


Investigation the Effects and Mechanisms of Autolytic Active Peptides from Shrimp Head on Anti-fatigue in Mice Based on the Keap1/Nrf2/ARE Signaling Pathway
-
摘要: 为研究虾头自溶活性多肽(SAP)对小鼠的抗疲劳作用,经灌胃给予小鼠低、中、高剂量虾头自溶活性多肽28 d后,分别测定其负重游泳时间、肝组织液中丙二醛含量、血乳酸含量、血尿素氮含量、乳酸脱氢酶活力、还原型谷胱甘肽含量、肝/肌糖原含量以及Keap1、Nrf2、HO-1和NQO-1 mRNA表达,并观察各组小鼠肝脏组织形态。结果表明:与正常空白组相比,虾头自溶活性多肽各剂量组小鼠负重游泳力竭时间显著延长(P<0.05),虾头自溶活性多肽高剂量组(600 mg/kg·d)丙二醛、血乳酸、血尿素氮含量显著降低(P<0.05)、乳酸脱氢酶、还原型谷胱甘肽、肝糖原含量显著升高(P<0.05),Nrf2、HO-1和NOQ-1 mRNA表达显著上调(P<0.05),Keap1 mRNA表达显著下调(P<0.05),抗疲劳效果与牛磺酸阳性对照组相当。此外,观察各组小鼠肝脏切片组织形态,虾头自溶活性多肽各剂量组与正常空白组形态类似,表明短期持续摄入适量虾头自溶活性多肽不影响小鼠的肝脏正常代谢功能,不会对其肝脏造成损伤。结果表明,虾头自溶活性多肽可激活小鼠体内Keap1/Nrf2/ARE信号通路,减轻氧化应激、保护细胞免受损伤以及调节能量代谢,从而缓解其疲劳症状,具有显著的抗疲劳作用Abstract: To investigate the influence of shrimp head autolytic active peptides (SAP) on the anti-fatigue ability of mice, low, middle, and high doses of SAP were administered orally to mice for 28 d. Subsequently, load-bearing swimming time, malondialdehyde content in liver tissue, blood lactic acid content, blood urea nitrogen content, lactate dehydrogenase activity, reduced glutathione content, liver/muscle glycogen content, and mRNA expression of Keap1, Nrf2, HO-1, and NQO-1 were measured and the liver tissue morphology of each group of mice was also observed. The results showed that compared with the normal control group (NC), the load-bearing swimming time of mice in each dose group of SAP was significantly prolonged (P<0.05), while the high-dose group of SAP (600 mg/kg·d) exhibited significantly decreased levels of malondialdehyde, blood lactic acid, and blood urea nitrogen (P<0.05), significantly increased levels of lactate dehydrogenase, reduced glutathione, and liver glycogen (P<0.05), significantly upregulated mRNA expression of Nrf2, HO-1, and NQO-1 (P<0.05), and significantly downregulated mRNA expression of Keap1 (P<0.05). The anti-fatigue effect was comparable to that of taurine positive control group (TP). In addition, by observing the tissue morphology of liver slices in each group of mice, each dose group of SAP showed similar morphology to the normal blank group, indicating that short-term continuous intake of appropriate amounts of SAP does could not affect the normal metabolic function of the mice liver and not cause damage to its liver. The results indicated that shrimp head SAP can activate the Keap1/Nrf2/ARE signaling pathway in mice, alleviate oxidative stress, protect cells from damage, and regulate energy metabolism, thus alleviating fatigue symptoms and exhibiting significant anti-fatigue effects.
-
湛江被称为“中国对虾之都”,对虾产量约占全国总产量的三分之一,是我国对虾种苗繁育、养殖、加工及交易主要基地。目前,湛江对虾加工企业超过250家,年对虾加工量超过80万吨,主要产品为虾滑、虾仁、去头虾等产品,对虾产品在加工过程中会产生大量的虾头、虾壳等副产物,约占总质量的30%,常作为甲壳素生产原料使用。对虾加工副产物除含有甲壳素外,还含有丰富的蛋白质,然而在甲壳素生产过程中,蛋白质因强酸强碱作用失去营养价值,且随污水排放,浪费资源的同时对环境也造成了严重污染。因此,如何综合利用对虾加工副产物中的蛋白质,提高产品附加值成为目前亟需解决的关键问题。
生物活性肽是一类对机体功能具有积极作用的特异性蛋白片段,在蛋白序列中通常不具有活性,但可通过酶解或微生物发酵等方式释放后发挥作用[1]。除营养价值外,生物活性肽还具有广泛的生理调节作用,如抗氧化、抗疲劳、降血压、抗菌等,其中抗疲劳肽能够维持机体的运动能力,快速消除疲劳、恢复体力,其已成为从事体力者的一种重要的功能性食品基料[2]。海洋生物作为抗疲劳活性肽的重要来源,Feng等[3]、左爱华等[4]以海参和海参内脏为原料,利用酶解法制备的小分子量海参肽对小鼠具有抗疲劳作用;贝类经复合蛋白酶酶解后得到的媒介产物也具有较明显的抗疲劳功效,可以延长小鼠力竭游泳时间,提高肝糖原含量、降低血清尿素氮和血乳酸[5−6]。此外,由于虾头含有丰富的蛋白质,含量高达60%(以干基计),虾头中蕴含的内源蛋白酶在特定条件下可引起虾头发生自溶解作用,大分子蛋白质被蛋白酶降解为可溶性蛋白或多肽片段[7]。如何高值化利用虾头也成为众多学者研究的热点。如朱国萍等[8]发现凡纳滨对虾虾头内源酶自溶产物具有血管紧张素转换酶(Angiotensin-I-converting enzyme,ACE)抑制活性,并从中分离纯化得到两条ACE抑制肽,氨基酸序列为Tyr-Pro和Leu-Pro/Ile-Pro;张风等[9]利用凡纳滨对虾的虾头虾壳制备酶解物,研究结果表明该酶解液具有优秀的抗氧化活性;王晋等[10]从南美白对虾虾头、虾壳蛋白质酶解液中分离纯化了具有抗氧化功能的多肽混合物,其抗氧化能力与VC相当。
自由基的大量产生是引起肌体疲劳的重要原因,众多研究表明虾蛋白多肽具有显著的抗氧化活性,推测凡纳滨对虾多肽亦具有抗疲劳功能。为此,本实验以凡纳滨对虾加工副产物(虾头)为原料,通过内源酶自溶制备蛋白质多肽,研究多肽混合物的抗疲劳能力,并对其相关作用机制进行探究,以期为对虾副产物的高值化利用和精深加工提供技术参考。
1. 材料与方法
1.1 材料与仪器
鲜活凡纳滨对虾 广东省湛江市南方水产交易市场,摘取头部组织,备用;SPF级雄性昆明小鼠 6~8周龄,体质量(25±2)g,湖南斯莱克景达实验动物有限公司(动物许可证号:SCXK(湘)2019-0004);血乳酸测定试剂盒、血乳酸脱氢酶测定试剂盒、血尿素氮检测试剂盒、丙二醛测定试剂盒、还原型谷胱甘肽测定试剂盒、肌/糖原测定试剂盒、Folin-酚试剂盒 苏州格锐思生物科技有限公司。
Varioskan Flash多功能酶标仪、StepOnePlus实时荧光定量PCR仪 赛默飞世尔科技(苏州)有限公司;ICEN-24R台式高速冷冻离心机、Nano-300微量分光光度计 杭州奥盛仪器有限公司;A200梯度PCR扩增仪 杭州朗基科学仪器有限公司;BX43正置显微镜 日本奥林巴斯仪器有限公司;WTM-1812G膜分离设备 杭州沃腾膜工程有限公司。
1.2 实验方法
1.2.1 凡纳滨对虾虾头自溶液及活性多肽的制备
1.2.1.1 凡纳滨对虾虾头自溶液的制备
参考Cao等[11]的方法,并经适当修改,将虾头与超纯水1:2混合,冰浴下匀浆(10000 r/min),用稀氢氧化钠溶液调节pH至7.5,在50 ℃ ,120 r/min的条件下,振荡反应2 h(内源酶在50 ℃时自溶效率最高)。待反应结束后,100 ℃沸水浴灭酶10 min,灭酶后的酶解液离心处理15 min (10000 r/min),上清液即为虾头自溶液。
1.2.1.2 虾头自溶活性多肽(SAP)的制备及测定
虾头自溶液经水性滤膜(0.45 μm)抽滤,滤出液置于截留分子量为3000 Da的超滤膜中处理,超滤工作压力为0.1 MPa,收集透过液,冷冻干燥后得到多肽冻干粉,以牛血清白蛋白作为标准溶液,采用Folin-酚法测定样品中的多肽含量[12]。按照Folin-酚法试剂盒操作说明进行测定,在波长500 nm 处测定吸光度值,绘制标准曲线、计算样品的多肽含量。
1.2.2 动物分组及给药
实验动物选用6~8周龄雄性昆明小鼠。试验用随机数法分为正常空白组(Normal control group,NC)、牛磺酸阳性对照组(Taurine positive control group,TP)、虾头自溶活性多肽低(Shrimp head autolysis active peptides low dose group,SAP-L)、中(Shrimp head autolysis active peptides medium dose group,SAP-M)、高剂量组(Shrimp head autolysis active peptides high dose group,SAP-H),参考姜丽冬等[13]研究结果,本实验灌胃剂量分别设为SAP-L(200 mg/kg·d)、SAP-M(400 mg/kg·d)和SAP-H(600 mg/kg·d),NC给予蒸馏水,TP给予中等剂量的牛磺酸(400 mg/kg·d),各组饲养时间28 d。上述实验动物饲养于重庆第二师范学院动物实验中心。此研究已获得重庆第二师范学院动物伦理委员会批准,批准号为:2023030701B,并严格遵循国家以及国际动物实验伦理原则。
1.2.3 小鼠负重游泳试验
参考陈立艺等[14]相关方法,选取正常空白组、低、中、高剂量组和阳性对照组实验小鼠,每组各8只,共40只小鼠用于负重游泳实验。采用50 cm×50 cm×40 cm游泳箱,水深≥30 cm,水温(25±1.0)℃,末次给予受试样品30 min后,将尾根部负荷5%体重铅皮的小鼠置于游泳箱中游泳,记录小鼠从入水游泳至力竭的时间(小鼠游泳力竭的判定标准为没入水面7 s不再上浮)。
1.2.4 虾头自溶活性多肽的抗疲劳能力评价
1.2.4.1 血清、肌组织以及肝组织的采集与制备
小鼠力竭后眼球取血,在4 ℃条件下离心15 min(4000 r/min)制备血清,取上清液于试管中,−20 ℃保存备用。眼球取血后将小鼠脱颈处死,取其后腿肌肉(股四头肌)和肝脏组织,分别用生理盐水漂洗3次,滤纸吸干后,称量0.5 g标本加入4.5 mL生理盐水,将其置于匀浆机中低温粉碎匀浆,在4 ℃条件下离心15 min(4000 r/min),取上清液于试管中,−20 ℃保存备用
1.2.4.2 抗疲劳生化指标检测
将制备的血清用于检测血清乳酸、尿素氮含量和乳酸脱氢酶活性,肝脏、腿肌组织液用于检测肝、肌糖原含量,并检测肝组织液中丙二醛和还原型谷胱甘肽的含量,各生化指标均按照试剂盒方法操作。
1.2.4.3 小鼠组织的病理学观察
取解剖后小鼠的肝脏组织分别用10%福尔马林组织固定液固定。送至Servicebio公司采用苏木精-伊红(Hematoxylin-eosin,H&E)染色法进行制片,最后在光学显微镜下观察小鼠肝脏组织的病理学变化,详细步骤参照Chamberland等[15]的研究方法。
1.2.4.4 RNA提取和荧光定量分析
分别取50 mg小鼠肝脏组织,用无菌生理盐水洗净后,加入200 μL TRIzol试剂匀浆后加入300 μL TRIzol试剂提取组织中的RNA,再将提取到的RNA用超微量分光光度计检测其浓度和纯度。按照cDNA逆转录试剂说明书,将RNA转录成cDNA,按2 μL模板(cDNA)、4 μL引物、10 μL SYBR及4 μL无菌超纯水的比例加入八连管,将八连管在荧光PCR仪器内按20 μL反应体积进行PCR反应。PCR扩增条件为:95 ℃预变性5 min,95 ℃变性10 s,60 ℃退火20 s,72 ℃延伸20 s,整个过程进行 40次循环。扩增结束后,以内参基因β-actin为基准,通过公式2−ΔΔCT计算各基因mRNA的相对表达量。所测基因分别为Kelch样环氧氯丙胺相关蛋白1(Kelch-like epichlorohydrin-associated protein 1,Keap1)、核因子E2相关因子2(Nuclear factor erythtoid 2 related factor 2,Nrf2)、血红素氧化酶1(Heme oxygenase 1,HO-1)和还原型辅酶/醌氧化还原酶-1(Quinone oxidoreductase-1,NQO-1)。上述基因引物由Invitrogen公司设计并合成,引物序列见表1。
表 1 引物序列Table 1. Primers sequence基因 引物序列 引物长度(bp) 上游引物(5′~3′) 下游引物(5′~3′) Beta-actin GGCTGTATTCCCCTCCATCG CCAGTTGGTAACAATGCCATGT 154 Keap1 TCGAAGGCATCCACCCTAAG CTCGAACCACGCTGTCAATCT 135 Nrf2 TAGATGACCATGAGTCGCTTGC GCCAAACTTGCTCCATGTCC 153 HO-1 GATAGAGCGCAACAAGCAGAA CAGTGAGGCCCATACCAGAAG 111 NQO-1 AGGATGGGAGGTACTCGAATC AGGCGTCCTTCCTTATATGCTA 144 1.3 数据处理
采用SPSS 26.0软件进行统计学分析,统计结果以平均值±标准差表示。多组间比较采用单因素方差分析,两两比较采用LSD检验,以P<0.05为差异有统计学意义。
2. 结果与分析
2.1 虾头自溶活性多肽冻干粉中多肽含量
实验测定的多肽含量线性回归方程为y=0.0005x−0.0013,线性回归系数R2为0.999,说明在测定浓度范围内多肽质量浓度与吸光度线性关系良好。虾头自溶活性多肽冻干粉中多肽含量为91.75%±0.25%,说明虾头中的蛋白质经内源酶酶解、超滤处理后,杂质去除彻底,故多肽含量较高。
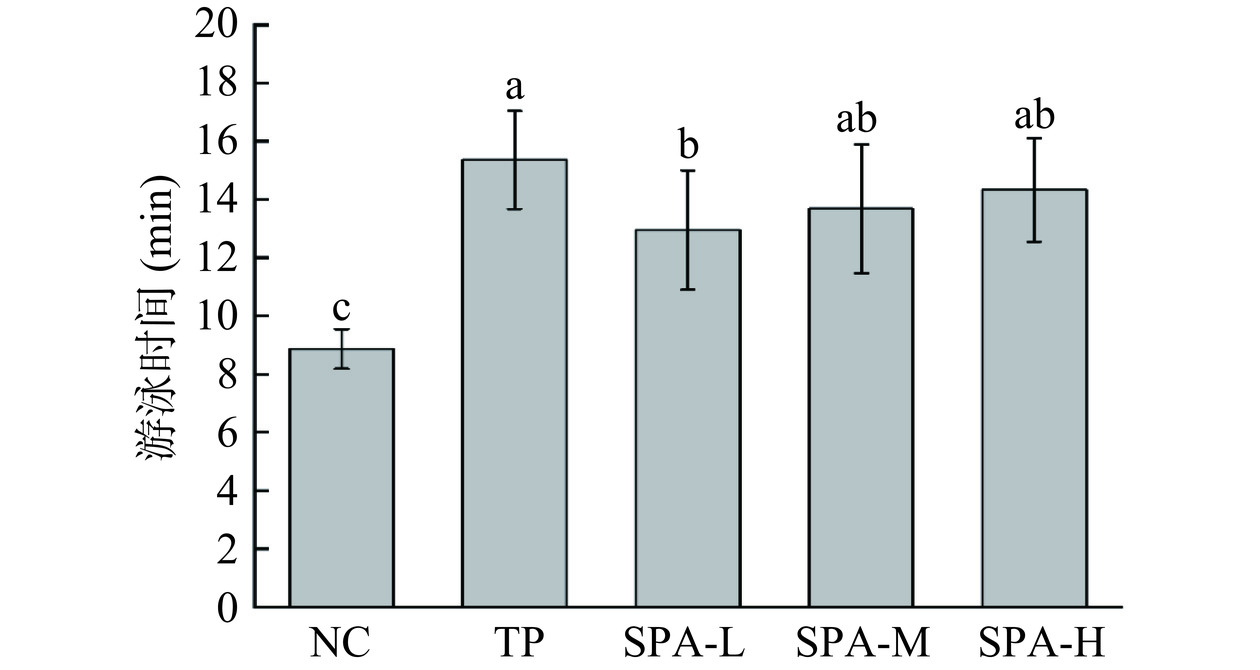
2.2 虾头自溶活性多肽对小鼠负重游泳时间的影响
由图1可知,虾头自溶活性多肽各剂量组与NC组相比,小鼠负重游泳时间显著延长(P<0.05),SAP-L、SAP-M和SAP-H组分别延长46.0%、54.2%和61.6%,TP组延长73.2%。实验表明,随着虾头自溶活性多肽剂量的增加,小鼠的负重游泳力竭时间逐渐延长,SAP-H组与TP组效果相当,表明虾头自溶活性多肽具有增加小鼠抗疲劳能力的作用。
![]()
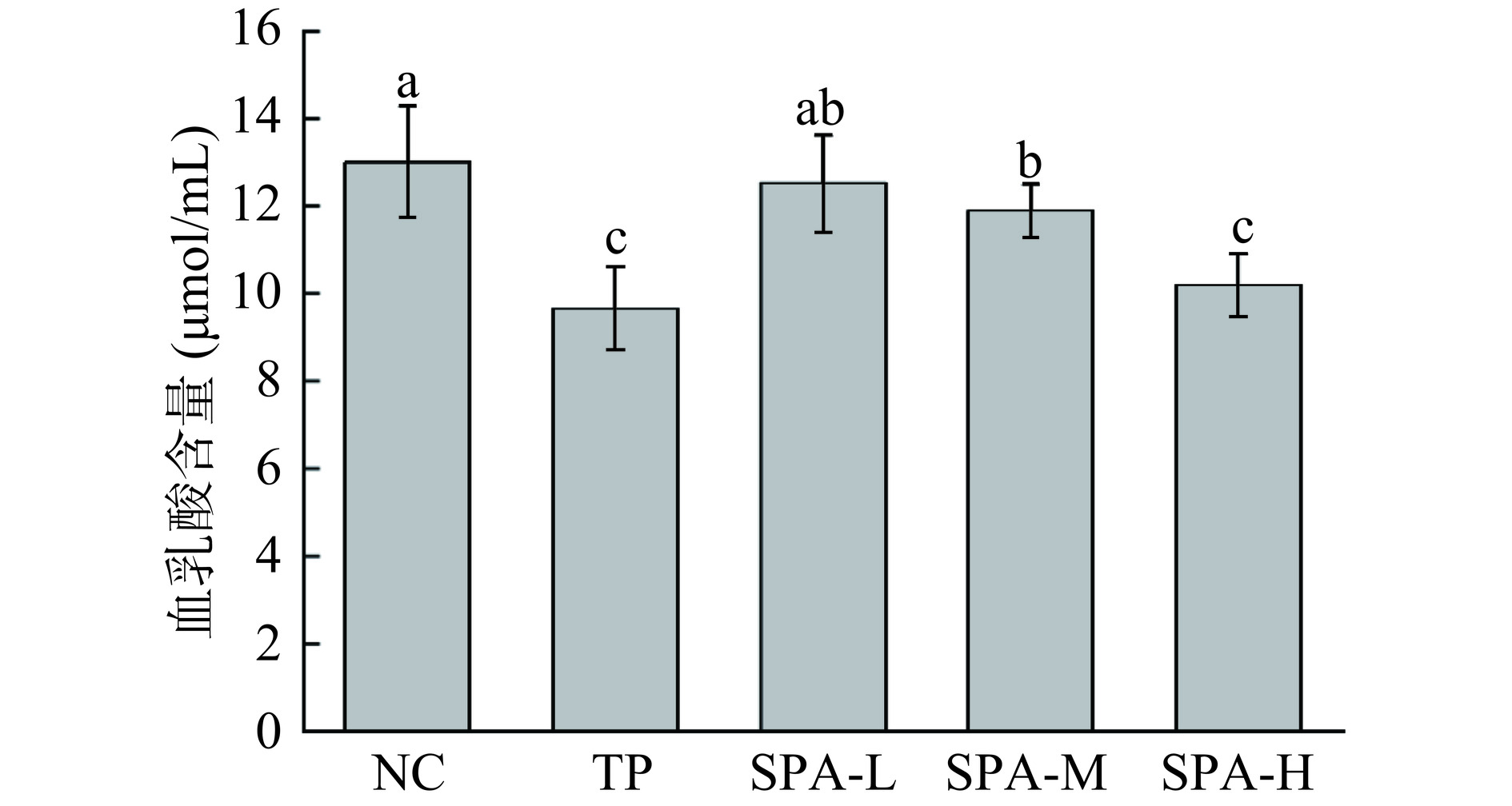
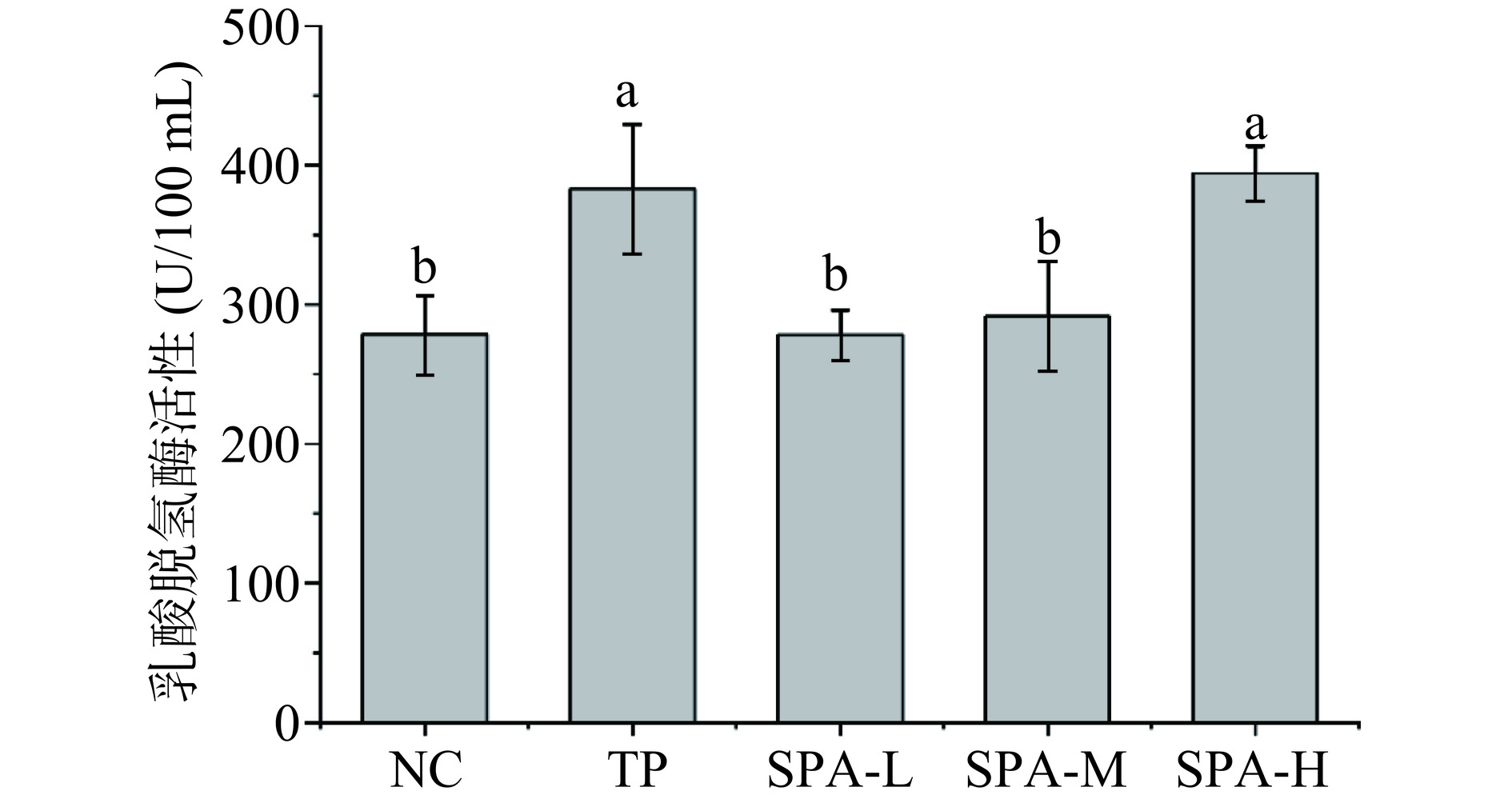
2.3 虾头自溶活性多肽对力竭游泳小鼠血乳酸、乳酸脱氢酶和血尿素氮的影响
动物机体运动后会导致血乳酸浓度升高,堆积的血乳酸是导致运动性疲劳出现的重要因素[16],而乳酸脱氢酶的作用是催化乳酸氧化为丙酮酸从而减少乳酸的积累[17],运动后小鼠体内积累的血乳酸需要大量的乳酸脱氢酶进行转化,其指标与血乳酸指标负相关。血尿素氮作为机体蛋白质代谢的主要终末产物,是疲劳水平的重要标志,在正常情况下,尿素氮水平上升可能与高强度运动、肌肉损伤等因素有关,尿素氮的升高意味着体内蛋白质分解增加,肌肉组织受损,从而导致疲劳感加重[18−19]。由图2所示,与NC组对比,SAP-M组、SAP-H组和TP组的血乳酸均显著下降(P<0.05),分别降低了8.51%、21.74%和25.81%,SAP-L组虽有小幅度下降,但不具有显著性差异(P>0.05);由图3所示,与NC组对比,TP组和SAP-H组乳酸脱氢酶活性显著上升(P<0.05),分别升高了37.41%和41.34%,SAP-L、SAP-M组乳酸脱氢酶活性虽有缓慢上升效果,但不具有显著性差异(P>0.05),表明虾头自溶活性多肽可提高乳酸脱氢酶的活性,且高剂量组提升效果显著(P<0.05);由图4所示,小鼠游泳力竭运动后SAP-L、SAP-M和SAP-H组血尿素氮含量均低于NC组,且灌胃剂量越大,小鼠的疲劳程度越低,SAP-H组效果与TP组效果相当,均具有显著性差异(P<0.05)。
![]() 图 2 虾头自溶活性多肽对力竭游泳小鼠血乳酸含量的影响Figure 2. Effect of SAP on the blood lactic acid content of exhausted swimming mice
图 2 虾头自溶活性多肽对力竭游泳小鼠血乳酸含量的影响Figure 2. Effect of SAP on the blood lactic acid content of exhausted swimming mice![]() 图 3 虾头自溶活性多肽对力竭游泳小鼠乳酸脱氢酶活性的影响Figure 3. Effect of SAP on the activity of lactic acid dehydrogenase of exhausted swimming mice
图 3 虾头自溶活性多肽对力竭游泳小鼠乳酸脱氢酶活性的影响Figure 3. Effect of SAP on the activity of lactic acid dehydrogenase of exhausted swimming mice![]() 图 4 虾头自溶活性多肽对力竭游泳小鼠血尿素氮含量的影响Figure 4. Effect of SAP on the content of blood urea nitrogen of exhausted swimming mice
图 4 虾头自溶活性多肽对力竭游泳小鼠血尿素氮含量的影响Figure 4. Effect of SAP on the content of blood urea nitrogen of exhausted swimming mice2.4 虾头自溶活性多肽对力竭游泳小鼠肝脏还原型谷胱甘肽和丙二醛的影响
还原型谷胱甘肽参与体内氧化还原过程,通过与过氧化物及自由基结合,来对抗氧化剂对巯基的破坏,并且能够改善自由基对重要脏器的损害[19];丙二醛具有细胞毒性,是衡量氧化胁迫程度的常用指标之一,细胞受脂质氧化的损伤程度可以通过丙二醛的含量反映出来,游泳小鼠剧烈运动后,会产生大量的脂质过氧化产物丙二醛,导致机体氧化应激反应失衡,抗氧化能力减弱,引发机体产生疲劳[20−21]。由图5~图6可知,与NC组对比,SAP-L、SAP-M和SAP-H组和TP组还原型谷胱甘肽含量均有不同程度的升高,丙二醛的含量也有不同程度的降低。与NC组对比,SAP-M和SAP-H组和TP组的还原型谷胱甘肽含量均显著升高,具有显著性差异(P<0.05),各组分别升高23.36%、46.98%、41.47%;三个剂量组和阳性对照组的丙二醛含量均有所下降,与NC组对比,TP组、SAP-M和SAP-H组下降显著(P<0.05),三者分别降低了29.49%、31.10%、35.66%。上述实验结果表明,虾头自溶活性多肽具有较强的抗氧化能力,SAP-H组与TP组效果相当。
![]() 图 5 虾头自溶活性多肽对力竭游泳小鼠肝脏还原型谷胱甘肽的影响Figure 5. Effect of SAP on the liver reduced glutathione of exhausted swimming mice
图 5 虾头自溶活性多肽对力竭游泳小鼠肝脏还原型谷胱甘肽的影响Figure 5. Effect of SAP on the liver reduced glutathione of exhausted swimming mice![]() 图 6 虾头自溶活性多肽对力竭游泳小鼠肝脏丙二醛的影响Figure 6. Effect of SAP on the liver malondialdehyde of exhausted swimming mice
图 6 虾头自溶活性多肽对力竭游泳小鼠肝脏丙二醛的影响Figure 6. Effect of SAP on the liver malondialdehyde of exhausted swimming mice2.5 虾头自溶活性多肽对力竭游泳小鼠肝/肌糖原的影响
肌糖原和肝糖原主要存在于骨骼肌和肝脏中,作为机体血糖的储存方式,其合成和分解对机体血糖的平衡具有重要的作用[22]。由图7所示,在肝糖原方面,与NC组对比,SAP-L、SAP-M和SAP-H组和TP组肝糖原和肌糖原含量均显著升高(P<0.05),且随着SAP灌胃剂量的增加,肝糖原含量随之升高,二者呈现一定的量效关系;在肌糖原方面,与NC组对比,SAP-L、SAP-M和SAP-H组和TP组肌糖原含量均显著升高(P<0.05),但SAP-M、SAP-H和TP组三组之间无显著性差异(P>0.05)。
![]() 图 7 虾头自溶活性多肽对力竭游泳小鼠肝/肌糖原含量的影响Figure 7. Effect of SAP on the liver/muscle glycogen content of exhausted swimming mice
图 7 虾头自溶活性多肽对力竭游泳小鼠肝/肌糖原含量的影响Figure 7. Effect of SAP on the liver/muscle glycogen content of exhausted swimming mice2.6 虾头自溶活性多肽对力竭游泳小鼠肝脏切片组织的影响
虾头自溶活性多肽对力竭游泳小鼠肝脏组织形态学的影响见图8。NC组的小鼠肝脏细胞无空泡现象,细胞界限清晰,呈圆形的细胞核位于细胞中央,肝小叶结构清晰,胞浆均匀,细胞核形态正常。与NC组对比,TP组和SAP-L、SAP-M和SAP-H组肝脏细胞形态表现均为正常,细胞界限清晰,细胞核位于细胞中央,细胞密度适中且排列紧密,细胞间质较少,与空白组正常小鼠肝脏细胞并无差异,表明短期持续摄入适量虾头自溶活性多肽不影响小鼠的肝脏正常代谢功能,也不会对其肝脏造成损伤。
![]() 图 8 虾头自溶活性多肽对力竭游泳小鼠肝脏组织形态学的影响Figure 8. Effect of SAP on the histomorphology of the liver of exhausted swimming mice
图 8 虾头自溶活性多肽对力竭游泳小鼠肝脏组织形态学的影响Figure 8. Effect of SAP on the histomorphology of the liver of exhausted swimming mice2.7 虾头自溶活性多肽对力竭游泳小鼠抗疲劳相关基因mRNA表达的影响
Keap1/Nrf2/ARE信号通路在抗疲劳和抗氧化方面均具有重要的作用,该通路相关信号因子在器官中分布广泛,但在肝脏、肾脏以及肠胃中含量较高[23−25]。如图9所示,与NC组相比,TP组、SAP-M和SAP-H组中Nrf2、HO-1和NOQ-1 mRNA表达显著上调(P<0.05),SAP-L组HO-1和NOQ-1 mRNA表达虽有上调,但不具有显著差异性(P>0.05);与之相反,TP组和SAP-M和SAP-H组Keap1 mRNA表达显著下调(P<0.05),SAP-L组Keap1 mRNA表达虽有一定程度的下调,但无显著性差异(P>0.05)。
![]() 图 9 虾头自溶活性多肽对力竭游泳小鼠抗疲劳相关基因mRNA表达的影响Figure 9. Effect of SAP on the mRNA expression of anti-fatigue related genes in the liver of mice during exhaustive swimming
图 9 虾头自溶活性多肽对力竭游泳小鼠抗疲劳相关基因mRNA表达的影响Figure 9. Effect of SAP on the mRNA expression of anti-fatigue related genes in the liver of mice during exhaustive swimming3. 讨论
本实验结果表明,虾头自溶活性多肽可有效延长小鼠负重游泳力竭时间(图1),其延长效果与虾头自溶活性多肽剂量的摄入成正比,且能够显著提高小鼠血液中乳酸脱氢酶的活性(最高升高41.34%)(图3),降低其血乳酸和血尿素氮的含量(图2和图4),呈现出较好的抗疲劳效果,其中SAP-H组(600 mg/kg·d)与TP组(400 mg/kg·d)效果相当。其原因推测为虾头自溶活性多肽可以有效降低小鼠血乳酸浓度、抑制肌肉蛋白质分解、提高乳酸代谢水平[16−18],上述作用互作协同,使小鼠呈现抗疲劳特性,从而延缓小鼠疲劳的产生和加速疲劳的恢复。徐恺[26]通过南极磷虾肽抗疲劳、耐缺氧以及抗衰老实验研究发现,南极磷虾肽能够增加小鼠的存活时间、耗氧量和游泳时间,提高乳酸脱氢酶、肝糖原含量,具有显著的抗疲劳活性;王建永[27]研究报道表明,牡蛎多肽能够有效降低大鼠血乳酸和血尿素氮的含量,改善大鼠线粒体氧化应激水平,保护线粒体免受损伤,对运动性疲劳具有一定的缓解作用。由此,凡纳滨对虾自溶多肽与其他来源多肽一样,具有缓解运动疲劳的功能活性。
相关研究表明,补充肝/肌糖原可显著提高小鼠跑步耐力,可显著提高小鼠抗疲劳能力[28−29]。图5~图7结果显示,虾头自溶活性多肽高剂量组显著提高了小鼠肝脏组织中还原型谷胱甘肽的含量,降低了丙二醛含量,并提高其肝/肌糖原含量。上述与抗疲劳相关生化指标的显著改善,均表明虾头自溶活性多肽可通过减轻肝脏细胞的氧化应激损伤,减少小鼠体内抗氧化物质的产生,以及增加能量储备(肝/肌糖原)等途径来提高小鼠的运动能力和抗疲劳能力。
Keap1/Nrf2/ARE信号通路在抗疲劳方面具有重要作用,Nrf2属于转录因子Cap-n-collar家族,通过与抗氧化蛋白启动子的ARE(抗氧化反应元件)区域结合,调节抗氧化和抗炎反应[30−31]。本研究对游泳力竭小鼠体内Keap1/Nrf2/ARE信号通路中各信号因子mRNA表达进行检测,由图9可知,虾头自溶活性多肽中、高剂量组中Nrf2、HO-1和NQO-1 mRNA表达显著上调,Keap1 mRNA表达显著下调,表明虾头自溶活性多肽能激活Nrf2信号通路,发挥其抗疲劳、抗氧化作用。此结果与相关报道类似,如Chen等[32]在疲劳模型小鼠中发现,激活Keap1-Nrf2信号通路可以减轻疲劳程度;Sechang等[33]发现激活Nrf2可以显著减轻疲劳小鼠的肌肉损伤、氧化应激程度,并提高运动能力。
综上,本研究结果表明虾头自溶活性多肽可激活小鼠体内Keap1/Nrf2/ARE信号通路,减轻氧化应激、保护细胞免受损伤以及调节能量代谢,从而缓解其疲劳症状,具有显著的抗疲劳作用,但是否存在其他相关信号通路仍需要进一步研究。
4. 结论
本实验结果表明虾头自溶活性多肽对小鼠具有显著的抗疲劳作用,结合生化指标及Keap1/Nrf2/ARE信号通路相关信号因子的mRNA表达,虾头自溶活性多肽的抗疲劳机制可能主要通过调控Keap1 mRNA负表达,减弱其与Nrf2的相互作用,激活Nrf2信号通路,提高Nrf2 mRNA表达;作为Nrf2下游信号因子,HO-1和NQO-1受其调控mRNA表达显著上调,进而降低小鼠肝脏丙二醛、血乳酸、血尿素氮含量,提高乳酸脱氢酶、还原型谷胱甘肽、肝糖原含量,发挥其抗氧化、抗疲劳作用。本研究对湛江对虾加工副产物的综合利用进行了有益探索,研究结果对利用低值对虾加工副产物生产高附加值抗疲劳保健产品具有一定的技术参考价值。
-
![]()
![]()
图 2 虾头自溶活性多肽对力竭游泳小鼠血乳酸含量的影响
Figure 2. Effect of SAP on the blood lactic acid content of exhausted swimming mice
![]()
图 3 虾头自溶活性多肽对力竭游泳小鼠乳酸脱氢酶活性的影响
Figure 3. Effect of SAP on the activity of lactic acid dehydrogenase of exhausted swimming mice
![]()
图 4 虾头自溶活性多肽对力竭游泳小鼠血尿素氮含量的影响
Figure 4. Effect of SAP on the content of blood urea nitrogen of exhausted swimming mice
![]()
图 5 虾头自溶活性多肽对力竭游泳小鼠肝脏还原型谷胱甘肽的影响
Figure 5. Effect of SAP on the liver reduced glutathione of exhausted swimming mice
![]()
图 6 虾头自溶活性多肽对力竭游泳小鼠肝脏丙二醛的影响
Figure 6. Effect of SAP on the liver malondialdehyde of exhausted swimming mice
![]()
图 7 虾头自溶活性多肽对力竭游泳小鼠肝/肌糖原含量的影响
Figure 7. Effect of SAP on the liver/muscle glycogen content of exhausted swimming mice
![]()
图 8 虾头自溶活性多肽对力竭游泳小鼠肝脏组织形态学的影响
Figure 8. Effect of SAP on the histomorphology of the liver of exhausted swimming mice
![]()
图 9 虾头自溶活性多肽对力竭游泳小鼠抗疲劳相关基因mRNA表达的影响
Figure 9. Effect of SAP on the mRNA expression of anti-fatigue related genes in the liver of mice during exhaustive swimming
表 1 引物序列
Table 1 Primers sequence
基因 引物序列 引物长度(bp) 上游引物(5′~3′) 下游引物(5′~3′) Beta-actin GGCTGTATTCCCCTCCATCG CCAGTTGGTAACAATGCCATGT 154 Keap1 TCGAAGGCATCCACCCTAAG CTCGAACCACGCTGTCAATCT 135 Nrf2 TAGATGACCATGAGTCGCTTGC GCCAAACTTGCTCCATGTCC 153 HO-1 GATAGAGCGCAACAAGCAGAA CAGTGAGGCCCATACCAGAAG 111 NQO-1 AGGATGGGAGGTACTCGAATC AGGCGTCCTTCCTTATATGCTA 144  下载: 导出CSV
下载: 导出CSV
-
[1] BHAT Z. Bioactive peptides of animal origin:A review[J]. Journal of Food Science and Technology,2015,57(3):566−578.
[2] LIU R, LI Z, YU X C, et al. The effects of peanut oligopeptides on exercise-induced fatigue in mice and its underlying mechanism[J]. Nutrients,2023,15:1743. doi: 10.3390/nu15071743
[3] FENG L, YUE W, JIAN Y, et al. Overview of the preparation method, structure and function, and application of natural peptides and polypeptides[J]. Biomedicine & Pharmacotherapy,2022(153):113493.
[4] 左爱华, 马普, 刘心田, 等. 海参低聚肽对肾阳虚小鼠的影响[J]. 海洋湖沼通报,2021(1):77−84. [ZUO A H, MA P, LIU X T, et al. Anti-fatigue and aphrodisiac effects of a Postichopus japonicus oligopeptide on mice with kidney-yang deficiency[J]. Transactions of Oceanology and Limnology,2021(1):77−84.] ZUO A H, MA P, LIU X T, et al. Anti-fatigue and aphrodisiac effects of a Postichopus japonicus oligopeptide on mice with kidney-yang deficiency[J]. Transactions of Oceanology and Limnology, 2021(1): 77−84.
[5] 车帅, 刘楚怡, 王长伟, 等. 一种含有牡蛎肽保健酒的功能评价[J]. 中国酿造,2021,40(5):97−102. [CHE S, LIU C Y, WANG C W, et al. Functional assessment of a novel health care wine containing oyster peptide[J]. China Brewing,2021,40(5):97−102.] doi: 10.11882/j.issn.0254-5071.2021.05.018 CHE S, LIU C Y, WANG C W, et al. Functional assessment of a novel health care wine containing oyster peptide[J]. China Brewing, 2021, 40(5): 97−102. doi: 10.11882/j.issn.0254-5071.2021.05.018
[6] 骆贤亮, 晏永球, 冯凤琴. 牡蛎肽与人参提取物配伍提高雄性小鼠性功能及抗疲劳作用研究[J]. 食品工业科技,2022,43(1):366−374. [LUO X L, YAN Y Q, FENG F Q. Effect of combined oyster peptide and ginseng extracts on improvement of sexual function and anti-fatigue in male mice[J]. Science and Technology of Food Industry,2022,43(1):366−374.] LUO X L, YAN Y Q, FENG F Q. Effect of combined oyster peptide and ginseng extracts on improvement of sexual function and anti-fatigue in male mice[J]. Science and Technology of Food Industry, 2022, 43(1): 366−374.
[7] 朱国萍, 曹文红, 章超桦, 等. 凡纳滨对虾虾头自溶动力学[J]. 水产学报,2010,34(3):395−403. [ZHU G P, CAO W H, ZHANG C H, et al. Autolysis kinetics of Litopenaeus vannamei head[J]. Journal of Fisheries of China,2010,34(3):395−403.] doi: 10.3724/SP.J.1231.2010.06457 ZHU G P, CAO W H, ZHANG C H, et al. Autolysis kinetics of Litopenaeus vannamei head[J]. Journal of Fisheries of China, 2010, 34(3): 395−403. doi: 10.3724/SP.J.1231.2010.06457
[8] 朱国萍, 章超桦, 曹文红, 等. 超滤分离对虾头自溶产物ACE抑制活性的影响[J]. 上海海洋大学学报,2013,22(3):452−457. [ZHU G P, ZHANG C H, CAO W H, et al. Effect of ultrafiltration on ACE inhibitory activity of autolysate of shrimp head[J]. Journal of Shanghai Ocean University,2013,22(3):452−457.] ZHU G P, ZHANG C H, CAO W H, et al. Effect of ultrafiltration on ACE inhibitory activity of autolysate of shrimp head[J]. Journal of Shanghai Ocean University, 2013, 22(3): 452−457.
[9] 张风, 夏旭, 周爱梅, 等. 虾头虾壳蛋白质酶解制备抗氧化肽的研究[J]. 南方水产科学,2015,11(6):79−87. [ZHANG F, XIA X, ZHOU A M, et al. Research of antioxidant peptides produced from protein extracted from white shrimp head and shell[J]. South China Fisheries Science,2015,11(6):79−87.] doi: 10.3969/j.issn.2095-0780.2015.06.011 ZHANG F, XIA X, ZHOU A M, et al. Research of antioxidant peptides produced from protein extracted from white shrimp head and shell[J]. South China Fisheries Science, 2015, 11(6): 79−87. doi: 10.3969/j.issn.2095-0780.2015.06.011
[10] 王晋, 张风, 周爱梅, 等. 虾头、虾壳抗氧化肽的分离纯化及其对秀丽隐杆线虫的抗氧化作用[J]. 食品科学,2019,40(3):56−63. [WANG J, ZHANG F, ZHOU A M, et al. Purification of antioxidant peptides derived from enzymatic hydrolysates of shrimp heads and shells and their antioxidant protection in Caenorhabditis elegans[J]. Food Science,2019,40(3):56−63.] doi: 10.7506/spkx1002-6630-20180103-030 WANG J, ZHANG F, ZHOU A M, et al. Purification of antioxidant peptides derived from enzymatic hydrolysates of shrimp heads and shells and their antioxidant protection in Caenorhabditis elegans[J]. Food Science, 2019, 40(3): 56−63. doi: 10.7506/spkx1002-6630-20180103-030
[11] CAO W, ZHANG C, HONG P, et al. Response surface methodology for autolysis parameters optimization of shrimp head and amino acids released during autolysis[J]. Food Chemistry,2008,109(1):176−183. doi: 10.1016/j.foodchem.2007.11.080
[12] GUPTA S. Quantifying protein and nitrogen levels in Paramphistomum species[J]. Journal for Research in Applied Sciences and Biotechnology,2023,2(1):278−281. doi: 10.55544/jrasb.2.1.41
[13] 姜丽冬, 金鑫, 朴春红, 等. 林蛙油生物活性肽的制备工艺及其抗疲劳研究[J]. 吉林农业大学学报,2016,38(4):482−488. [JIANG L D, JIN X, PIAO C H, et al. Preparation technology and antifatigue effects of oviductus ranae bioactive peptide[J]. Journal of Jilin Agricultural University,2016,38(4):482−488.] JIANG L D, JIN X, PIAO C H, et al. Preparation technology and antifatigue effects of oviductus ranae bioactive peptide[J]. Journal of Jilin Agricultural University, 2016, 38(4): 482−488.
[14] 陈立艺, 李玉新, 黄倍源, 等. 参附注射液对小鼠抗缺氧抗疲劳作用的实验研究[J]. 中国中医急症,2015,24(6):960−961,978. [CHEN L Y, LI Y X, HUANG B Y, et al. Experimental study on anti-hypoxic and anti-fatigue effect of shenfu injection on mice[J]. Journal of Emergency in Traditional Chinese Medicine,2015,24(6):960−961,978.] doi: 10.3969/j.issn.1004-745X.2015.06.008 CHEN L Y, LI Y X, HUANG B Y, et al. Experimental study on anti-hypoxic and anti-fatigue effect of shenfu injection on mice[J]. Journal of Emergency in Traditional Chinese Medicine, 2015, 24(6): 960−961,978. doi: 10.3969/j.issn.1004-745X.2015.06.008
[15] CHAMBERLAND V, RIOUX P. Not only students can express alcohol dehydrogenase:Goldfish can too![J]. Advances in Physiology Education,2010,34(4):222−227. doi: 10.1152/advan.00088.2009
[16] RUSDIAWAN A, SHOLIKHAH A M, PRIHATININGSIH S. The changes in pH levels, blood lactic acid and fatigue index to anaerobic exercise on athlete after NaHCO3 administration[J]. Malaysian Journal of Medicine & Health Sciences,2020,16(supp16):50−56.
[17] ZHANG W, XIANG Q, ZHAO J, et al. Purification, structural elucidation and physicochemical properties of a polysaccharide from Abelmoschus esculentus L (okra) flowers[J]. International Journal of Biological Macromolecules,2020,155:740−750. doi: 10.1016/j.ijbiomac.2020.03.235
[18] KUMAR D, SINGH A K. Decoding the mechanism behind exercise-induced fatigue[J]. Neuroquantology,2022,20(15):5187−5124.
[19] FENG T, HUANG Y, TANG Z, et al. Anti-fatigue effects of pea (Pisum sativum L.) peptides prepared by compound protease[J]. Journal of Food Science and Technology,2021,58:2265−2272. doi: 10.1007/s13197-020-04737-3
[20] ZHANG L, MA Q, ZHOU Y. Strawberry leaf extract treatment alleviates cognitive impairment by activating Nrf2/HO-1 signaling in rats with streptozotocin-induced diabetes[J]. Frontiers in Aging Neuroscience,2020,12:201. doi: 10.3389/fnagi.2020.00201
[21] THORPE R T, ATKINSON G, DRUST B, et al. Monitoring fatigue status in elite team-sport athletes:Implications for practice[J]. International Journal of Sports Physiology and Performance,2017,12(S2):S227−S234. doi: 10.1123/ijspp.2016-0434
[22] GWYNETH S, MICHAEL T. The liver and glycogen:In sickness and in health[J]. International Journal of Molecular Sciences,2023,24(7):6133. doi: 10.3390/ijms24076133
[23] TU W, WANG H, LI S, et al. The anti-inflammatory and anti-oxidant mechanisms of the Keap1/Nrf2/ARE signaling pathway in chronic diseases[J]. Aging and Disease,2019,10(3):637−651. doi: 10.14336/AD.2018.0513
[24] MENG C L, JI J A, ZHENG Y J, et al. The Keap1–Nrf2–ARE pathway as a potential preventive and therapeutic target:An update[J]. Medicinal Research Reviews,2016,36(5):924−963. doi: 10.1002/med.21396
[25] SHI C, WANG X, LIU Y, et al. Effect and mechanism of banxia xiexintang on rats with chronic atrophic gastritis based on Keap1/Nrf2/ARE signaling pathway[J]. Chinese Journal of Experimental Traditional Medical Formulae,2021,24:31−37.
[26] 徐恺. 南极磷虾肽抗疲劳、耐缺氧以及抗衰老、提高免疫力实验研究[D]. 青岛:中国海洋大学, 2012: 24−39. [XU K. Experimental study of the functions of Antarctic krill peptide on fatigue resistance, anti-hypoxia, anti-aging and immunity[D]. Qingdao:Ocean University of China, 2012: 24−39.] XU K. Experimental study of the functions of Antarctic krill peptide on fatigue resistance, anti-hypoxia, anti-aging and immunity[D]. Qingdao: Ocean University of China, 2012: 24−39.
[27] 王建永. 牡蛎多肽对运动疲劳大鼠骨骼肌线粒体功能的影响[J]. 安徽大学学报(自然科学版),2020,44(5):93−99. [WANG J Y. Effects of oyster polypeptide on mitochondrial function of skeletal muscle in exercise induced fatigue rats[J]. Journal of Anhui University (Natural Science Edition),2020,44(5):93−99.] WANG J Y. Effects of oyster polypeptide on mitochondrial function of skeletal muscle in exercise induced fatigue rats[J]. Journal of Anhui University (Natural Science Edition), 2020, 44(5): 93−99.
[28] ILIANA L, GUINOVART J, DURAN J. Increased liver glycogen levels enhance exercise capacity in mice[J]. Journal of Biological Chemistry,2021,297(2):100976. doi: 10.1016/j.jbc.2021.100976
[29] ØRTENBLAD N, WESTERBLAD H, NIELSEN J. Muscle glycogen stores and fatigue[J]. The Journal of Physiology,2013,591(18):4405−4413. doi: 10.1113/jphysiol.2013.251629
[30] 熊款款, 谭磊, 王爱兵, 等. Keap1-Nrf2/ARE信号通路抗氧化机制及抗氧化剂的研究进展[J]. 动物医学进展,2021,42(4):89−94. [XIONG K K, TAN L, WANG A B, et al. Progress on anti-oxidation mechanisms and antioxidants of the Keap1-Nrf2/ARE signaling pathway[J]. Progress in Veterinary Medicine,2021,42(4):89−94.] XIONG K K, TAN L, WANG A B, et al. Progress on anti-oxidation mechanisms and antioxidants of the Keap1-Nrf2/ARE signaling pathway[J]. Progress in Veterinary Medicine, 2021, 42(4): 89−94.
[31] MA C, DENG Y, XIAO R, et al. Anti-fatigue effect of phlorizin on exhaustive exercise-induced oxidative injury mediated by Nrf2/ARE signaling pathway in mice[J]. European Journal of Pharmacology,2022,918:174563. doi: 10.1016/j.ejphar.2021.174563
[32] CHEN Y, WANG J, JING Z, et al. Anti-fatigue and anti-oxidant effects of curcumin supplementation in exhaustive swimming mice via Nrf2/Keap1 signal pathway[J]. Current Research in Food Science,2022,5:1148−1157. doi: 10.1016/j.crfs.2022.07.006
[33] SECHANG O, WARABI E, YAMAMOTO M, et al. Nrf2 activation remarkably improves exercise endurance capacity in mice[J]. Free Radical Biology and Medicine,2012,53(1):36−37.
 下载:
下载:















计量
- 文章访问数: 47
- HTML全文浏览量: 6
- PDF下载量: 7



