


Identification of A1/A2 β-Casein Genotypes and Comparison of Their Digestive Properties
-
摘要: A1 β-酪蛋白和A2 β-酪蛋白由于一级结构上的差异,消化后产生的物质也不相同。为了更全面地探究其对人体可能产生的生物活性影响,通过分析和监测该蛋白质在消化过程中的变化情况,进一步实现对A1/A2 β-酪蛋白生物功能的探索。在本研究中,对采自某一牧场不同奶牛的牛奶进行等位基因特异性PCR鉴别,并借助阴离子交换色谱和十二烷基硫酸钠聚丙烯酰胺凝胶电泳法对牛奶分离出的β-酪蛋白进行纯化和鉴定,最后构建静态体外消化模型比较β-酪蛋白在消化性能上的差异。结果显示,分离纯化的A1和A2 β-酪蛋白的浓度分别为0.62 mg/mL和0.66 mg/mL,纯度分别为95.28%和96.60%。两种β-酪蛋白的最终消化率均在90%左右,差异不显著。A2 β-酪蛋白自肠消化阶段开始直至消化结束表现出了更高的水解度,为25.34%。随着胃肠消化时间的增加,两种β-酪蛋白的粒径不断减小,电位绝对值不断增大,并且A2 β-酪蛋白在粒径和电位上的变化程度大于A1 β-酪蛋白。综上所述,本研究揭示了A1和A2 β-酪蛋白在静态体外消化过程中的理化特性差异,并在一定程度上表明A2 β-酪蛋白可能具有比A1 β-酪蛋白更好的消化性能,为进一步拓宽A2 β-酪蛋白在乳制品中的应用提供一定科学依据。
-
关键词:
- A1 β-酪蛋白 /
- A2 β-酪蛋白 /
- 等位基因特异性PCR /
- 体外模拟消化 /
- 消化性能
Abstract: A1 β-casein and A2 β-casein produce different substances after digestion due to differences in primary structure. To more comprehensively explore the possible biological activity effects on the human body, by analyzing and monitoring the changes of this protein during the digestion process, the exploration of the biological functions of A1/A2 β-casein is further achieved. In this study, allele-specific PCR was initially performed on milk samples collected from different cows in the pasture to identify A1 and A2 milk types. Afterwards, β-casein isolated from A1 and A2 milk was purified and characterized with the help of anion exchange chromatography and sodium dodecyl sulfate polyacrylamide gel electrophoresis. Finally, a static in vitro digestion model was constructed to compare A1 and A2 β-casein in terms of digestive properties. The results showed that the concentrations of isolated and purified A1 and A2 β-casein were 0.62 mg/mL and 0.66 mg/mL and the purities were 95.28% and 96.60%. After 5.0 h of static in vitro simulated digestion, the final digestibility of both β-caseins was around 90%, and the difference was not significant. A2 β-casein exhibited a higher degree of hydrolysis of 25.34% from the beginning of the intestinal digestion phase until the end of digestion. With the increase of gastrointestinal digestion time, the particle size of the two β-caseins decreased and the absolute value of the potential increased, and the degree of change in particle size and potential was greater for A2 β-casein than for A1 β-casein. In conclusion, the present study revealed the differences in the physicochemical properties of A1 and A2 β-caseins during static in vitro digestion and indicated to some extent that A2 β-caseins might have better digestive properties than A1 β-caseins, which provided a certain scientific basis for further expanding the application of A2 β-caseins in dairy products.-
Keywords:
- A1 β-casein /
- A2 β-casein /
- allele-specific PCR /
- in vitro mock digestion /
- digestive properties
-
β-酪蛋白是牛乳酪蛋白中四种单体之一,占牛乳酪蛋白总量的22%以上,且其具有较高的遗传变异性[1−2],由于其肽链中特定位点上氨基酸的不同,已被鉴定出A1、A2、A3、A4、B、C、D、E、F、G、H1、H2、I,共计13种类型,其中最常见的是A1和A2型[3]。A2型被认为是β-酪蛋白原始基因型,A1型则是基因突变的产物。随着自然基因突变的发生,部分A2型β-酪蛋白氨基酸肽链上的第67位氨基酸由脯氨酸(Pro)突变为组氨酸(His),成为A1型酪蛋白[4]。现有研究表明,亚洲地区本土的水牛等奶牛品种生产的牛乳大部分是A2型,但也存在少量A1型[5],而欧美及澳新地区的杂交奶牛如荷斯坦牛、娟姗牛、丹麦红牛、爱尔夏牛等品种则普遍生产A1牛奶。当前陆续出现了有关A1 β-酪蛋白经人体消化后,产生更高含量的β-酪啡肽-7(β-Casomorphin-7,BCM7)这种具有类似阿片类药物活性的肽的报道,并且BCM7可能通过与部分阿片受体结合导致消化不良[6]、1型糖尿病[7]等症状,而A2 β-酪蛋白则凭借BCM7的低释放量,以及对乳糖不耐症人群消化友好等优点,成为潜在的有益活性物质。动物实验研究表明A2 β-酪蛋白具有改善宿主肠道微生物群失调和缓解免疫功能下降等方面的功能[8]。为了更全面地探究A2 β-酪蛋白对人体可能产生的生物活性影响,有必要系统地分析和监测该蛋白质在人体消化过程中的变化情况,实现对A2 β-酪蛋白生物功能的进一步探索。
BCM7作为一种在A1和A2 β-酪蛋白中差异释放的具有一定促炎作用的肽段[9],逐渐成为研究这两种酪蛋白消化过程差异的有效切入点。Cattaneo等[10]通过体外静态消化A1和A2牛奶,发现A1 β-酪蛋白更容易释放BCM7,可能会对人体消化系统产生不良影响。同时,对A1和A2 β-酪蛋白在人胃肠道转运过程中差异的研究也成为一种思路。在一项研究中,与仅含有A2 β-酪蛋白的牛奶相比,含有A1和A2 β-酪蛋白的牛奶在胃中的转运速度更快,被认为可能会引起人体的乳糖不耐受症状[11]。另外,在对含有A1 β-酪蛋白和仅含有A2 β-酪蛋白奶粉的消化过程中,研究人员发现两种奶粉在胃蛋白酶分解水平上存在显著差异,表明这两种酪蛋白可能在体内胃消化和排空中发挥重要作用[12]。最近,Meng等[13]还评估了仅含有A2 β-酪蛋白的成长奶对幼儿胃肠道耐受性的有效性,发现与食用传统牛奶相比,食用A2 β-酪蛋白的孩子便秘较少,能够改善幼儿的整体消化舒适度和胃肠道相关症状。然而,大部分研究人员只是直接获得了相应的A2产品,并没有对产品实际基因型进行确认,因此可能会导致结果出现一定偏差。
本研究以牧场采集牛奶为研究对象,利用等位基因特异性PCR法区分样品源奶牛的基因型,将A1和A2牛奶分离纯化得到的A1和A2 β-酪蛋白,通过比较二者在体外消化上的差异,进而探究A2牛奶在人体胃肠道中发挥的积极作用,旨在为A2 β-酪蛋白在消化特性研究、提高生物利用度、拓宽食品应用等方面提供理论基础,为人们选择更健康的乳制品提供科学依据。
1. 材料与方法
1.1 材料与仪器
原料奶 河北某牧场4岁澳洲进口纯种荷斯坦奶牛和娟姗牛,每头奶牛取500 mL;琼脂糖、三氟乙酸、PBS缓冲液、磺基水杨酸、邻苯二甲醛、四硼酸钠、十二烷基硫酸钠、考马斯亮蓝R-250 美国Sigma公司;胰蛋白酶(100 U/mg)、胃蛋白酶(3000 U/mg) 北京索莱宝科技有限公司;蛋白预染Marker、5×蛋白上样缓冲液 赛默飞世尔科技有限公司;SDS-PAGE试剂盒 上海雅酶生物医药科技有限公司;BCA蛋白浓度试剂盒 安徽白鲨生物科技有限公司;其他化学试剂均为分析纯。
CF 1542 R小型台式高速冷冻离心机 德国Sigma有限公司;PILOT 10-15 M制备型冷冻干燥机 北京博医康实验仪器有限公司/中国;PP-1105电泳仪、MP-8120转移电泳仪槽、MP-3030垂直电泳槽 北京凯元信瑞仪器有限公司;NANO TRAC WAVE II纳米粒度及Zeta电位分析仪 美国MICROTRAC(麦奇可)公司;DU800紫外可见分光光度计 美国贝克曼库尔特有限公司;ID 3全自动酶标仪 美谷分子仪器(上海)有限公司;LAS 500凝胶成像仪、Pure M蛋白纯化分析系统 美国GE公司;GM-05 PCR基因扩增仪 力新仪器(上海)有限公司。
1.2 实验方法
1.2.1 基因型的鉴定与分型
参考张园园[14]的方法,并在此基础上进行修改,利用等位基因特异性PCR法对采集牛奶进行基因型鉴定。10组原料奶进行编号,设置两个单独的反应引物,CSNA1F和CSNAR用于扩增244 bp A1变体(CCA-组氨酸),CSNA2F和CSNAR用于扩增244 bp A2变体,正向引物:CSNAR(5’-GCCCAGATGAGAGAAGTGAGG-3’),反向引物:CSNA1F(5’-GATGTTTTGTGGGAGGCTGTTAT-3’),CSNA2F(5’-GATGTTTTGTGGGAGGCTGTTAG-3’),反应体系20 μL:50 mg DNA模板,10 μL Premix TaqTM,正向和反向引物各0.8 μL,ddH2O至20 μL,程序如下:95 ℃预变性5 min,95 ℃变性15 s,58 ℃退火15 s,72 ℃延伸60 s,35个循环后72 ℃延伸5 min。扩增后的PCR产物用2%,120 V琼脂糖凝胶电泳检测40 min并进行基因分型,以备后续实验使用。
1.2.2 A1和A2 β-酪蛋白的分离纯化
1.2.2.1 β-酪蛋白的粗提取
分别取A1和A2各100 mL乳样,4 ℃下5000 r/min离心25 min,除去牛乳中大部分乳脂,利用1 mol/L盐酸将牛乳的pH调至4.6,静置35 min,将获取的酪蛋白进行中性洗涤处理后,选用截留分子量范围在8000~14000 Da的透析袋,在2~5 ℃的低温环境下透析48 h。期间每隔7~8 h及时更换新鲜的透析液。待透析步骤全部完成后,采用冻干机对酪蛋白溶液进行冷冻干燥处理,最后将其储存在−20 ℃的低温环境中以确保其稳定性。
1.2.2.2 β-酪蛋白的离子交换色谱分离
在刘纳[15]的基础上,对β-酪蛋白的阴离子交换色谱法进行改进。流动相B由1 mol/L氯化钠溶于流动相A组成。将粗提取得到的酪蛋白溶解在流动相A中,调至浓度为10 mg/mL的样品溶液。0.22 μm滤膜过滤以实现除菌处理,最后完成上样步骤。梯度洗脱条件参照鲁杏茹[16]的方法。
1.2.2.3 SDS-PAGE鉴定色谱分离峰
参照赵烜影[17]的方法,运用十二烷基硫酸钠聚丙烯酰胺凝胶电泳(Sodium Dodecyl Sulfate Polyacrylamide Gel Electropheresis,SDS-PAGE)技术手段,按照SDS-PAGE试剂盒说明书制胶,配制浓度为15%的分离胶和5%的浓缩胶,待浓缩胶凝固后,拔去梳子准备加样。收集液和β-酪蛋白标准溶液各取10 μL并分别与5×蛋白上样缓冲液等体积混合,沸水浴加热3~5 min,在4 ℃,12000 r/min条件下离心2 min后上样,上样体积为10 μL,电泳初始电压为80 V,约1 h溴酚蓝指示线到达分离胶后调节至120 V。电泳结束后,染色50 min,并根据凝胶本身的脱色情况决定脱色时间。之后进行分析与鉴定工作,以准确辨识蛋白质的类别及其纯度水平。
1.2.2.4 分离产物的脱盐及浓度测定
添加相当于样品溶液体积4倍的超纯水,以置换并去除收集液中的盐离子成分,随后将溶液浓缩至20 mL,并将该溶液冷冻干燥处理,于−20 ℃下保存。将A1和A2 β-酪蛋白分别溶解在PBS缓冲液之中,配制成终浓度为1 mg/mL的溶液。用BCA蛋白浓度测定试剂盒对溶解后的β-酪蛋白溶液进行蛋白浓度的测定。
1.2.3 A1和A2 β-酪蛋白消化产物特性
1.2.3.1 消化产物的制备
参照Egger等[18]的方法并进行调整,静态体外消化A1和A2 β-酪蛋白。将10 mL 4%的A1和A2 β-酪蛋白溶液与10 mL含有胃蛋白酶(3000 U/mL)的模拟胃液等体积混合,置于培养箱(37 ℃,130 r/min)中,分别在消化的30、60、90和120 min取出,最大胃期终点为120 min。完成胃期后,从胃相取20 mL消化液与20 mL含有胰蛋白酶(100 U/mL)和胆盐(10 mmol/L)的模拟肠液混合,并在消化的150、180、210、240和300 min取出。通过添加NaOH抑制胃期收集样品中的胃蛋白酶活性,直到pH调至7.0,煮沸冷却后放置在−20 ℃冻存备用。肠期样品收集方式与胃期相同。
1.2.3.2 消化率的测定
取出已进行消化反应的消化液,通过加入0.1 mol/L盐酸溶液调节pH至4.6,在1400 r/min下离心2 min,将上清液弃去。使用PBS缓冲液将离心后得到的沉淀物重新溶解。利用BCA蛋白浓度测定试剂盒来测定溶解后溶液中总蛋白质的质量浓度(μg/mL)。按照下式计算β-酪蛋白体外消化率:
β-酪蛋白体外消化率(%)=P0−P1P0×100 (1) 式中:P0是模拟消化前状态下蛋白质样品中的初始蛋白质浓度,μg/mL;P1是经过消化处理后蛋白质样品中剩余的蛋白质浓度,μg/mL。
1.2.3.3 水解度的测定
静态体外消化后,使用邻苯二甲醛(OPA)方法定量上清液中的游离氨基,确定蛋白质水解的程度,参照鲁杏茹[16]的方法并按照下式计算水解度:
水解度(%)=SerineNH2−βα×htot×100 (2) SerineNH2=OD2−OD0OD1−OD0×0.9516×N×VX×P (3) 式中:Serine NH2是每克蛋白质中丝氨酸氨基的含量,mmol/g;β和α分别是常数0.4和1.0;htot是酪蛋白肽键总数,为8.2;OD0、OD1和OD2分别是空白组(去离子水)、标准品和样品的吸光度;0.9516是丝氨酸标准溶液浓度,meqv/L;N是稀释倍数;V是上清液体积,L;X是样品质量,g;P是样品中蛋白质含量,%。
1.2.3.4 粒径与电位的测定
采用Nano ZS马尔文纳米粒度电位分析仪,对β-酪蛋白静态体外消化后的粒径和电位变化情况进行分析。粒径测定中参照何思宇[19]的方法并稍作调整,将1 mL的消化液以50倍的比例用超纯水稀释,取出1 mL稀释后的液体注入到比色皿中,静置2 min。采用动态光散射模式对其进行测量,设置的参数条件如下:采用光源功率为4 mW的He-Ne激光器,分散介质的折射率为1.330,实验温度恒定在25 ℃,样本测试容器选用10 mm光程长度的石英材质比色皿。电位测定中参照田浩[20]的方法并稍作调整,取100 µL的消化液,然后用0.01 mol/L、pH为7.3的磷酸盐缓冲液对其作1000倍的稀释处理,充分混匀后,使用一次性移液器将样品溶液缓慢且平稳地注入样品池内。为了防止因气泡形成而引入实验误差,务必在注入样品后用盖子严密封闭样品池,确保无气泡产生。
1.2.3.5 分子量的测定
利用SDS-PAGE凝胶电泳法对A1和A2 β-酪蛋白在0、30、60、90、120、150、180、210、240和300 min的消化液进行分析,并对二者在体外消化性能上的差异进行比较。电泳操作步骤同1.2.2.3。
1.3 数据处理
研究中采用了SPSS 27.0.1版本软件实施单变量方差分析(ANOVA),同时运用PeakFit v4.12、Microsoft Excel 2018以及GraphPad Prism 8.02软件进行深入的统计处理及可视化图表制作。实验设计中,所有测试均执行了三个平行重复,以“平均值±标准差(mean±SD)”的形式表示测定结果。P<0.05表示结果具有显著差异。
2. 结果与分析
2.1 原料奶的基因分型
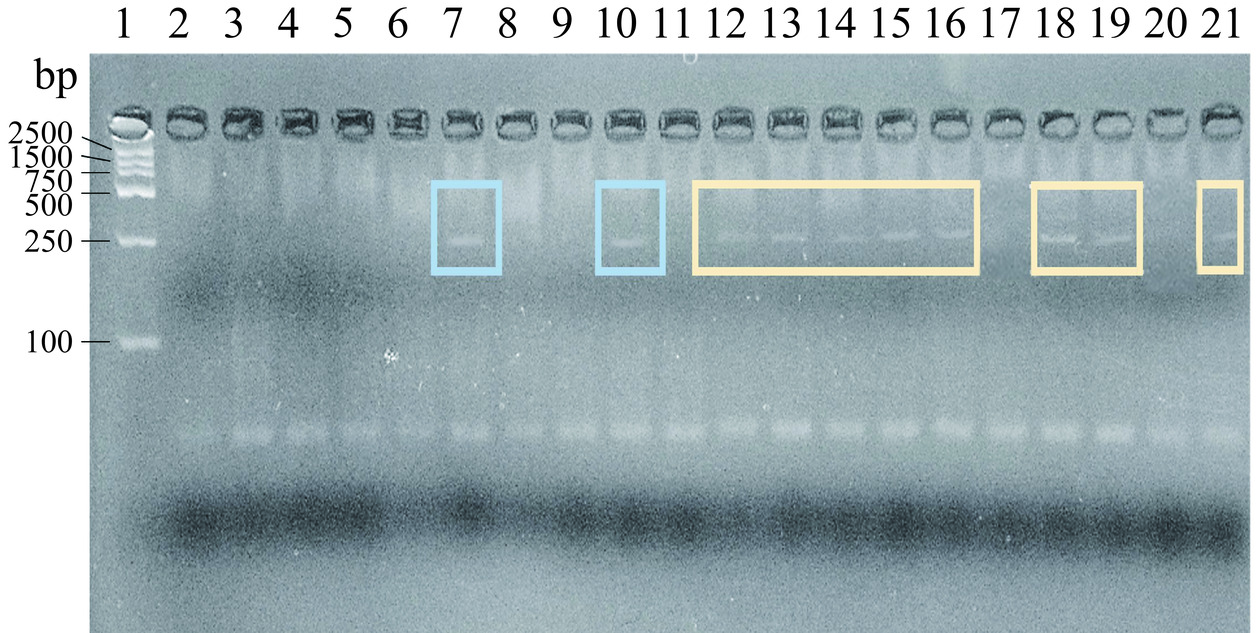
Kulibaba等[21]对A1和A2β-酪蛋白采用等位基因特异性PCR法进行检测,获得了相应244 bp的DNA扩增片段,本实验也获得了对应分子量的条带,β-酪蛋白基因部分PCR产物的琼脂糖凝胶电泳图谱如图1所示,在A1特异性引物扩增结果中,7与10泳道出现清晰条带,17与20泳道并未出现条带,故鉴定7和10泳道的样品为A1基因型,如图1中蓝框所示,且7泳道条带相比10泳道更为清晰,故选为后续A1的实验样品;在A2特异性引物扩增结果中,12、13、14、15、16、18、19、21泳道出现清晰条带,2、3、4、5、6、8、9、11泳道并未出现条带,故鉴定12、13、14、15、16、18、19、21泳道的样品为A2基因型,如图1中黄框所示,且其中18泳道条带最为清晰,故选为后续A2的实验样品。Rahimi等[22]在研究β-酪蛋白基因分型时也采用了此种方法,并初步获得了其所调查地区牛群的基因频率情况,发现该地区β-酪蛋白A1等位基因的频率相对较高,但因本研究调查样本较小,所以暂时无法体现出采样地区牛群的基因频率情况。
![]() 图 1 244 bp β-酪蛋白基因型AS-PCR产物的琼脂糖凝胶电泳图谱注:泳道1为标准分子量Marker(100~2000 bp);泳道2~11为A1特异性引物扩增结果;泳道12~21为A2特异性引物扩增结果。Figure 1. Agarose gel electrophoresis pattern of AS-PCR products of β-casein genotypes with 244 bp
图 1 244 bp β-酪蛋白基因型AS-PCR产物的琼脂糖凝胶电泳图谱注:泳道1为标准分子量Marker(100~2000 bp);泳道2~11为A1特异性引物扩增结果;泳道12~21为A2特异性引物扩增结果。Figure 1. Agarose gel electrophoresis pattern of AS-PCR products of β-casein genotypes with 244 bp2.2 A1和A2 β-酪蛋白的分离纯化
2.2.1 β-酪蛋白的离子交换色谱分离
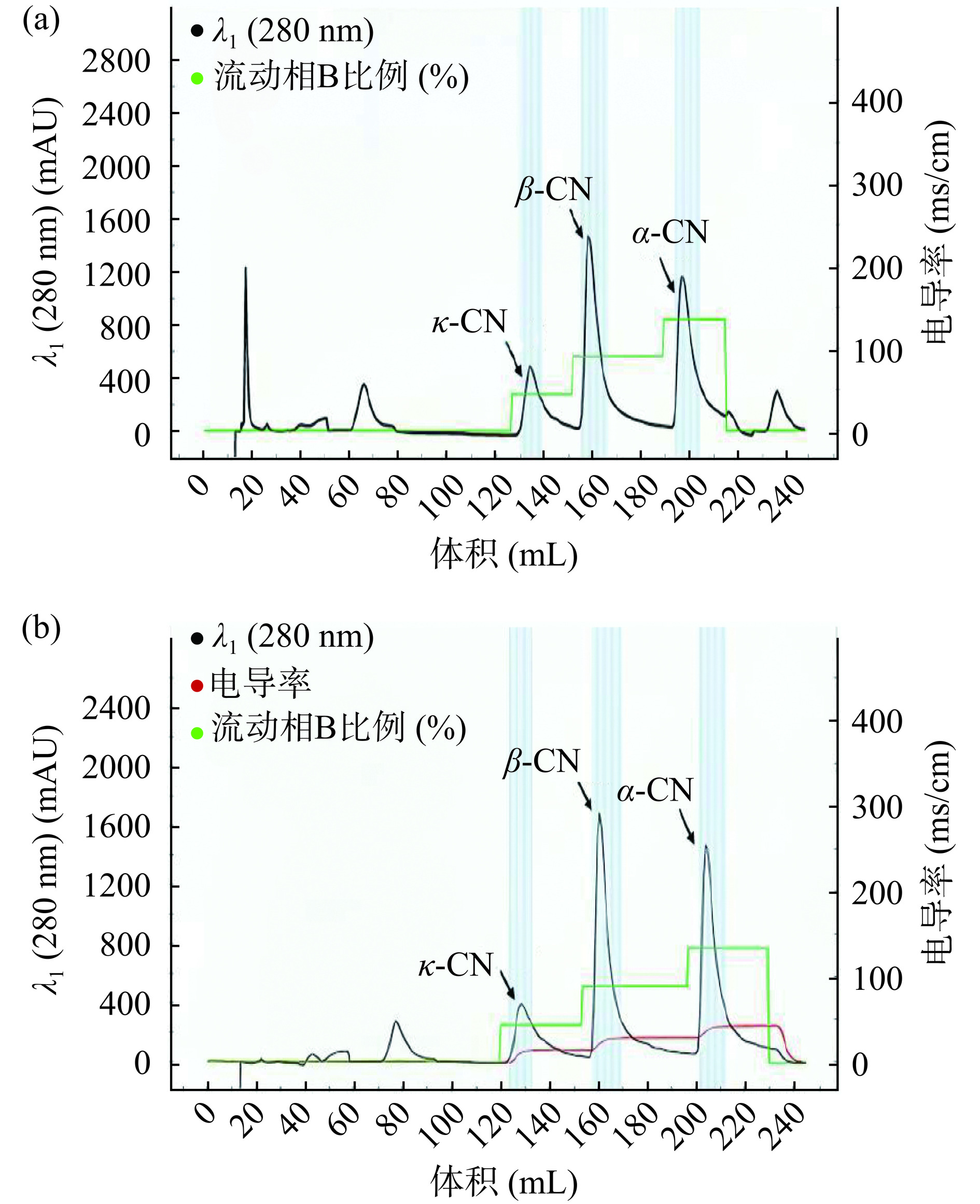
以A2牛奶为例,通过等电点沉淀法对酪蛋白进行粗提,并对其进行了分离纯化,结果如图2所示。利用阴离子交换色谱柱进行蛋白梯度洗脱。酪蛋白可分为αS1-酪蛋白、αS2-酪蛋白、β-酪蛋白和κ-酪蛋白,在牛乳中含量分别为12~15、3~4、9~11和3~4 mg/mL,分子量分别为23.6、25.2、24.0和19.0 kDa,等电点分别为4.9~5.0、5.2~5.4、5.1~5.4和5.4~5.6[23]。鉴于α-酪蛋白在酪蛋白组分中具有最低的等电点范围(4.9~5.4),同时其含量相对较高,所以相比其他酪蛋白,α-酪蛋白在缓冲液中表现出更大的净电荷量。基于此特性,在离子强度较低的情况下,α-酪蛋白不易从体系中分离出来,因此最后出峰。同理,最先出峰的为κ-酪蛋白,中间峰为β-酪蛋白,这与β-酪蛋白标准品的出峰时间类似,与Thekkilaveedu等[24]和Li等[25]的研究结果相符。为进一步鉴定β-酪蛋白并提高实验精确度,对收集的中间峰的洗脱液进行SDS-PAGE鉴定。
![]() 图 2 阴离子交换色谱柱分离A1(a)和A2(b)酪蛋白层析图注:α-CN、β-CN、κ-CN分别表示α-酪蛋白、β-酪蛋白、κ-酪蛋白。Figure 2. Separation of A1 (a) and A2 (b) casein chromatograms on an anion-exchange column
图 2 阴离子交换色谱柱分离A1(a)和A2(b)酪蛋白层析图注:α-CN、β-CN、κ-CN分别表示α-酪蛋白、β-酪蛋白、κ-酪蛋白。Figure 2. Separation of A1 (a) and A2 (b) casein chromatograms on an anion-exchange column2.2.2 色谱分离峰的SDS-PAGE鉴定
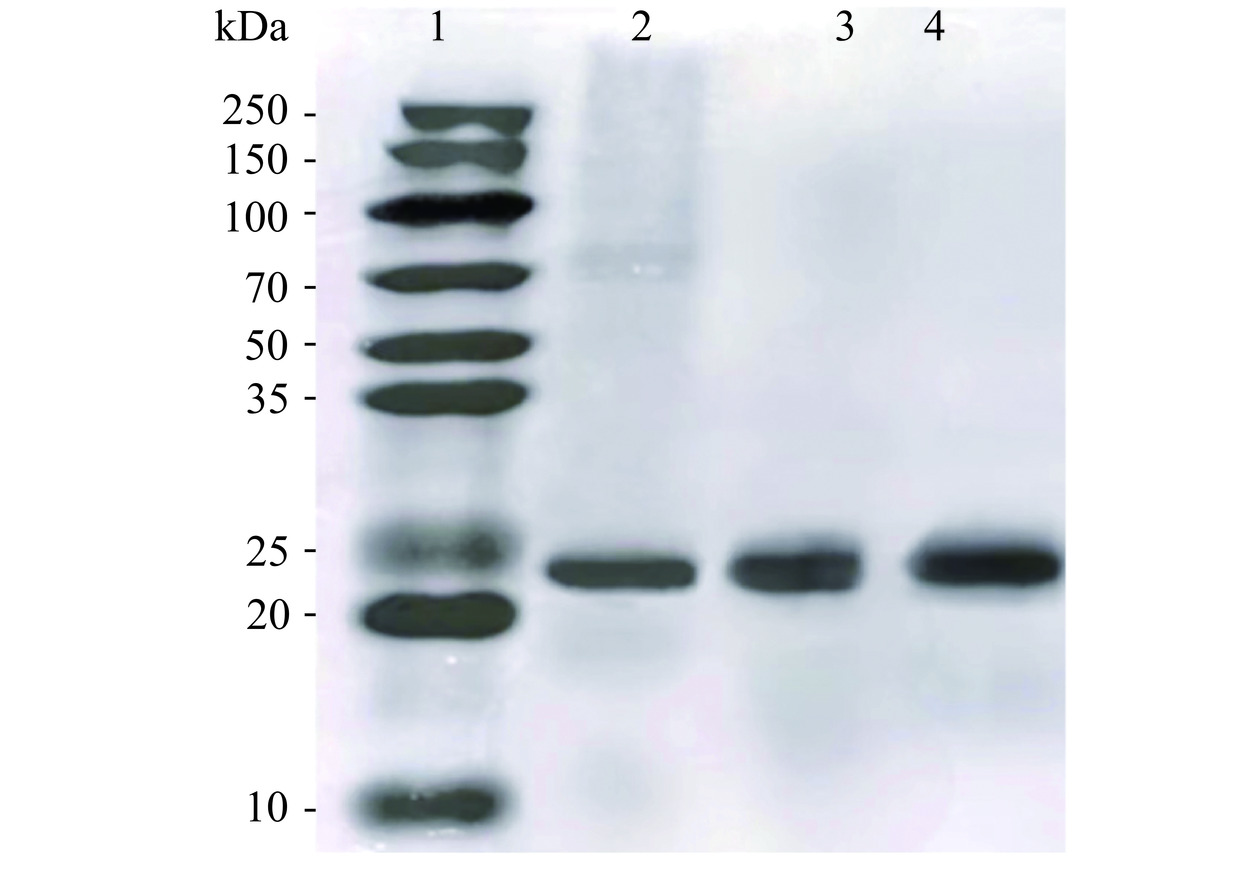
取中间峰的洗脱液进行SDS-PAGE电泳验证,采用考马斯亮蓝进行染色,结果如图3所示,将泳道3、4条带所处位置与泳道1进行对比可以得出,样品的分子量约在22~25 kDa,而β-酪蛋白的平均分子量在23.6 kDa[26],因此,可以推测3、4泳道样品为β-酪蛋白,这与阴离子交换层析图谱的分析结果一致,也与Okawara等[27]的电泳实验结果相符。图中可见洗脱出来的蛋白质较为单一,且纯度较高,说明洗脱效果较好,能够满足实验要求,但由于A1、A2 β-酪蛋白分子量差距较小,两种酪蛋白的质量分数相差不到千分之二,所以在鉴定时发现二者泳道条带无明显差别。
![]() 图 3 A1与A2 β-酪蛋白分离峰的洗脱液SDS-PAGE图谱注:泳道1:标准分子量Marker;泳道2:β-酪蛋白标准品;泳道3:A1 β-酪蛋白样品;泳道4:A2 β-酪蛋白样品。Figure 3. SDS-PAGE spectrum of eluates from A1 and A2 casein separated peaks
图 3 A1与A2 β-酪蛋白分离峰的洗脱液SDS-PAGE图谱注:泳道1:标准分子量Marker;泳道2:β-酪蛋白标准品;泳道3:A1 β-酪蛋白样品;泳道4:A2 β-酪蛋白样品。Figure 3. SDS-PAGE spectrum of eluates from A1 and A2 casein separated peaks2.2.3 A1和A2 β-酪蛋白纯度及浓度分析
按照BCA蛋白浓度试剂盒的方法得出蛋白浓度标准曲线y=0.4988x+0.1148,R2=0.9999,可以计算A1 β-酪蛋白的浓度为0.62 mg/mL,A2 β-酪蛋白的浓度为0.66 mg/mL,由Quantity One软件推算A1和A2 β-酪蛋白的纯度分别为95.28%和96.60%,这说明采用透析法对蛋白质进行离子置换和浓缩,可以对从离子交换层析中得到的酪蛋白进行高效的脱盐提纯。
2.3 A1和A2 β-酪蛋白消化产物特性
2.3.1 体外消化率测定
消化率是反映蛋白质在胃肠中消化水平的直观指标。本研究通过静态体外消化A1和A2 β-酪蛋白,分析两种β-酪蛋白的消化特性。两种β-酪蛋白的消化情况如图4所示,随着A1和A2 β-酪蛋白在胃肠模拟消化液中孵育时间的增加,消化率显著上升(P<0.05),表明这两种酪蛋白在胃蛋白酶和胰蛋白酶的作用下发生水解,形成小分子肽和游离氨基酸等。徐一宁等[28]在研究肌原纤维蛋白体外消化特性的过程中也得到了相似的结论。此外,从消化开始直至结束,A2 β-酪蛋白的消化率一直高于A1 β-酪蛋白,在消化结束时,A2 β-酪蛋白消化率为92.82%,A1 β-酪蛋白消化率为89.62%。两种β-酪蛋白的最终消化率均在90%左右,表明β-酪蛋白基本在肠中被完全消化,且无显著差异(P>0.05)。
![]()
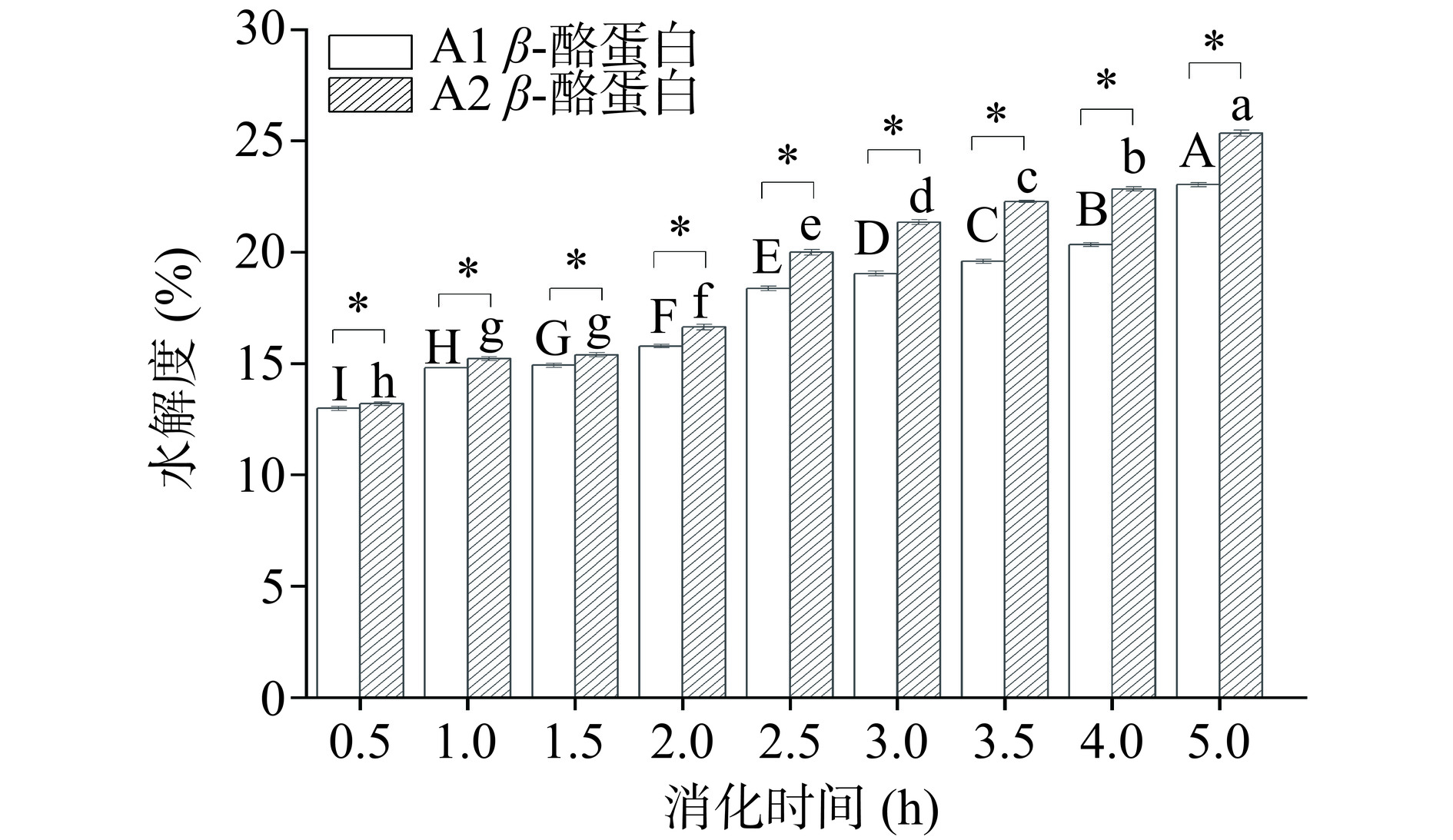
2.3.2 水解度变化
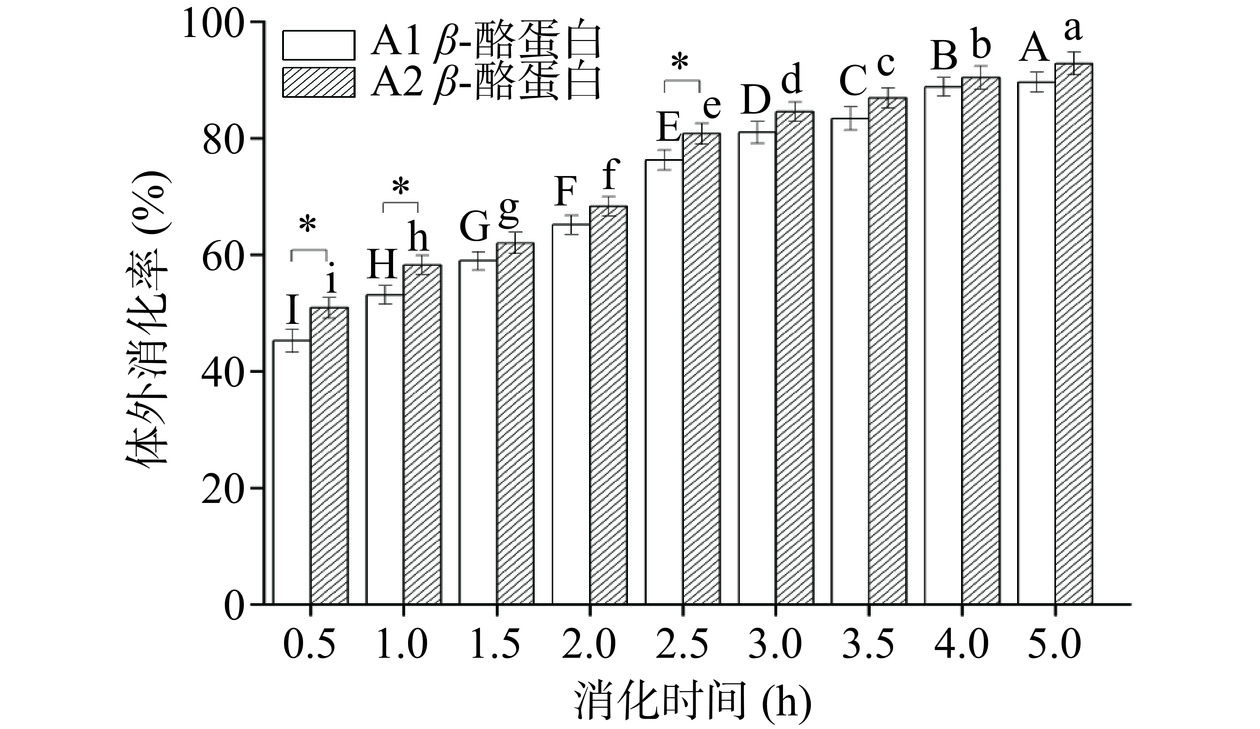
水解度作为评价蛋白体外消化的重要指标之一,其原理是通过测定消化液中的游离氨基量反映出消化过程中肽键的水解情况[28]。结果如图5所示。A1和A2 β-酪蛋白的水解度随着消化时间的增加而显著增加(P<0.05),但A1 β-酪蛋白在胃消化阶段(0~2 h)水解度变化不大,进入肠消化阶段(2~5 h)后水解度显著增加(P<0.05),有研究表明在胰蛋白酶水解过程中,β-酪蛋白多肽链的疏水区域不会被强烈水解[29],这可能意味着A1 β-酪蛋白多肽链具有更多的疏水区域从而导致了其相对A2 β-酪蛋白低的水解度。结合2.3.1的测定结果,两种β-酪蛋白在肠期基本完成水解,但即使A2 β-酪蛋白表现出较高的水解度,也仅为25.34%,说明依旧存在许多长肽链未被水解,这些长肽链将延缓人体肠道的吸收速率,这也表明单一的消化率不足以说明蛋白质的消化特性。同时,两种酪蛋白在水解度上的差异要稍大于消化率,这可能意味着A2 β-酪蛋白会产生更多小分子肽,这些肽可能发挥一定的有益作用。此外,Vorob’ev等[30]还发现不同基因型的β-酪蛋白经体外消化后衍生的同种肽在丰度上存在显著差异,这也可能是导致A1和A2 β-酪蛋白在人体中表现出功能差异性的原因之一。
![]() 图 5 A1和A2 β-酪蛋白体外水解度变化Figure 5. Changes of in vitro hydrolysis degree of A1 and A2 β-casein
图 5 A1和A2 β-酪蛋白体外水解度变化Figure 5. Changes of in vitro hydrolysis degree of A1 and A2 β-casein2.3.3 消化物粒径和电位的变化
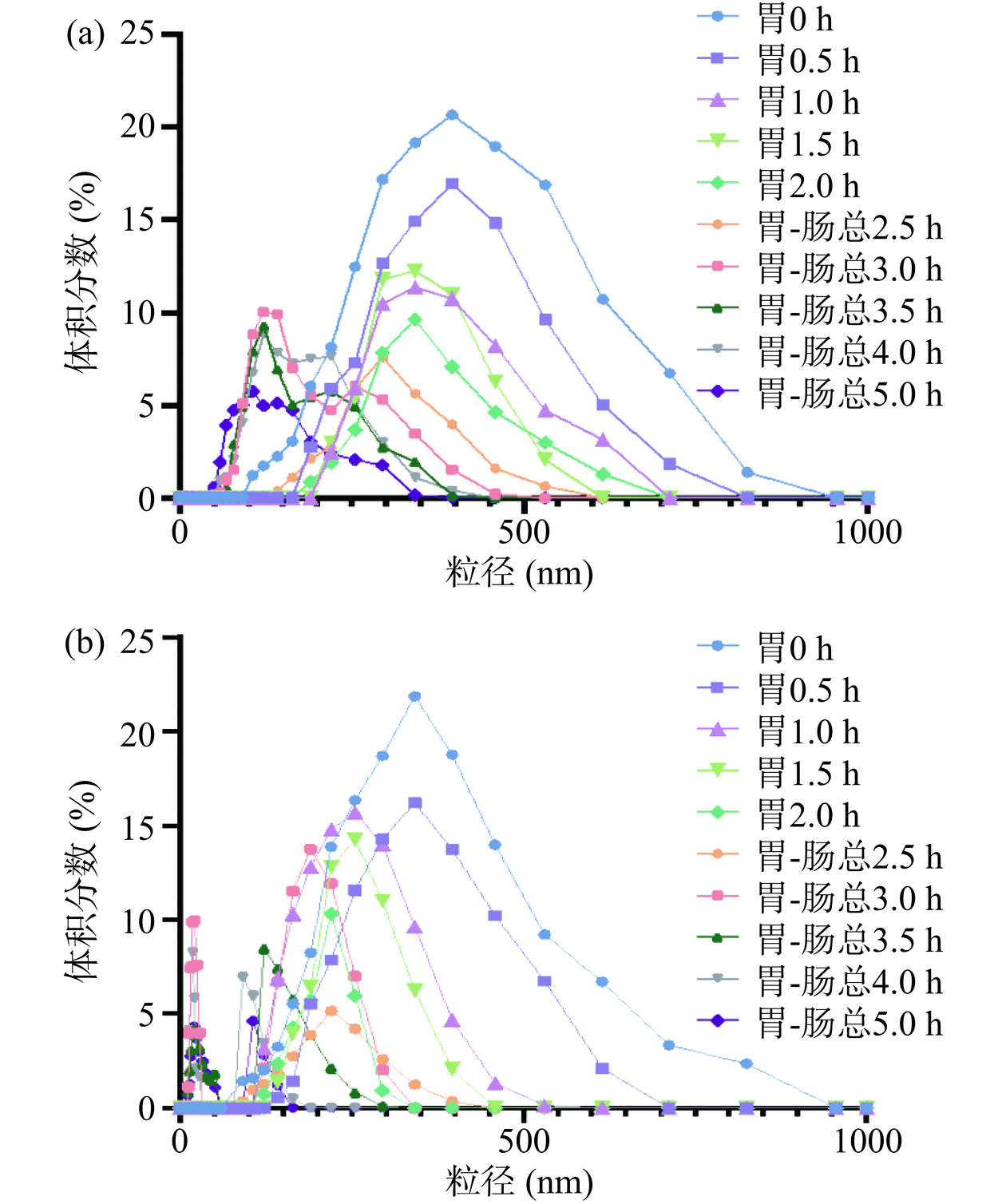
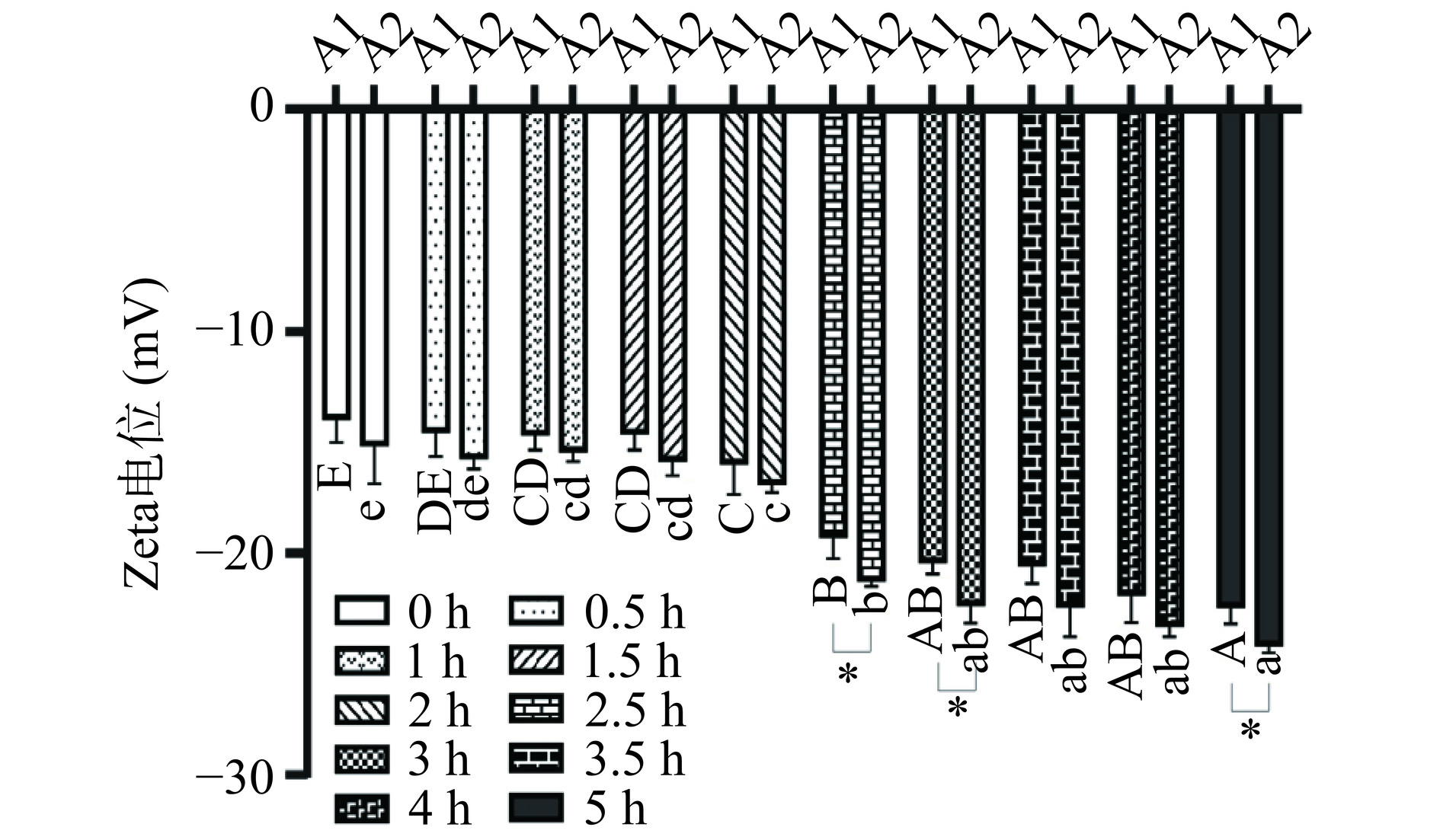
粒径的大小及分布反映了蛋白质在消化过程中被分解的程度。A1和A2 β-酪蛋白在静态体外消化过程中粒径的大小及分布如图6所示。β-酪蛋白粒径主要集中在150~850 nm之间,随着消化过程的进行,β-酪蛋白在胃蛋白酶和胰蛋白酶的作用下粒径不断减小,发生不同程度的水解,产生许多具有ACE抑制活性[31]、抗菌活性[32]、抗癌活性[33]等生物活性的肽以及游离氨基酸。在胃消化期间,A2 β-酪蛋白粒径峰值(由350 nm变为225 nm)的变化程度要大于A1 β-酪蛋白(400 nm变为350 nm)。进入肠消化阶段,A1和A2 β-酪蛋白粒径分布曲线均出现了两个峰值,但A2 β-酪蛋白更为明显,分别在50 nm和150 nm附近,随着时间的延长,峰值不断向左移动。相比之下,A1 β-酪蛋白的峰值最终停留的位置要相对A2 β-酪蛋白右移,这说明A1 β-酪蛋白的水解程度要弱于A2 β-酪蛋白,这也印证了2.3.2的结论。电位绝对值的大小反映出乳液体系的稳定性[34],如图7所示,随着消化的进行,A1和A2 β-酪蛋白电位绝对值不断增大,表明其粒子表面负电荷不断增多,粒子之间排斥增大,形成的乳液体系趋于稳定,且最终A2 β-酪蛋白电位绝对值显著大于A1 β-酪蛋白(P<0.05),说明A2 β-酪蛋白产生了相对多的带电荷氨基酸,即发生水解程度更大一些,这与粒径实验获得的结果相一致。
![]() 图 6 A1(a)和A2(b)β-酪蛋白静态体外消化粒径分布Figure 6. Particle size distribution of A1 (a) and A2 (b) β-casein in static in vitro digestion
图 6 A1(a)和A2(b)β-酪蛋白静态体外消化粒径分布Figure 6. Particle size distribution of A1 (a) and A2 (b) β-casein in static in vitro digestion![]() 图 7 A1和A2 β-酪蛋白静态体外消化Zeta电位图Figure 7. Zeta potential of A1 and A2 β-casein in static in vitro digestion
图 7 A1和A2 β-酪蛋白静态体外消化Zeta电位图Figure 7. Zeta potential of A1 and A2 β-casein in static in vitro digestion2.3.4 分子量的变化
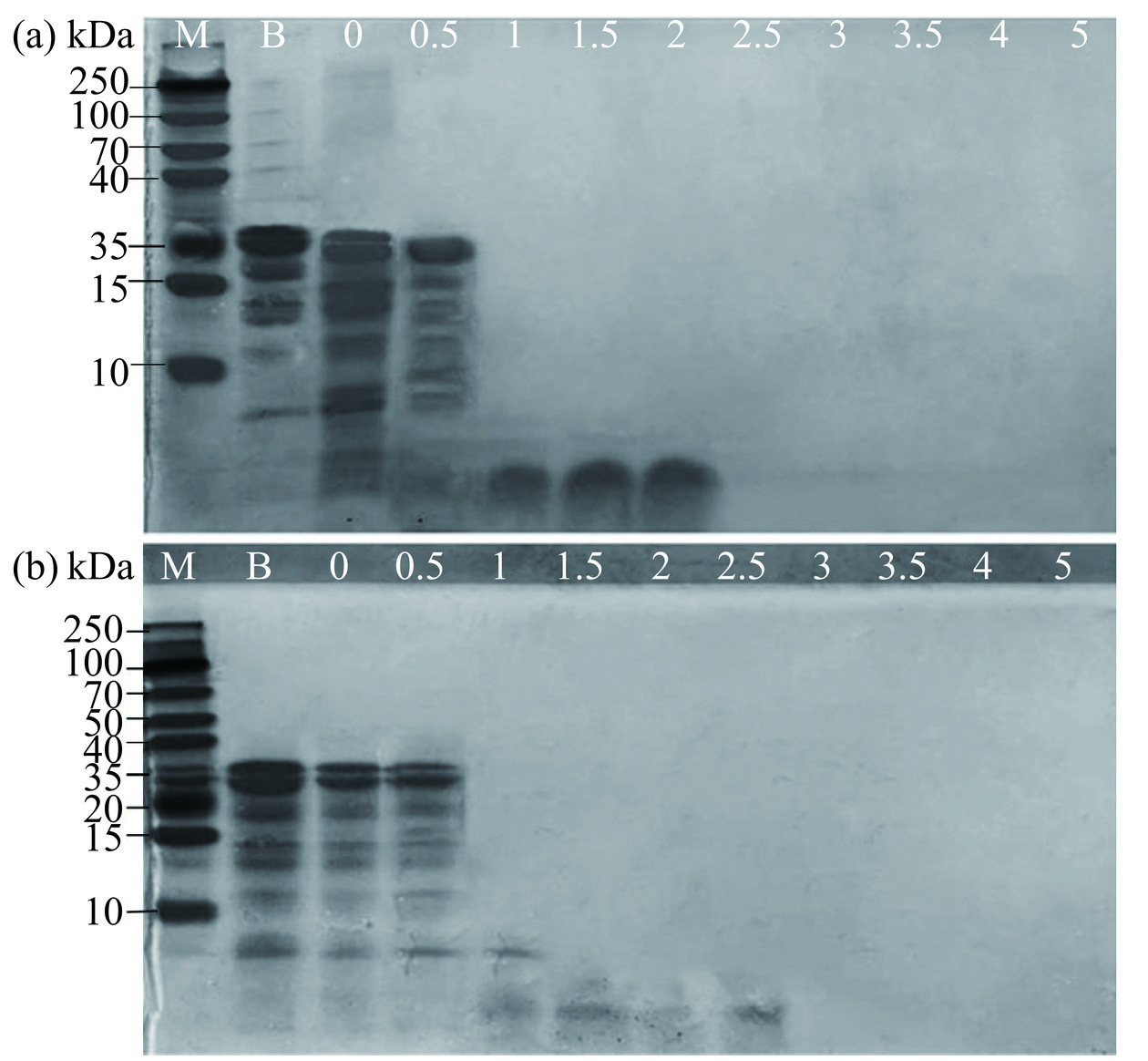
通过对SDS-PAGE图谱的结果分析,随着消化时间的延长,大分子的β-酪蛋白分子量逐渐消失,而低分子量多肽带数增加,分解为分子量<10 kDa小分子量的肽段。由图8可知,A2 β-酪蛋白在1.5 h后就无明显条带,但A1 β-酪蛋白在1.5 h后依然存在少量小分子物质,这可能是因为A2 β-酪蛋白更易暴露出活性位点与胃蛋白酶结合发生更大程度的水解,这与Daniloski等[12]得出了相反的结论,他们发现A2 β-酪蛋白通过形成更致密的蛋白网络延缓了其分解速度,这可能是因为其使用了体外半动态胃消化,与本研究体外静态消化存在差异。目前关于A2 β-酪蛋白的实验可能存在样本规模较小的问题,因此,对于A1和A2 β-酪蛋白在胃消化阶段的具体情况仍需进一步研究。
![]() 图 8 A1 (a)和A2 (b) β-酪蛋白静态体外消化SDS-PAGE 图谱注:M为标准分子量Marker;B为β-酪蛋白标准品;0、0.5、1、1.5、2、2.5、3、3.5、4、5为A1和A2 β-酪蛋白经模拟胃肠消化后不同时间段的产物。Figure 8. SDS-PAGE spectrum of A1 (a) and A2 (b) β-casein in static in vitro digestion
图 8 A1 (a)和A2 (b) β-酪蛋白静态体外消化SDS-PAGE 图谱注:M为标准分子量Marker;B为β-酪蛋白标准品;0、0.5、1、1.5、2、2.5、3、3.5、4、5为A1和A2 β-酪蛋白经模拟胃肠消化后不同时间段的产物。Figure 8. SDS-PAGE spectrum of A1 (a) and A2 (b) β-casein in static in vitro digestion3. 结论
本研究对来自同一牧场的不同奶牛的原料奶进行鉴定,分别得到A1牛奶和A2牛奶,并通过分离纯化获得A1和A2 β-酪蛋白,其浓度分别为0.62 mg/mL和0.66 mg/mL,纯度分别为95.28%和96.60%。此外,在整个消化期间,随着时间的增加,A1和A2 β-酪蛋白的消化率显著增加(P<0.05),并在5.0 h基本被完全消化,且两种β-酪蛋白的最终消化率均在90%左右,并无显著差异(P>0.05)。同时随着酶解的进行,A1和A2 β-酪蛋白水解度也显著增加(P<0.05),然而,A1 β-酪蛋白最终水解度低于A2 β-酪蛋白。消化结束后,A1 β-酪蛋白水解产物的粒径大于A2 β-酪蛋白,并且A2 β-酪蛋白消化后的电位绝对值也大于A1 β-酪蛋白,这些结果表明A2 β-酪蛋白的水解程度要更高一些,形成的粒径更小一些,更利于人体消化吸收。本项研究揭示了A1和A2 β-酪蛋白在静态体外消化过程中表现出的理化特性差异,在一定程度上表明A2 β-酪蛋白可能具有较高的生物利用度,为后续食品工业对A2 β-酪蛋白衍生产品的开发提供了一定参考依据。
-
![]()
图 1 244 bp β-酪蛋白基因型AS-PCR产物的琼脂糖凝胶电泳图谱
注:泳道1为标准分子量Marker(100~2000 bp);泳道2~11为A1特异性引物扩增结果;泳道12~21为A2特异性引物扩增结果。
Figure 1. Agarose gel electrophoresis pattern of AS-PCR products of β-casein genotypes with 244 bp
![]()
图 2 阴离子交换色谱柱分离A1(a)和A2(b)酪蛋白层析图
注:α-CN、β-CN、κ-CN分别表示α-酪蛋白、β-酪蛋白、κ-酪蛋白。
Figure 2. Separation of A1 (a) and A2 (b) casein chromatograms on an anion-exchange column
![]()
图 3 A1与A2 β-酪蛋白分离峰的洗脱液SDS-PAGE图谱
注:泳道1:标准分子量Marker;泳道2:β-酪蛋白标准品;泳道3:A1 β-酪蛋白样品;泳道4:A2 β-酪蛋白样品。
Figure 3. SDS-PAGE spectrum of eluates from A1 and A2 casein separated peaks
![]()
![]()
图 5 A1和A2 β-酪蛋白体外水解度变化
Figure 5. Changes of in vitro hydrolysis degree of A1 and A2 β-casein
![]()
图 6 A1(a)和A2(b)β-酪蛋白静态体外消化粒径分布
Figure 6. Particle size distribution of A1 (a) and A2 (b) β-casein in static in vitro digestion
![]()
图 7 A1和A2 β-酪蛋白静态体外消化Zeta电位图
Figure 7. Zeta potential of A1 and A2 β-casein in static in vitro digestion
-
[1] GIGLIOTI R, OKINO C H, AZEVEDO B T, et al. Novel LNA probe-based assay for the A1 and A2 identification of β-casein gene in milk samples[J]. Food Chemistry:Molecular Sciences,2021,3:100055.
[2] CHEN X Q, FAN R, WANG Y B, et al. Bovine milk β-casein:Structure, properties, isolation, and targeted application of isolated products[J]. Comprehensive Reviews in Food Science and Food Safety,2024,23(2):e13311. doi: 10.1111/1541-4337.13311
[3] SUN Y L, DING Y X, LIU B Q, et al. Recent advances in the bovine β-casein gene mutants on functional characteristics and nutritional health of dairy products:Status, challenges, and prospects[J]. Food Chemistry,2024,443:138510. doi: 10.1016/j.foodchem.2024.138510
[4] OSMAN A, ZUFFA S, WALTON G, et al. Post-weaning A1/A2 β-casein milk intake modulates depressive-like behavior, brain μ-opioid receptors, and the metabolome of rats[J]. Iscience,2021,24(9):103048. doi: 10.1016/j.isci.2021.103048
[5] 陈龙, 付王艳, 方琼燕, 等. 牛乳中β-酪蛋白基因分型及β-酪啡肽-7的研究进展[J]. 中国乳品工业,2019,47(8):29−34. [CHEN L, FU W Y, FANG Q Y, et al. Recent advances of β-casein genotyping in bovine milk and potential impact of β-casomorphin-7 on human health[J]. China Dairy Industry,2019,47(8):29−34.] doi: 10.3969/j.issn.1001-2230.2019.08.007 CHEN L, FU W Y, FANG Q Y, et al. Recent advances of β-casein genotyping in bovine milk and potential impact of β-casomorphin-7 on human health[J]. China Dairy Industry, 2019, 47(8): 29−34. doi: 10.3969/j.issn.1001-2230.2019.08.007
[6] DE VASCONCELOS M L, OLIVEIRA L M F S, HILL J P, et al. Difficulties in establishing the adverse effects of β-casomorphin-7 released from β-casein variants—A review[J]. Foods,2023,12(17):3151. doi: 10.3390/foods12173151
[7] CHIA J S J, MCRAE J L, KUKULJAN S, et al. A1 beta-casein milk protein and other environmental pre-disposing factors for type 1 diabetes[J]. Nutrition & Diabetes,2017,7(5):e274.
[8] LI X, LU X R, LIU M, et al. The immunomodulatory effects of A2 β-casein on immunosuppressed mice by regulating immune responses and the gut microbiota[J]. Nutrients,2024,16(4):519. doi: 10.3390/nu16040519
[9] BROOKE-TAYLOR S, DWYER K, WOODFORD K, et al. Systematic review of the gastrointestinal effects of A1 compared with A2 β-casein[J]. Advances in Nutrition,2017,8(5):739−748. doi: 10.3945/an.116.013953
[10] CATTANEO S, MASOTTI F, STUKNYTE M, et al. Impact of in vitro static digestion method on the release of β-casomorphin-7 from bovine milk and cheeses with A1 or A2 β-casein phenotypes[J]. Food Chemistry,2023,404:134617. doi: 10.1016/j.foodchem.2022.134617
[11] RAMAKRISHNAN M, ZHOU X P, DYDAK U, et al. Gastric emptying of new-world milk containing A1 and A2 β-casein is more rapid as compared to milk containing only A2 β-casein in lactose maldigesters:A randomized, cross-over trial using magnetic resonance imaging[J]. Nutrients,2023,15(4):801. doi: 10.3390/nu15040801
[12] DANILOSKI D, HAILU Y, BRODKORB A, et al. Impact of β-casein phenotype on the physical properties of skim milk powders and their subsequent digestion characteristics[J]. Food Hydrocolloids,2024,152:109918. doi: 10.1016/j.foodhyd.2024.109918
[13] MENG Y, ZHOU Y B, LI H T, et al. Effectiveness of growing-up milk containing only A2 β-casein on digestive comfort in toddlers:A randomized controlled trial in China[J]. Nutrients,2023,15(6):1313. doi: 10.3390/nu15061313
[14] 张园园. β酪啡肽-7功能评价及A2 β酪蛋白基因型奶牛鉴定[D]. 扬州:扬州大学, 2023. [ZHANG Y Y. Function evaluative of β casomorphim-7 and identification of A2 β-casein genotype cows[D]. Yangzhou:Yangzhou University, 2023.] ZHANG Y Y. Function evaluative of β casomorphim-7 and identification of A2 β-casein genotype cows[D]. Yangzhou: Yangzhou University, 2023.
[15] 刘纳. 酪蛋白组分分离、纯化研究[D]. 成都:西华大学, 2011. [LIU N. Studies on fractionation and purification of casein individuals[D]. Chengdu:Xihua University, 2011.] LIU N. Studies on fractionation and purification of casein individuals[D]. Chengdu: Xihua University, 2011.
[16] 鲁杏茹. A2型β-酪蛋白消化产物特性及免疫调节作用研究[D]. 哈尔滨:东北农业大学, 2023. [LU X R. Study on digestive product characteristics and immunomodulatory effects of A2 β-casein[D]. Harbin:Northeast Agricultural University, 2023.] LU X R. Study on digestive product characteristics and immunomodulatory effects of A2 β-casein[D]. Harbin: Northeast Agricultural University, 2023.
[17] 赵烜影. 水牛β-酪蛋白亚型结构表征及体外消化和抗氧化性能分析[D]. 哈尔滨:东北农业大学, 2019. [ZHAO H Y. Structure characterization, digestion in vitro and antioxidation of β-casein phenotypes in buffalo[D]. Harbin:Northeast Agricultural University, 2019.] ZHAO H Y. Structure characterization, digestion in vitro and antioxidation of β-casein phenotypes in buffalo[D]. Harbin: Northeast Agricultural University, 2019.
[18] EGGER L, MENARD O, BAUMANN C, et al. Digestion of milk proteins:Comparing static and dynamic in vitro digestion systems with in vivo data[J]. Food Research International,2019,118:32−39. doi: 10.1016/j.foodres.2017.12.049
[19] 何思宇. 叶绿素—酪蛋白纳米分散体系的构建及初步评价[D]. 上海:上海交通大学, 2019. [HE S Y. Preparation and preliminary evaluation of chlorophylls-casein nanodispersion[D]. Shanghai:Shanghai Jiao Tong University, 2019.] HE S Y. Preparation and preliminary evaluation of chlorophylls-casein nanodispersion[D]. Shanghai: Shanghai Jiao Tong University, 2019.
[20] 田浩. 可溶性大豆多糖在酸性条件下稳定不同乳蛋白的影响因素研究[D]. 无锡:江南大学, 2021. [TIAN H. Factors influencing the stabilization of different milk proteins by soluble soybean polysaccharides under acidic conditions[D]. Wuxi:Jiangnan University, 2021.] TIAN H. Factors influencing the stabilization of different milk proteins by soluble soybean polysaccharides under acidic conditions[D]. Wuxi: Jiangnan University, 2021.
[21] KULBABA R, SAKHATSKYI M, LIASHENKO Y. Comparative analysis of A1 and A2 allele detection efficiency for bovine CSN2 gene by AS-PCR methods[J]. Acta Biochimica Polonica,2023,70(1):205−209.
[22] RAHIMI Z, GHOLAMI M, RAHIMI Z, et al. Evaluation of beta-casein locus for detection of A1 and A2 alleles frequency using allele specific PCR in native cattle of Kermanshah, Iran[J]. Biharean Biologist,2015,9(2):85−87.
[23] 何圣发, 龙彩云, 王娇, 等. 牛乳主要过敏原αS1-酪蛋白的纯化鉴定及其多克隆抗体的制备[J]. 食品工业科技,2022,43(15):106−114. [HE S F, LONG C Y, WANG J, et al. Purification, identification and polyclonal antibody development for cow’s milk major allergen αS1-casein[J]. Science and Technology of Food Industry,2022,43(15):106−114.] HE S F, LONG C Y, WANG J, et al. Purification, identification and polyclonal antibody development for cow’s milk major allergen αS1-casein[J]. Science and Technology of Food Industry, 2022, 43(15): 106−114.
[24] THEKKILAVEEDU S, KRISHNASWAMI V, MOHANAN D P, et al. Lactic acid-mediated isolation of alpha-, beta- and kappa-casein fractions by isoelectric precipitation coupled with cold extraction from defatted cow milk[J]. International Journal of Dairy Technology,2020,73(1):31−39. doi: 10.1111/1471-0307.12646
[25] LI S S, HU Q, CHEN C, et al. Formation of bioactive peptides during simulated gastrointestinal digestion is affected by α s1-casein polymorphism in buffalo milk[J]. Food Chemistry,2020,313:126159. doi: 10.1016/j.foodchem.2020.126159
[26] KERN L, FABRE O, SCHER J, et al. Chemical fractionation of caseins by differential precipitation:Influence of pH, calcium addition, protein concentration and temperature on the depletion in α-and β-caseins[J]. International Journal of Food Science & Technology,2020,55(2):542−552.
[27] OKAWARA Y, HIRANO H, KIMURA A, et al. Phos-tag diagonal electrophoresis precisely detects the mobility change of phosphoproteins in Phos-tag SDS-PAGE[J]. Journal of Proteomics,2021,231:104005. doi: 10.1016/j.jprot.2020.104005
[28] 徐一宁, 曹传爱, 孔保华, 等. 不同形态的可得然凝胶对肌原纤维蛋白凝胶特性和体外消化特性的影响[J]. 食品工业科技,2024,45(16):73−84. [XU Y N, CAO C A, KONG B H, et al. Effects of incorporation of different forms of curdlan gels on the gelling properties and in vitro digestibility of myofibrillar protein gels[J]. Science and Technology of Food Industry,2024,45(16):73−84.] XU Y N, CAO C A, KONG B H, et al. Effects of incorporation of different forms of curdlan gels on the gelling properties and in vitro digestibility of myofibrillar protein gels[J]. Science and Technology of Food Industry, 2024, 45(16): 73−84.
[29] JIANG Y, LI S, YUAN R, et al. Preparation of casein hydrolysate by two-step enzymatic hydrolysis and its immunomodulatory activities in vitro and in vivo[J]. Food Bioscience,2024,59:104007. doi: 10.1016/j.fbio.2024.104007
[30] VOROB’EV M M, ACIKGOZ B D, GULER G, et al. Proteolysis of micellar β-casein by trypsin:Secondary structure characterization and kinetic modeling at different enzyme concentrations[J]. International Journal of Molecular Sciences,2023,24(4):3874. doi: 10.3390/ijms24043874
[31] REICHE A M, MARTIN-HERNANDEZ M C, NEFF A S, et al. The A1/A2 β-casein genotype of cows, but not their horn status, influences peptide generation during simulated digestion of milk[J]. Journal of Dairy Science,2024,107(9):6425−6436. doi: 10.3168/jds.2024-24403
[32] TU M L, LIU H X, ZHANG R Y, et al. Analysis and evaluation of the inhibitory mechanism of a novel angiotensin-I-converting enzyme inhibitory peptide derived from casein hydrolysate[J]. Journal of Agricultural and Food Chemistry,2018,66(16):4139−4144. doi: 10.1021/acs.jafc.8b00732
[33] MUDGIL P, ALMAZROUI M, REDHA A A, et al. Cow and camel milk-derived whey and casein protein hydrolysates demonstrated effective antifungal properties against selected candida species[J]. Journal of Dairy Science,2022,105(3):1878−1888. doi: 10.3168/jds.2021-20944
[34] SONG Y, WANG Z C, JI H, et al. Fatty acid modification of casein bioactive peptides nano-assemblies, synthesis, characterization and anticarcinogenic effect[J]. International Journal of Biological Macromolecules,2024,254:127718. doi: 10.1016/j.ijbiomac.2023.127718
-
其他相关附件
-
PDF格式
EI Certificate 48KB
-
 下载:
下载:







 下载:
下载:

计量
- 文章访问数: 117
- HTML全文浏览量: 35
- PDF下载量: 15



