS、总酚和VC含量,减少贮藏期间的损失。综上所述,1-MCP在贮藏期间降低了西葫芦果实呼吸强度、失重率,维持了TSS、可溶性蛋白、总酚、VC的含量,保持了果实良好的品质。
3.2 1-MCP处理对西葫芦果实线粒体抗氧化作用的影响
ROS与果蔬成熟衰老密切相关,其一方面它协助细胞进行正常的生理活动,另一方面它的积累会引起线粒体内膜通透性、膜内氧化还原电势降低,引起膜脂过氧化,对细胞器造成严重的损伤,破坏线粒体的完整性;ROS还参与线粒体凋亡、细胞分化及基因表达的调控[27]。当然,线粒体中也存在SOD、CAT、APX等去消除ROS产生的损伤,如SOD,其作为抗氧化的第一道防线,能催化超氧化物如O2−·分解生成水(H2O)和氧气(O2)[28],CAT的作用是分解过氧化氢H2O2生成H2O,APX可以催化分解抗坏血酸盐[29]。线粒体内多种抗氧化物酶相互协作,共同抵御ROS,保护线粒体膜结构和功能,从而保持线粒体膜的氧化平衡和膜结构完整性。前人研究表明,15 μmol·L−1一氧化氮(NO)处理可以减少冷藏桃果实线粒体ROS含量的累积,保持较高的SOD、CAT、POD的活性[30]。本研究中,在贮藏期间H2O2含量和O2−·产生速率均是对照组较处理组高,且1-MCP处理也保持了线粒体较高的SOD、APX的活性,推测是1-MCP通过提高果实线粒体SOD、APX的活性,抑制ROS的积累,维持ROS平衡。这一结果与胡珊等[31]的报道一致。LOX是引起膜脂过氧化作用的重要酶,根据相关性分析的结果显示,MDA与APX呈极显著性正相关、与LOX呈极显著性正相关(P<0.01),表明西葫芦果实线粒体内的抗氧化物酶APX参与抗氧化过程,抑制了MDA含量的上升,且线粒体MDA的含量会影响膜脂过氧化程度高低。本研究结果表明,1-MCP处理能有效抑制LOX活性的上升,延缓西葫芦线粒体MDA含量的上升,表明1-MCP减轻西葫芦膜脂过氧化作用明显,这与Jing等[32]的研究结果一致。综上所述,与对照组相比,1-MCP处理可以提高西葫芦果实线粒体抗氧化酶SOD、APX的活性,降低LOX活性,清除线粒体过多的ROS、减少MDA的积累,降低膜结构损伤,保持线粒体内氧化还原平衡,保持线粒体正常功能。
3.3 1-MCP处理对西葫芦果实线粒体能量代谢的影响
线粒体是能量产生的主要场所,SDH、CCO都是线粒体内膜上的标志酶,其活性能反映线粒体能量状态,SDH在TCA中将琥珀酸转变为延胡索酸,为呼吸链提供电子进行产能[33]。CCO作为电子传递链末端的酶,具有质子泵的作用可将H+泵到膜间,同时能将电子从细胞色素C传递给O2,在电子传递过程中生成大量ATP,用于生物体内各种生命活动[8]。ATP的生成与能量代谢相关酶密不可分,线粒体膜结构和功能的损伤会使得这些酶的活性受到影响,使得果蔬在采后出现氧化胁迫、呼吸链受损,造成ATP合成速度减慢、细胞能量供应不足、代谢紊乱,细胞出现死亡[28]。H+-ATPase是细胞膜质子泵,泵出H+在膜内外产生电化学梯度,产生跨膜电动势,催化ATP的生成,Ca2+-ATPase是线粒体内膜Ca2+泵,维持膜内外Ca2+的平衡,若Ca2+平衡被打破,则会导致线粒体膜受损,引发线粒体功能障碍[34]。本研究结果表明,在贮藏前期,西葫芦果实的SDH、CCO、H+-ATPase、Ca2+-ATPase有一个上升的过程,这可能是由于前期果实在面对逆境胁迫所表现出的为抵御胁迫刺激能量上升的应激反应,在贮藏后期,西葫芦果实的SDH、CCO、H+-ATPase、Ca2+-ATPase活性随贮藏时间都呈现下降的趋势,这是由于线粒体ROS爆发,线粒体膜完整性被破坏,相关酶活性降低,线粒体功能受到损伤、果实组织逐渐衰老。1-MCP处理提高了SDH、CCO、H+-ATPase、Ca2+-ATPase活性,保持正常的能量供应,保证机体正常生命活动。研究表明NO处理可以维持桃果实贮藏期间SDH、CCO的活性,且能量不足会产生更多的ROS攻击细胞膜,因此,维持果实内高能量水平对保持果实品质具有重要的作用[35]。此外,能量代谢与抗氧化代谢还存在密切的关系,根据相关性分析的结果显示,SDH与果实品质相关指标TSS、可溶性蛋白、总酚、VC呈极显著性正相关、与抗氧化物酶SOD、APX呈极显著性正相关、与LOX呈极显著性负相关(P<0.01)。这表明维持能量水平可以保持果实品质,并可能从一定水平上调节诱导抗氧化物酶的活性,从而调控果实成熟衰老,这与前人的研究结果一致[36]。
4. 结论
20 μL·L−1 1-MCP处理对西葫芦果实的品质和生理生化特性的影响主要表现在:1-MCP处理可以抑制西葫芦果实生理活动,降低呼吸强度和失重率,减少TSS、可溶性蛋白含量的损失,维持较高的总酚、VC等抗氧化物质含量,有利于保持果实良好的品质,延缓衰老。更深入地分析发现,20 μL·L−1的1-MCP处理可以抑制果实线粒体ROS含量的上升,延缓MDA、LOX上升,保持线粒体抗氧化物酶SOD、APX的活性,减轻膜脂过氧化,保护线粒体膜免受损伤;在能量水平上,20 μL·L−1 1-MCP处理还能保持西葫芦果实线粒体能量代谢酶SDH、CCO、H+-ATPase、Ca2+-ATPase的活性,维持果实组织的正常能量代谢。以上研究结果为1-MCP处理应用于果蔬采后保鲜方面提供了理论依据,今后的努力方向应从分子水平上对1-MCP调控成熟衰老机理进行深入研究。
-
![]()
![]()
图 2 1-MCP处理对西葫芦果实呼吸强度(A)、线粒体MDA(B)、H2O2含量(C)、O2−·产生速率变化(D)的影响
Figure 2. Effects of 1-MCP treatment on respiration intensity (A), mitochondrial MDA (B), H2O2 content (C) and O2−· (D) production rate of zucchini fruits
![]()
图 3 1-MCP对西葫芦果实线粒体SOD(A)、APX(B)和LOX(C)活性的影响
Figure 3. Effects of 1-MCP on the SOD (A), APX (B) and LOX (C) activities in mitochondria of zucchini fruits
![]()
图 4 1-MCP处理对西葫芦线粒体SDH(A)、CCO(B)、H+-ATPase(C)和Ca2+-ATPase(D)活性的影响
Figure 4. Effects of 1-MCP on SDH (A), CCO (B), H+-ATPase (C) and Ca2+-ATPase (D) activity of zucchini fruits
表 1 1-MCP处理西葫芦果实采后各项生理指标的相关性
Table 1 Correlation of physiological indexes of postharvest zucchini fruits treated by 1-MCP
项目 TSS 可溶性蛋白 VC 总酚 MDA SOD APX LOX SDH 呼吸强度 0.233 0.316 0.268 −0.398* −0.352 −0.524** 0.339 −0.383* −0.188 TSS 0.944** 0.803** 0.519** −0.702** 0.383* 0.709** −0.831** 0.670** 可溶性蛋白 0.924** 0.326 −0.837** 0.312 0.797** −0.916** 0.684** VC 0.066 −0.954** 0.158 0.686** −0.940** 0.605** 总酚 0.112 0.776** 0.19 −0.019 0.527** MDA 0.056 −0.618** 0.906** −0.423* SOD 0.233 −0.097 0.705** APX −0.743** 0.616** LOX −0.556** 注:**表示极显著相关(P<0.01);*表示显著相关(P<0.05)。  下载: 导出CSV
下载: 导出CSV
-
[1] 刘宜生. 西葫芦史话[J]. 中国瓜菜,2008(1):49−50. [LIU Y S. Zucchini history[J]. Chinese Vegetables,2008(1):49−50. doi: 10.3969/j.issn.1673-2871.2008.01.023 [2] 盖晓阳, 张敏, 李佳乐, 等. 不同湿度的热空气处理对西葫芦果实低温贮藏抗冷性和活性氧代谢的影响[J]. 安徽农业大学学报,2021,48(3):382−389. [GAI X Y, ZHANG M, LI J L, et al. Effects of hot air treatment with different humidity on cold resistance and active oxygen metabolism of Zucchini fruits during cryogenic storage[J]. Journal of Anhui Agricultural University,2021,48(3):382−389. doi: 10.13610/j.cnki.1672-352x.20210706.011 [3] 李富军, 杨洪强, 翟衡, 等. 1-甲基环丙烯延缓果实衰老作用机制研究综述[J]. 园艺学报,2003(3):361−365. [LI F J, YANG H Q, ZHAI H, et al. Review on the mechanism of 1-methylcyclopropene inhibiting fruit senescence[J]. Chinese Journal of Horticulture,2003(3):361−365. doi: 10.3321/j.issn:0513-353X.2003.03.031 [4] GUO Y Z, LU Y, DU C, et al. An antioxidant system through conjugating superoxide dismutase onto metal-organic framework for cardiac repair[J]. Bioactive Materials,2022,10:56−67. doi: 10.1016/j.bioactmat.2021.08.019
[5] 王美红, 李一诺, 葛柯良, 等. 1-MCP(1-甲基环丙烯)与CaCl2处理对桃果实采后贮藏品质及生理的影响[J]. 植物生理学报,2021,57(5):1113−1122. [WANG M H, LI Y N, GE K L, et al. Effects of 1-MCP (1-methylcyclopropylene) and CaCl2 treatment on storage quality and physiology of peach fruit after harvest[J]. Chinese Journal of Plant Physiology,2021,57(5):1113−1122. [6] 张明明, 热合满·艾拉, 王玉红, 等. 乙烯与1-MCP处理对伯谢克辛甜瓜采后活性氧代谢的影响[J]. 食品工业科技,2018,39(18):270−273,210. [ZHANG M M, RE A, WANG Y H, et al. Effects of ethylene and 1-MCP treatments on postharvest reactive oxygen metabolism of muskmelon[J]. Science and Technology of Food Industry,2018,39(18):270−273,210. doi: 10.13386/j.issn1002-0306.2018.18.046 [7] SONG L L, YI R X, LUO H B, et al. Postharvest 1-methylcyclopropene application delays leaf yellowing of pak choi (Brassica rapa subsp. chinensis) by improving chloroplast antioxidant capacity and maintaining chloroplast structural integrity during storage at 20 ℃[J]. Scientia Horticulturae,2020,270:109466. doi: 10.1016/j.scienta.2020.109466
[8] KARBOWSKI M, OSHIMA Y, VERHOEVEN N. Mitochondrial proteotoxicity: Implications and ubiquitin-dependent quality control mechanisms[J]. Cellular Molecular Life Sciences,2022,79(11):574. doi: 10.1007/s00018-022-04604-8
[9] 王志华, 贾朝爽, 王文辉, 等. 低温贮藏对‘金红’苹果能量代谢和品质的影响[J]. 园艺学报,2020,47(12):2277−2289. [WANG Z H, JIA C S, WANG W H, et al. Effects of low temperature storage on energy metabolism and quality of 'Jinhong' apple[J]. Journal of Horticulture,2020,47(12):2277−2289. doi: 10.16420/j.issn.0513-353x.2020-0418 [10] 张正敏, 杨艺琳, 李美琳, 等. 2,4-表油菜素内酯处理对桃果实软腐病及能量代谢的影响[J]. 食品科学,2019,40(5):207−213. [ZHANG Z M, YANG Y L, LI M L, et al. Effects of 2,4-epbrassinolide treatment on soft rot and energy metabolism of peach fruit[J]. Food Science,2019,40(5):207−213. doi: 10.7506/spkx1002-6630-20180828-312 [11] 曹建康, 姜微波, 赵玉梅. 果蔬采后生理生化实验指导[M]. 北京: 中国轻工业出版社, 2013: 39-41 CAO J K, JIANG W B, ZHAO Y M. Experimental Instruction of postharvest physiology and biochemistry of fruits and vegetables[M]. Beijing: China Light Industry Press, 2013: 39-41.
[12] QIU X L, GUO Y X, ZHANG Q F. Chemical profile and antioxidant activity of Gynura bicolor DC ethanolic extract[J]. International Journal of Food Properties,2018,21(1):407−415. doi: 10.1080/10942912.2018.1424199
[13] MILLAR A H, LIDDELL A, LEAVER C J. Isolation and subfractionation of mitochondria from plants[M]. University of Oxford, Methods in Cell Biology, 2001, 65: 65−90.
[14] LI R R, ZHENG Q W, LU J L, et al. Chemical composition and deterioration mechanism of Pleurotus tuoliensis during postharvest storage[J]. Food Chemistry,2021,338:127731. doi: 10.1016/j.foodchem.2020.127731
[15] 贾乐, 韩延超, 房祥军, 等. 褪黑素处理对香菇采后品质及活性氧代谢的影响[J]. 食品科学,2021,42(23):229−236. [JIA L, HAN Y C, FANG X J, et al. Effects of melatonin treatment on postharvest quality and active oxygen metabolism of Lentinus edodes[J]. Food Science,2021,42(23):229−236. doi: 10.7506/spkx1002-6630-20210316-201 [16] ZHENG X L, TIAN S P, MENG X H, et al. Physiological and biochemical responses in peach fruit to oxalic acid treatment during storage at room temperature[J]. Food Chemistry,2007,104(1):156−162. doi: 10.1016/j.foodchem.2006.11.015
[17] SHARMA S, SEHRAWAT A, DESWAL R. Asada-Halliwell pathway maintains redox status in Dioscorea alata tuber which helps in germination[J]. Plant Science,2016,250:20−29. doi: 10.1016/j.plantsci.2016.05.013
[18] MAO L C, WANG G Z, ZHU C G, et al. Involvement of phospholipase D and lipoxygenase in response to chilling stress in postharvest cucumber fruits[J]. Plant Science,2007,172(2):400−405. doi: 10.1016/j.plantsci.2006.10.002
[19] ACKRELL B A, KEAMERY E B, SINGER T P. Mammalian succinate dehydrogenase[J]. Methods in Enzymology,1978,53:466−483.
[20] ERREDE B, KAMEN M D, HATEFI Y. Preparation and properties of complex IV (Ferrocytochrome c: Oxygen oxidoreductase EC 1.9.3.1)[M]. Methods in Enzymology, Academic Press, 1978, 53: 40−47.
[21] JIN P, ZHU H, WANG J, et al. Effect of methyl jasmonate on energy metabolism in peach fruit during chilling stress[J]. Journal of the Science of Food and Agriculture,2013,93(8):1827−1832. doi: 10.1002/jsfa.5973
[22] XU F X, LIU S Y, LIU Y F, et al. Effectiveness of lysozyme coatings and 1-MCP treatments on storage and preservation of kiwifruit[J]. Food Chemistry,2019,288:201−207. doi: 10.1016/j.foodchem.2019.03.024
[23] 安容慧, 陈皖豫, 胡花丽, 等. 1-甲基环丙烯对娃娃菜贮藏品质及抗氧化活性的影响[J]. 食品与发酵工业,2021,47(20):194−203. [AN R H, CHEN W Y, HU H L, et al. Effects of 1-methylcyclopropene on storage quality and antioxidant activity of baby Chinese cabbage[J]. Food and Fermentation Industries,2021,47(20):194−203. doi: 10.13995/j.cnki.11-1802/ts.026847 [24] 李君兰, 杨澜, 吴潇霞, 等. 葡萄糖酸钙采前处理对鲜枣果实低温贮藏品质及活性氧代谢的影响[J]. 食品与发酵工业,2019,45(7):144−150. [LI J L, YANG L, WU X X, et al. Effects of calcium gluconate pretreatment on low temperature storage quality and active oxygen metabolism of fresh Jujube fruits[J]. Food and Fermentation Industries,2019,45(7):144−150. doi: 10.13995/j.cnki.11-1802/ts.018672 [25] LI H Y, XIE L, MA Y, et al. Effects of drying methods on drying characteristics, physicochemical properties and antioxidant capacity of okra[J]. LWT-Food Science and Technology,2018,101:630−638.
[26] WANG Y T, LI L X, LIU H, et al. Bioactive compounds and in vitro antioxidant activities of peel, flesh and seed powder of kiwifruit[J]. International Journal of Food Science & Technology,2018,53(9):2239−2245.
[27] HUANG D D, HU S, ZHU S H, et al. Regulation by nitric oxide on mitochondrial permeability transition of peaches during storage[J]. Plant Physiology and Biochemistry,2019,138:17−25. doi: 10.1016/j.plaphy.2019.02.020
[28] WANG Y, BRANICKY R, NOE A, et al. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling[J]. The Journal of Cell Biology,2018,217(6):1915−1928. doi: 10.1083/jcb.201708007
[29] PANDEY S, FARTYAL D, AGARWAL A, et al. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase[J]. Frontiers in Plant Science,2017,8:581. doi: 10.3389/fpls.2017.00581
[30] 王彩云. NO对桃果实线粒体功能及氧化损伤的调控作用[D]. 泰安: 山东农业大学, 2021 WANG C Y. Regulation of NO on mitochondrial function and oxidative damage in peach fruit[D]. Taian: Shandong Agricultural University, 2021.
[31] 胡珊, 叶志, 冯建荣, 等. 一氧化氮对低温贮藏桃果实线粒体氧化损伤的影响[J]. 新疆农业科学,2018,55(12):2157−2165. [HU S, YE Z, FENG J R, et al. Effects of nitric oxide on mitochondrial oxidative damage of peach fruits stored at low temperature[J]. Xinjiang Agricultural Sciences,2018,55(12):2157−2165. [32] JING G Q, ZHOU J, ZHU S H. Effects of nitric oxide on mitochondrial oxidative defence in postharvest peach fruits[J]. Journal of the Science of Food and Agriculture,2016,96(6):1997−2003. doi: 10.1002/jsfa.7310
[33] LI L, ZHANG Y Y, FAN X L, et al. Relationship between activated oxygen metabolism and browning of 'Yali' pears during storage[J]. Journal of Food Processing and Preservation,2020,44(4):14392.
[34] HARRIS J R, BOEKEMA E J. Protein complexes: Structure and function volume 87|| Mitochondrial respiratory chain complexes[J]. Subcellular Biochemistry,2018,87(7):167−227.
[35] 张小康, 上官相超, 陈长宝, 等. 一氧化氮对冷藏桃果实能量代谢的调控作用[J]. 保鲜与加工,2019,19(4):1−9. [ZHANG X K, SHANG GUAN X C, CHEN C B, et al. Regulation effect of nitric oxide on energy metabolism of frozen peach fruit[J]. Preservation and Processing,2019,19(4):1−9. doi: 10.3969/j.issn.1009-6221.2019.04.001 [36] 张群. 欧亚种提子类葡萄贮藏期间果实能量亏损与品质劣变机理研究[D]. 长沙: 中南林业科技大学, 2018 ZHANG Q. Mechanism of energy loss and quality deterioration of Eurasian seed grape during storage[D]. Changsha: Central South University of Forestry and Technology, 2018.





 下载:
下载:




计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
