


Efficacy and Beneficial Metabolites of Postbiotics Derived from Composite Probiotics
-
摘要: 为了研究复合益生菌后生元的功能和作用机制,本研究以副干酪乳酪杆菌ProSci-101和青春双歧杆菌B8589复合发酵的后生元为研究对象,利用代谢组学技术分析了复合益生菌后生元的有益代谢产物。结果显示,复合益生菌后生元的抑菌能力和抗氧化能力显著(P<0.05)强于单菌株发酵后生元。代谢组学分析结果显示,复合益生菌后生元与ProSci-101和B8589单菌株发酵的后生元相比,分别存在159和161个差异代谢产物,差异代谢通路主要为氨基酸和甘油磷脂代谢等。复合益生菌后生元中富集了生物碱、黄酮、有机酸等有益代谢物。研究为复合益生菌后生元的开发及应用提供了理论依据。Abstract: In order to study the functions and mechanisms of the postbiotics derived from composite probiotics, this study focused on the postbiotics produced by the co fermentation of Lacticaseibacillus paracasei ProSci-101 and Bifidobacterium adolescentis B8589. Metabolomics techniques were used to analyze the beneficial metabolites of the postbiotics. The results showed that the antibacterial and antioxidant abilities of the postbiotics derived from composite probiotics were significantly (P<0.05) stronger than those of single strain fermentation postbiotics. Metabolomics analysis showed that there were 159 and 161 differential metabolites in the postbiotics derived from composite probiotics compared to the postbiotics derived from ProSci-101 and B8589 single strains, respectively. The differential metabolic pathways were mainly amino acid and glycerophospholipid metabolism. postbiotics derived from composite probiotics are enriched with beneficial metabolites such as alkaloids, flavonoids, and organic acids. The study provides a theoretical basis for the development and application of postbiotics derived from composite probiotics.
-
Keywords:
- probiotics /
- postbiotics /
- antibacterial /
- antioxidant /
- metabolites
-
2019年,国际益生菌和益生元科学协会将灭活益生菌归为后生元,并将后生元定义为灭活菌体和/或灭活菌体的代谢产物并且对人体健康有益的微生物制剂[1]。在2023年初中国生物发酵产业协会发表益生菌制品乳酸菌类后生元的团体标准,将乳酸菌类后生元定义为:将检验合格的原辅料经配料、混合、杀菌、冷却后,接一种或几种乳酸菌类益生菌进行发酵,将发酵后的液体或固体进行灭活,并对其中菌体细胞进行破壁或不破壁处理、固液分离或不分离,添加或不添加载体,再经干燥或不干燥处理的一类后生元[2]。
后生元主要包括灭活微生物、微生物裂解物、细胞成分和代谢产物[3]。灭活微生物是指通过特定方法(热处理或非热处理)杀死菌株的细胞,制备后生元的菌株包括国家卫生行政部门发布的适合在食品中使用的菌株以及经过安全评估的益生菌菌株,灭活乳杆菌和双歧杆菌最早被人们所研究并推广应用[4]。目前市场上的后生元产品多以单一菌株为主,但是单一菌株有益代谢产物相对有限[3],功效难以最大化。已有众多研究发现复合益生菌产品效果要优于单菌株益生菌产品[5−6],复合益生菌后生元含有更丰富的碳水化合物、蛋白质、酶、脂质、维生素等[7]。因此,开发多菌株复合后生元产品将是未来发展的趋势。
因为对健康的积极影响和良好的加工贮藏特性,后生元在食品工业中得到了广泛应用,然而,目前多菌株复合益生菌后生元的功能和作用机制尚未完全明了,严重影响了后生元产业的可持续发展[8−9]。实验室前期通过筛选发现副干酪乳酪杆菌ProSci-101和青春双歧杆菌B8589具有良好的益生特性和发酵特性,其中青春双歧杆菌B8589已被证明和副干酪乳酪杆菌共同发酵制作发酵乳饮料具有更多的对人体有益的代谢产物[10]。本研究通过副干酪乳酪杆菌ProSci-101和青春双歧杆菌B8589发酵复合益生菌后生元,对复合益生菌后生元的基本的特性和有益代谢物进行研究,以期为复合益生菌后生元工业化生产提供一定的理论依据和数据支持。
1. 材料与方法
1.1 材料与仪器
副干酪乳酪杆菌ProSci-101(Lacticaseibacillus paracasei ProSci-101,ProSci-101)、青春双歧杆菌B8589(Bifidobacterium adolescentis B8589,B8589) 均由内蒙古农业大学乳酸菌种质资源库(LABCC)提供;肠炎沙门氏菌(Salmonella enterica CICC 10982)、大肠埃希氏菌(Escherichia coli CICC 23657) 购自中国工业微生物菌种保藏管理中心;金黄色葡萄球菌(Staphylococcus aureus ATCC 12600) 购自美国模式培养物集存库;单核增生李斯特氏菌(Listeria monocytogenes CMCC 53-3) 由内蒙古农业大学乳品生物技术与工程教育部重点实验室保藏;大豆粉C42 黑龙江农垦龙王食品有限公司;大豆分离蛋白粉 90% 皇氏集团股份有限公司;脱脂乳粉 广东鼎盛食品配料有限公司;乳糖酶(≥5000 NLU/g) 晟东方贸易(北京)有限公司;NB液体培养基 广州环凯微生物科技有限公司;DPPH自由基清除能力检测试剂盒、羟自由基清除能力检测试剂盒、超氧阴离子含量检测试剂盒、总抗氧化能力(T-AOC)检测试剂盒 北京索莱宝科技有限公司。
HJH-C1112超净工作台 上海智诚分析仪器制造有限公司;HA-300MII1高压蒸汽灭菌锅 日本HIRAYAMA公司;SRH 60-70均质机 上海申鹿均质机有限公司;雷磁pHS-3C型pH计 上海精密科学仪器有限公司;BSA124S-CW天平 Sartorius;GNP-9080恒温培养箱 天津市中环实验电炉有限公司;Nanodrop 1000微量紫外分光光度计 美国赛默飞世尔科技公司;CytoFLEX流式细胞仪 美国Beckman公司。
1.2 实验方法
1.2.1 复合益生菌后生元的制备
将170 g脱脂乳粉、5 g大豆粉C42和0.3 g乳糖酶加入1000 mL蒸馏水中,60 ℃下搅拌15 min,水合30 min,之后于20 Mpa压力下均质10 min处理后,放入水浴锅,升温至90 ℃,杀菌30 min,在水浴冷却至37 ℃制备用于益生菌发酵的豆乳;对杨阳[11]的方法进行改进,分别按照1×107 CFU/mL和5×106 CFU/mL的接种量接种ProSci-101和B8589,于37 ℃恒温发酵至pH4.60以下终止发酵。待发酵完成后将其预热至60 ℃,添加5%的麦芽糊精搅拌15 min,进行杀菌灭活(95 ℃ 15 min),然后进行零压均质10 min以及胶体磨研磨。最后将灭活的发酵豆乳经喷雾干燥(进风温度160 ℃,出风温度45 ℃)得到复合益生菌后生元。设定实验组为PPB(ProSci-101接种量为1×107 CFU/mL和B8589接种量为5×106 CFU/mL复合发酵),对照组为PP(ProSci-101单菌发酵,接种量为1×107 CFU/mL)和PB(B8589单菌发酵,接种量为5×106 CFU/mL)。
1.2.2 复合益生菌后生元的基本特性的测定
1.2.2.1 复合益生菌后生元酸度的测定
酸度包括pH和滴定酸度。采用pH计测量样本的pH,并根据国家标准GB 5009.239-2016采用“NaOH标准溶液滴定法”测定滴定酸度,结果以吉尔涅尔度(°T)表示。每个样品设置三个平行实验进行测定。
1.2.2.2 复合益生菌后生元制备前活菌数的测定
将样品用无菌的PBS复溶后,5000×g、4 ℃离心5 min,重悬,弃沉淀,连续稀释样品,稀释105倍。然后将复溶后的样品用浓度为10 μmol/L的cFDA分子探针进行染色,在37 ℃避光孵育20 min,取出后再加入最终浓度为5 μmol/L的PI染料对样品进行染色,振荡1 min,暗室孵育15 min,使用流式细胞仪检测荧光强度[12]。
1.2.2.3 复合益生菌后生元抑制致病菌能力的测定
以肠炎沙门氏菌、大肠埃希氏菌、金黄色葡萄球菌、单核增生李斯特菌为指示菌应用抑菌圈法测定后生元的抑菌能力。采用平板计数法来调整指示菌的菌悬液浓度至约105 CFU/mL,取200 μL的菌悬液加到20 mL的液体NB培养基中(温度控制在45 ℃以下),混合均匀后立即倒入培养皿中待其凝固,用1 mL灭菌后的枪头在培养基上打孔,初始抑菌圈为1 cm。随后按1%、5%、50%的浓度,用无菌的生理盐水重新溶解后生元,分别向琼脂孔中加入200 μL,设置两组重复实验。将培养基在4 ℃条件下培养4 h以促扩散,之后转移到37 ℃的培养箱中继续培养12 h,最终测量并记录抑菌圈的直径[13]。
1.2.2.4 复合益生菌后生元抗氧化性的测定
将复合益生菌后生元按1%、5%、50%的比例用灭菌后的生理盐水复溶后测定其抗氧化能力。测定的指标分别为DPPH自由基的清除能力、羟自由基的清除能力、超氧阴离子的含量、总抗氧化能力(T-AOC),根据索莱宝检测试剂盒的说明书分别测定。
1.2.3 复合益生菌后生元代谢物质的测定
将制备好的后生元样品委托迈维代谢公司检测代谢产物。使用ProteoWizard软件将原始数据转换为mzXML格式,然后使用R语言(使用XCMS内核)进行处理,如峰值识别、提取、配准和整合。随后,将数据与自建的BiotreeDB二级质谱数据库(2.1版)进行比较,以进行物质注释,算法评分阈值设定为0.3。
1.3 数据处理
在本研究中,所有数据都经过至少三次平行实验的验证。使用Microsoft Excel对实验数据进行汇编和结构化处理。使用SPSS 21.0软件进行独立样本T检验分析,显著性水平设定为0.05。使用Origin Lab 2021软件生成了数据图表。对于代谢数据的分析,采用了迈维云工具和MetaboAnalyst软件进行主成分分析(PCA)和正交偏最小二乘判别分析(OPLS-DA)。差异代谢物筛选基于P值小于0.05和VIP值大于1的标准,随后对这些差异代谢物使用迈维云工具进行了代谢通路富集分析。
2. 结果与分析
2.1 复合益生菌后生元的理化特性
2.1.1 复合益生菌后生元的pH、滴定酸度
经喷雾干燥后制备的三组后生元样品如图1所示,三组样品在外观上无明显差异。
![]() 图 1 PPB组、PP组、PB组的样品注:A为PPB样品;B为PP样品;C为PB样品。Figure 1. Samples from PPB, PP and PB groups
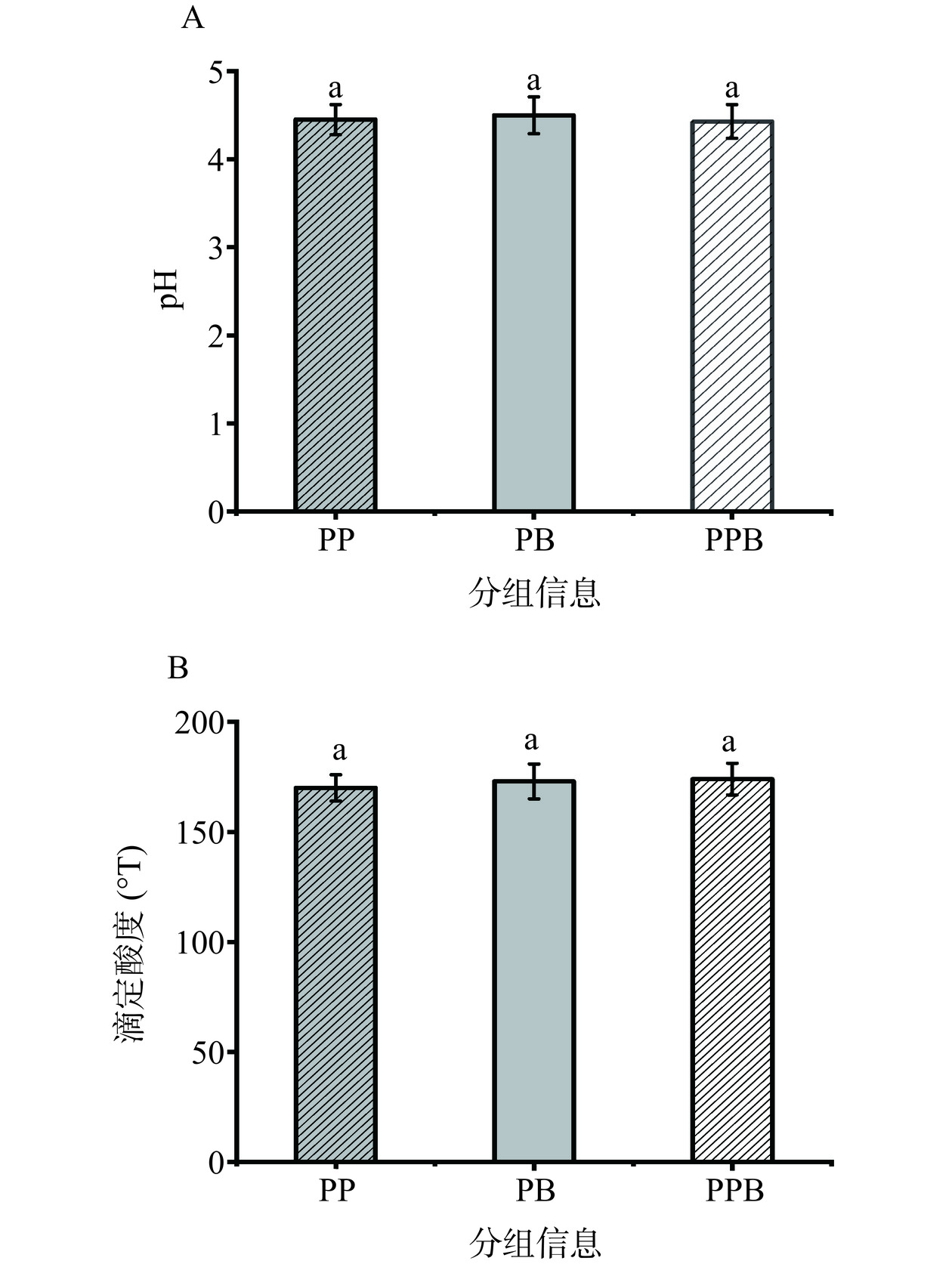
图 1 PPB组、PP组、PB组的样品注:A为PPB样品;B为PP样品;C为PB样品。Figure 1. Samples from PPB, PP and PB groupsPP、PB、PPB组的pH分别为4.45、4.50、4.43,三组之间的pH无显著差异(P>0.05)(图2A)。PP、PB、PPB组的滴定酸度分别为170 °T、173 °T、174 °T,三组之间的滴定酸度也无显著差异(P>0.05)(图2B),均在人体可摄入的范围之内。
![]() 图 2 样品组和对照组pH及滴定酸度的测定结果注:柱状图上不同小写字母代表不同组间差异显著(P<0.05),图3同。Figure 2. Results of pH values and titration acidity in the sample and control groups
图 2 样品组和对照组pH及滴定酸度的测定结果注:柱状图上不同小写字母代表不同组间差异显著(P<0.05),图3同。Figure 2. Results of pH values and titration acidity in the sample and control groups2.1.2 制备后生元的发酵乳的菌落总数
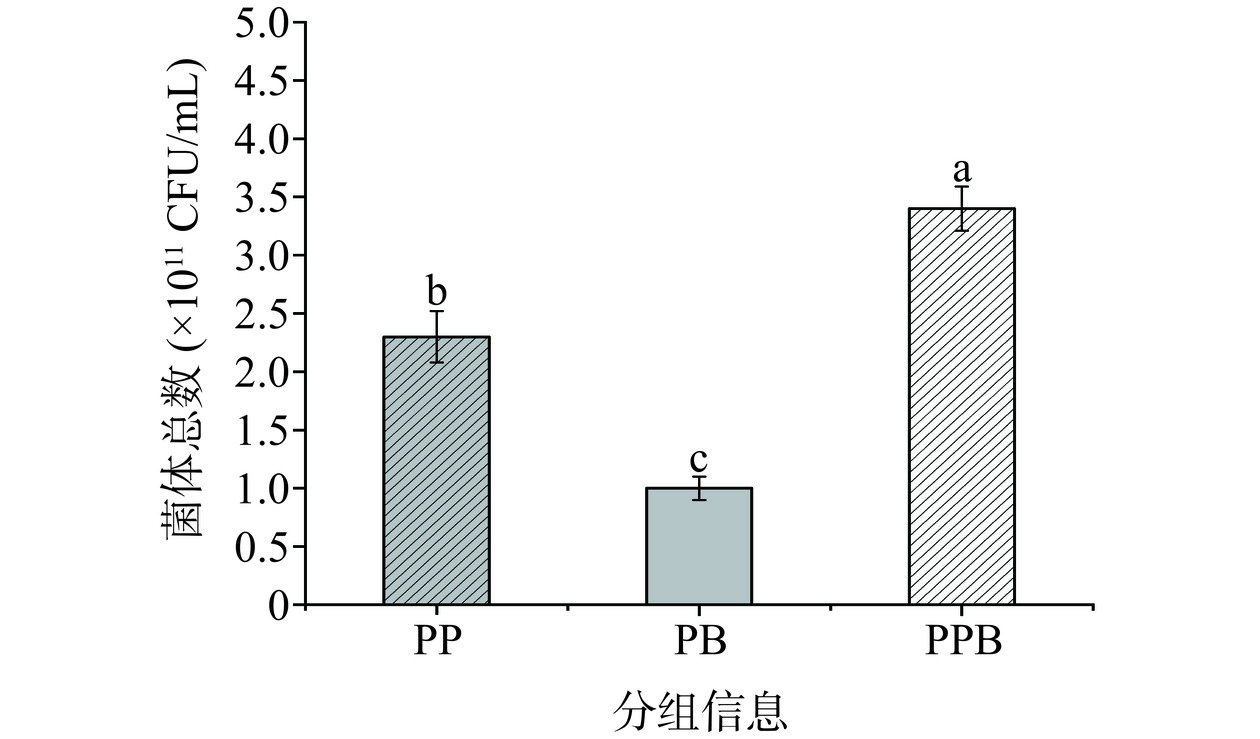
PP组的菌落总数为2.3×1011 CFU/mL,PB组的菌落总数为1×1011 CFU/mL,PPB组的菌落总数为3.4×1011 CFU/mL,PPB组显著高于PP组和PB组(P<0.05)(图3)。复合益生菌后生元(PPB组)的菌落总数与两株菌单独发酵的菌落总数之和基本一致,表明菌株在复合发酵时生长并未受到抑制。
2.1.3 复合益生菌后生元的抑菌活性
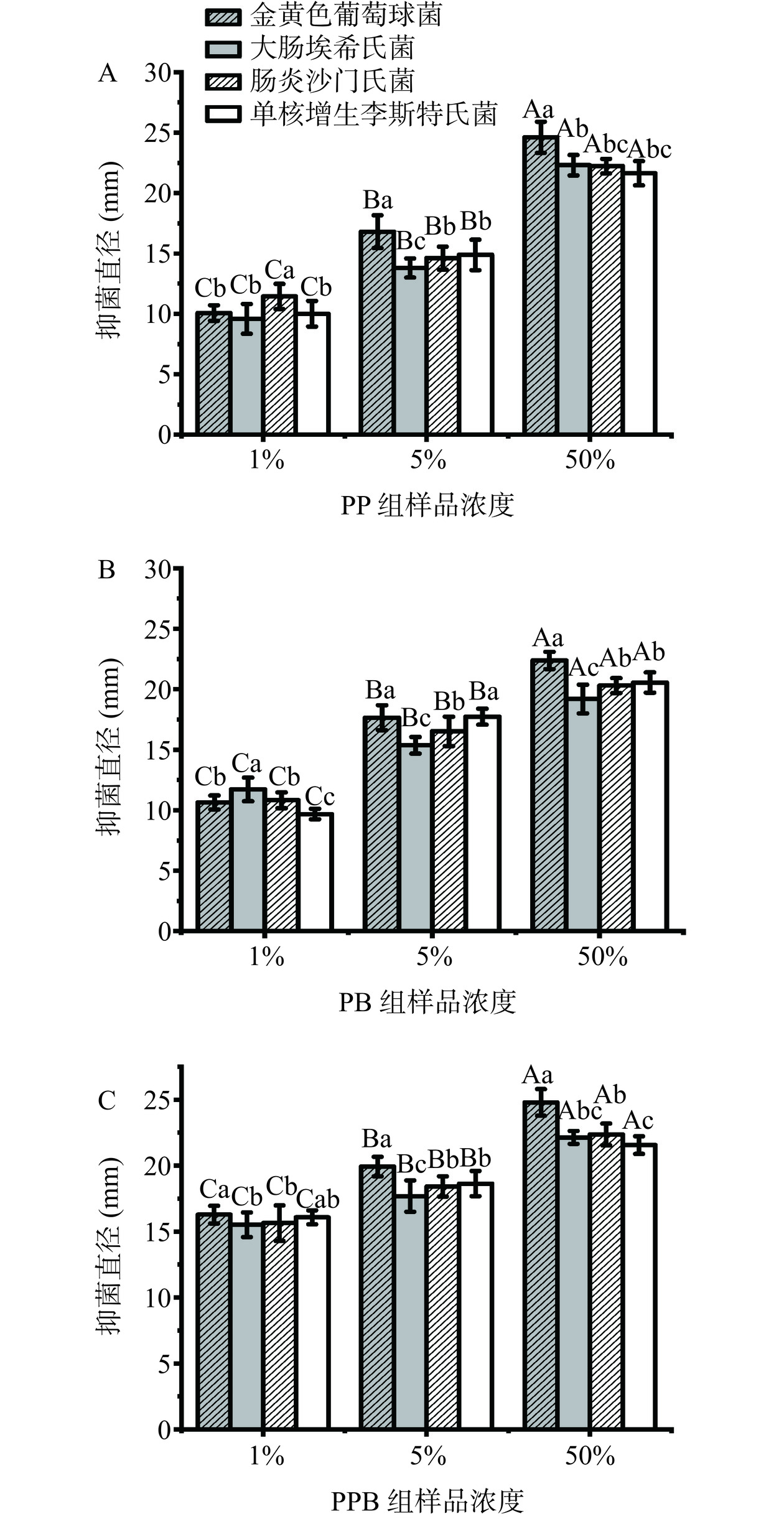
图4显示了PP组、PB组和PPB组对几种细菌(金黄色葡萄球菌、大肠埃希氏菌、肠炎沙门氏菌、单核增生李斯特菌)的抑菌活性。随着三组样品浓度的升高,抑菌活性也有显著的增加(P<0.05)。在样品浓度为1%和5%时,PPB组对金黄色葡萄球菌、大肠埃希氏菌、肠炎沙门氏菌、单核增生李斯特氏菌的抑菌直径显著大于PP组和PB组(P<0.05)。在样品浓度为50%时,PPB组的抑菌直径也略大于PP组和PB组。由此可知,PPB组对金黄色葡萄球菌、大肠埃希氏菌、肠炎沙门氏菌、单核增生李斯特菌有更好的抑菌效果。同时,PPB组对于金黄色葡萄球菌的抑菌能力也强于多种分离自羊奶中乳酸菌所制备的后生元[14],对于大肠杆菌的抑菌能力也强于戊糖片球菌所制备的后生元[15]。
![]() 图 4 不同浓度的三组样品抑菌活性的测定结果注:图中X轴1%、5%、50%分别为复合益生菌后生元复溶后的浓度;柱状图上不同大写字母代表不同浓度组间差异显著(P<0.05);不同小写字母代表同一浓度组内差异显著(P<0.05)。Figure 4. Results of the antibacterial activity of three groups of samples at different concentrations
图 4 不同浓度的三组样品抑菌活性的测定结果注:图中X轴1%、5%、50%分别为复合益生菌后生元复溶后的浓度;柱状图上不同大写字母代表不同浓度组间差异显著(P<0.05);不同小写字母代表同一浓度组内差异显著(P<0.05)。Figure 4. Results of the antibacterial activity of three groups of samples at different concentrations2.1.4 复合益生菌后生元的抗氧化能力
2.1.4.1 DPPH自由基的清除能力
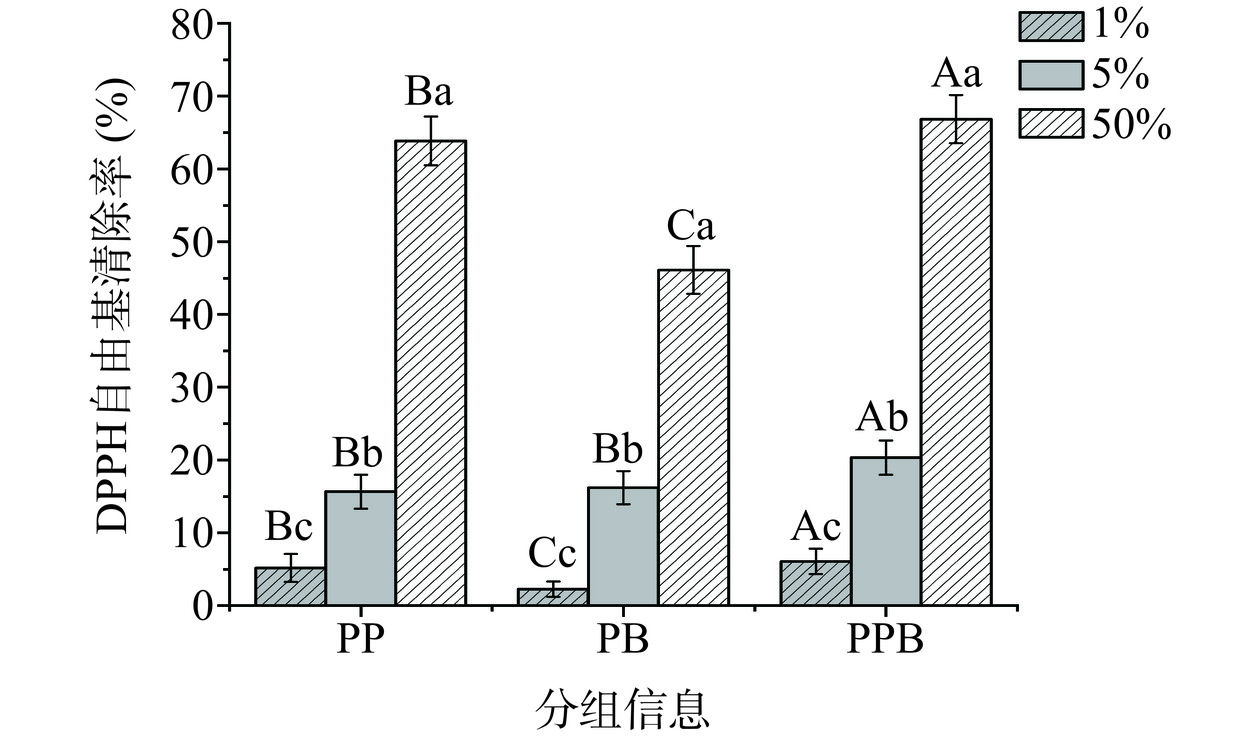
DPPH自由基是一种稳定的氮中心的自由基,是评估样本抗氧化活性的关键指标[16]。由图5可知,当样品浓度由1%升高到50%时,三组样品的DPPH自由基清除能力随着样品浓度升高而增强。而PPB组样品的DPPH自由基清除率在3个浓度下均显著高于其他两组(P<0.05)。此外,1%浓度的PP组DPPH自由基清除率高于肠膜明串珠菌胞外多糖制备后生元的DPPH自由基清除率,且PPB组样品的DPPH自由基清除能力更优异[17]。综上所述,PPB组相较于其他两组有更强的DPPH自由基清除能力。
![]()
2.1.4.2 羟自由基的清除能力
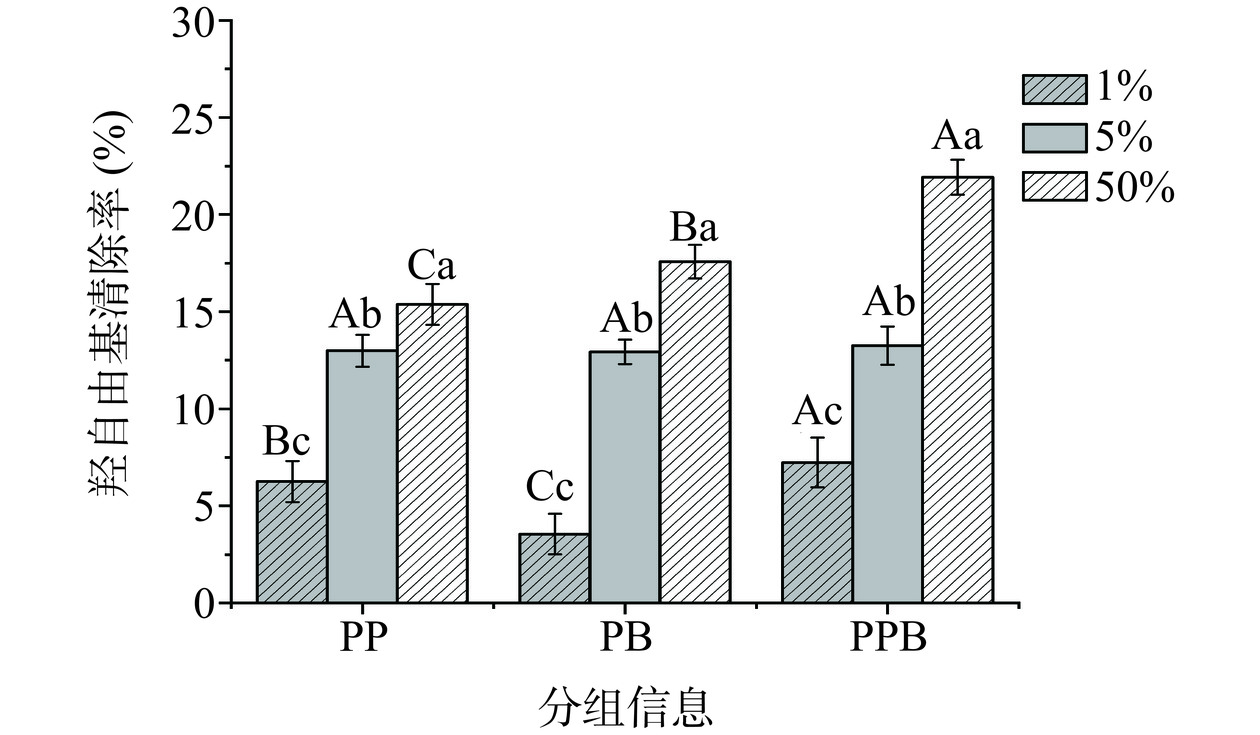
羟自由基(·OH)是人体内一种活性很强的自由基,通常被认为是已知化学结构中最活跃的活性氧自由基之一。在人体内,羟基自由基能够与生物体内的各种成分迅速反应,对机体产生潜在的损害[18]。由图6可知,当PPB组的样品浓度为1%时,其羟基自由基清除率为7.23%,高于PP组的6.26%、PB组的3.55%(P<0.05)。所有样品清除羟基自由基的能力都随着样品浓度的增加而增强,其中PPB组的羟基自由基清除率最终达到了21.92%,高于相同浓度下的PP组及PB组(P<0.05)。因此,可以得出结论,PPB组具有较强的羟基自由基清除能力,这对于其在抗氧化和保健方面的应用具有重要意义。
![]() 图 6 不同浓度的三组样品羟自由基的清除能力Figure 6. Hydroxyl radical scavenging capacity of the three groups of samples at different concentrations
图 6 不同浓度的三组样品羟自由基的清除能力Figure 6. Hydroxyl radical scavenging capacity of the three groups of samples at different concentrations2.1.4.3 超氧阴离子的含量
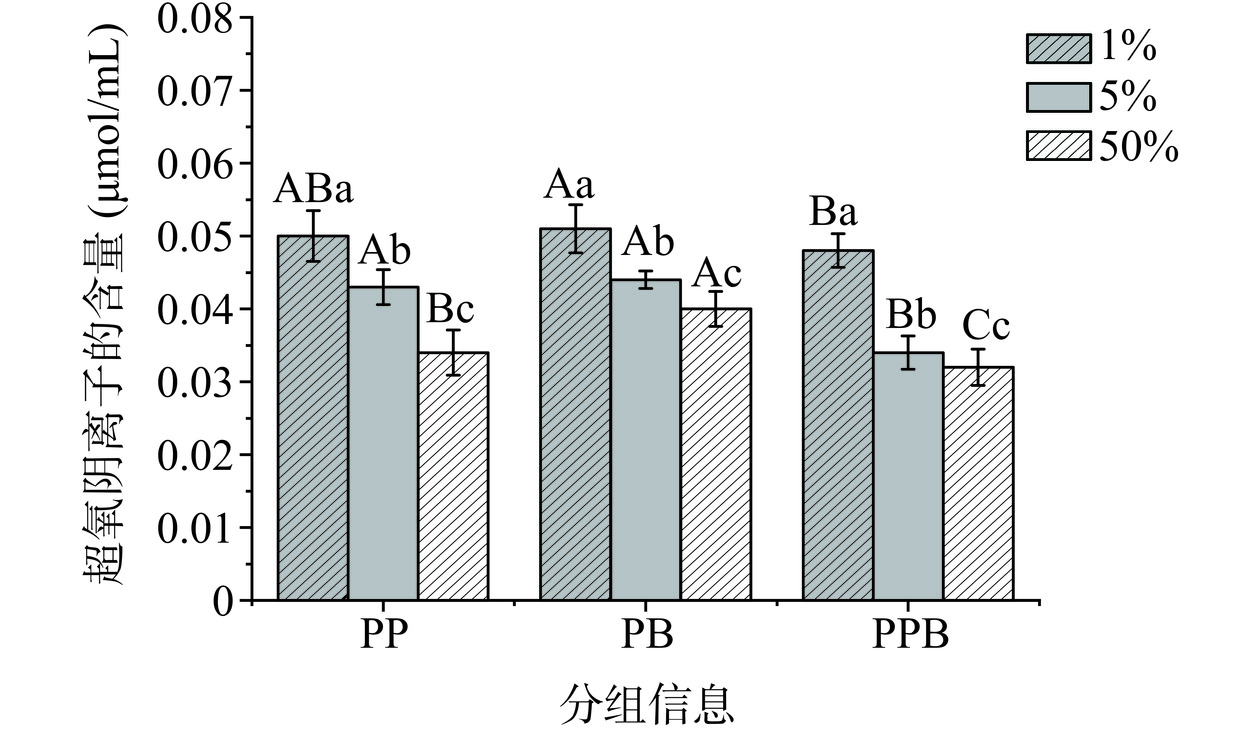
超氧阴离子在生物体内参与免疫反应和信号传递,但其过量积累可能对细胞膜和生物大分子造成损害[19]。图7中,超氧阴离子含量与样品浓度呈现反比,当样品浓度为1%时,PP组、PB组、PPB组的超氧阴离子含量分别为0.050、0.051、0.048 μmol/mL,PPB组的超氧阴离子含量显著低于PB组(P<0.05)。随着样品浓度的增加超氧阴离子的含量会逐渐下降,当浓度为50%时PPB组(0.032 μmol/mL)超氧阴离子含量显著低于PP组(0.034 μmol/mL)和PB组(0.040 μmol/mL)(P<0.05)。结果表明,PPB组的超氧阴离子含量较低,对细胞膜及生物大分子产生破坏作用最小。
![]() 图 7 不同浓度的三组样品超氧阴离子的含量Figure 7. Content of superoxide anion in three sets of samples at different concentrations
图 7 不同浓度的三组样品超氧阴离子的含量Figure 7. Content of superoxide anion in three sets of samples at different concentrations2.1.4.4 总抗氧化能力
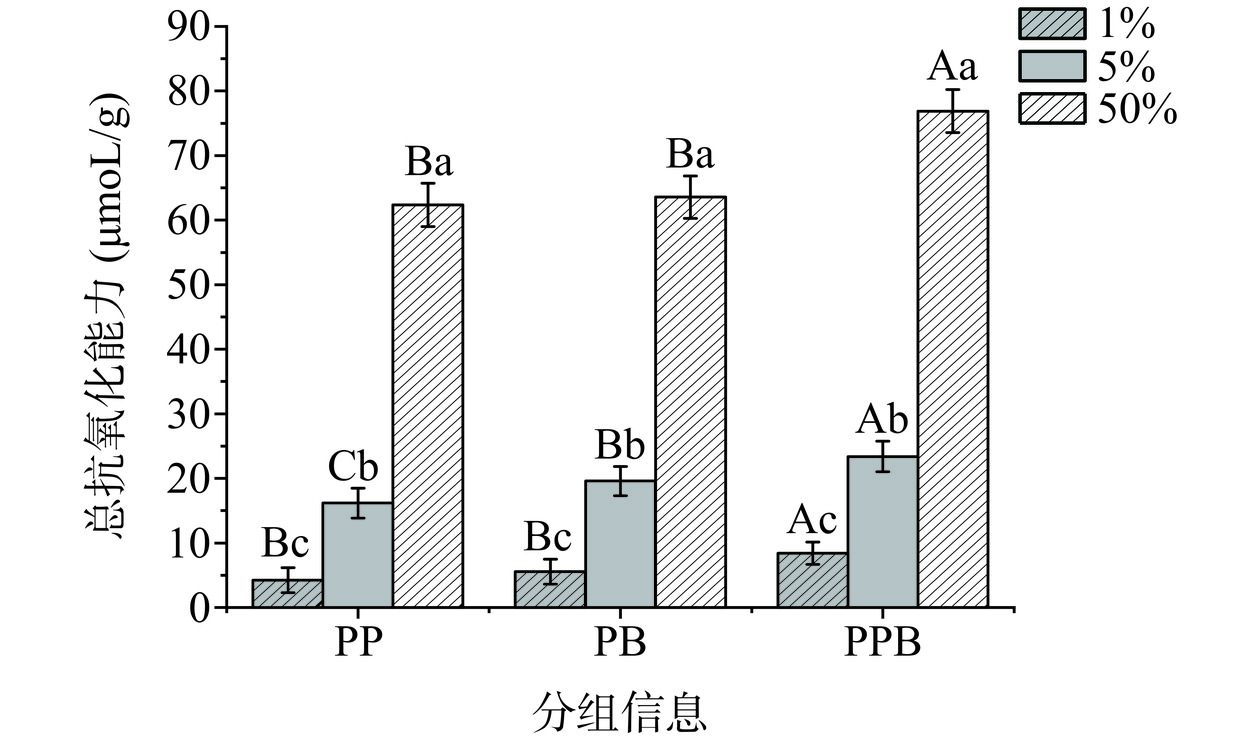
由图8可知,随着样品浓度升高,其总抗氧化性能明显增强。虽然在样品浓度为1%时整体的抗氧化能力较低,但PPB组的抗氧化能力为8.39 μmol/mL,高于PP组的4.23 μmol/mL、PB组的5.55 μmol/mL。当样品浓度增加为50%时,PP组的抗氧化能力为62.36 μmol/mL,PB组的抗氧化能力为63.56 μmol/mL,PPB组的抗氧化能力为76.89 μmol/mL,PPB组显著高于其他两组(P<0.05),因此PPB组相较于其他组有较强的总抗氧化能力。此外,由于实验采用豆乳发酵,发酵豆乳成分也可能会对后生元的抗氧化能力产生影响,但本文重点讨论复合益生菌后生元与单菌株发酵后生元的比较。因此,豆乳作为三者都有的控制变量可以消除由其贡献的抗氧化能力对实验结果的影响。
![]() 图 8 不同浓度的三组样品总抗氧化能力Figure 8. Total antioxidant capacity of the three groups of samples at different concentrations
图 8 不同浓度的三组样品总抗氧化能力Figure 8. Total antioxidant capacity of the three groups of samples at different concentrations2.2 复合益生菌后生元代谢物检测
2.2.1 代谢物主成分分析
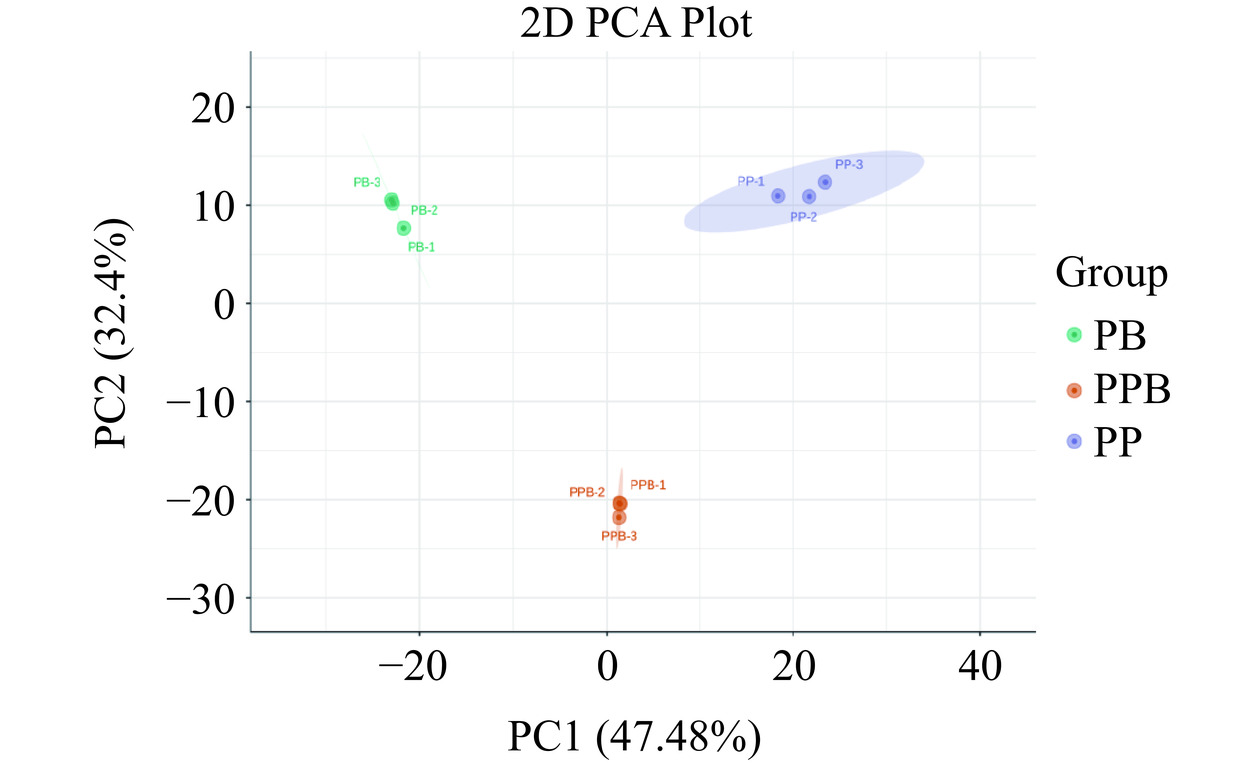
通过观察PP组、PB组和PPB组的得分图(图9),模型的总变异性为79.88%,说明实验模型质量较好。PP组、PB组和PPB组的组内相对聚集,有较好的平行性。而组间差异明显,说明ProSci-101、B8589的复合发酵(PPB组)和分别单独发酵(PP组和PB组)对复合益生菌后生元的代谢产物产生明显影响。
2.2.2 代谢物正交偏最小二乘判别法分析
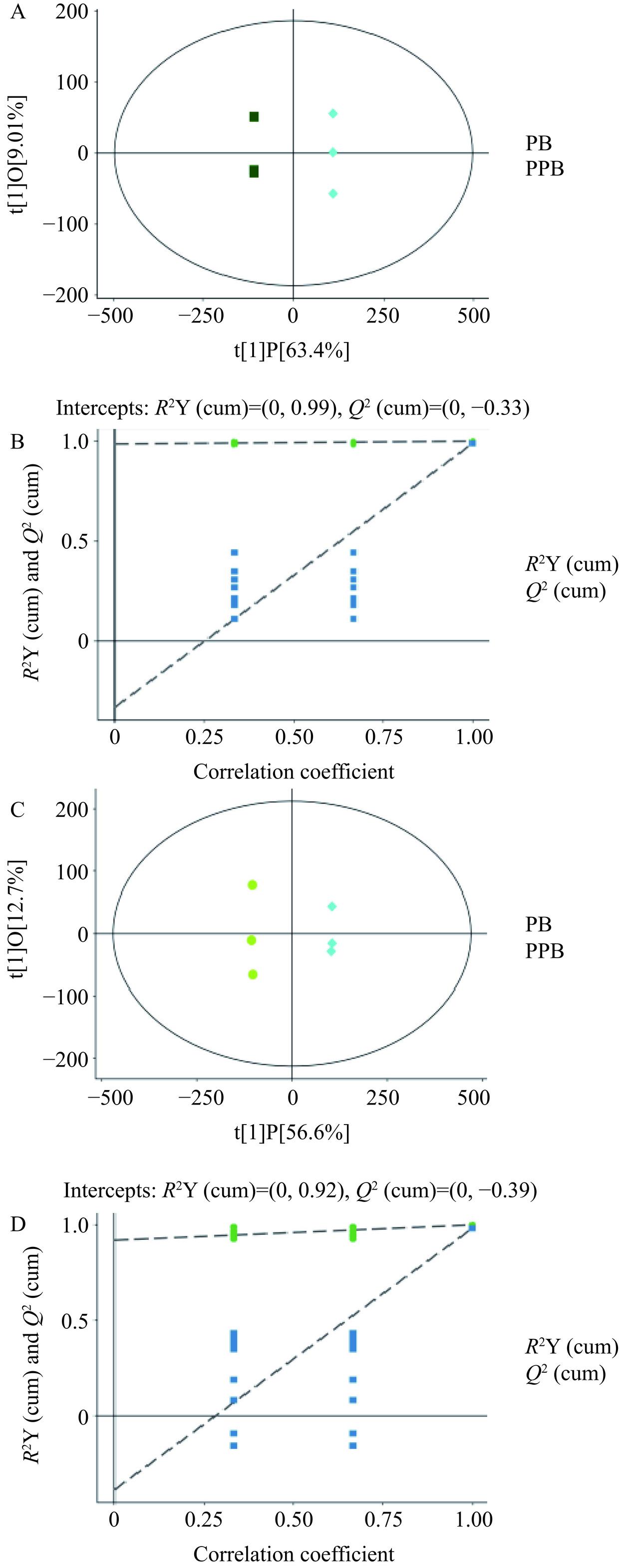
通过正交偏最小二乘法判别分析显示(图10),PB组与PPB组、PP组与PPB组组间差异较大且组内重复性好,表明不同样本代谢物之间存在显著差异。同时在置换检验图中,R2和Q2值大于任意一个随机排列后得到的R2和Q2值,且模型Q2的斜率均大于R2的斜率,说明模型不存在过拟合现象,实验数据真实可靠,其结果可信。
![]() 图 10 PB、PPB、PP三组样品的OPLS-DA图和置换检验图注:A、B分别代表复合益生菌后生元PB组与PPB组的OPLS-DA模型及置换检验图;C和D分别代表复合益生菌后生元PP组与PPB组的OPLS-DA模型及置换检验图。Figure 10. OPLS-DA plots and displacement test plots of the three samples of PB, PPB and PP
图 10 PB、PPB、PP三组样品的OPLS-DA图和置换检验图注:A、B分别代表复合益生菌后生元PB组与PPB组的OPLS-DA模型及置换检验图;C和D分别代表复合益生菌后生元PP组与PPB组的OPLS-DA模型及置换检验图。Figure 10. OPLS-DA plots and displacement test plots of the three samples of PB, PPB and PP2.2.3 复合益生菌后生元差异代谢物分析
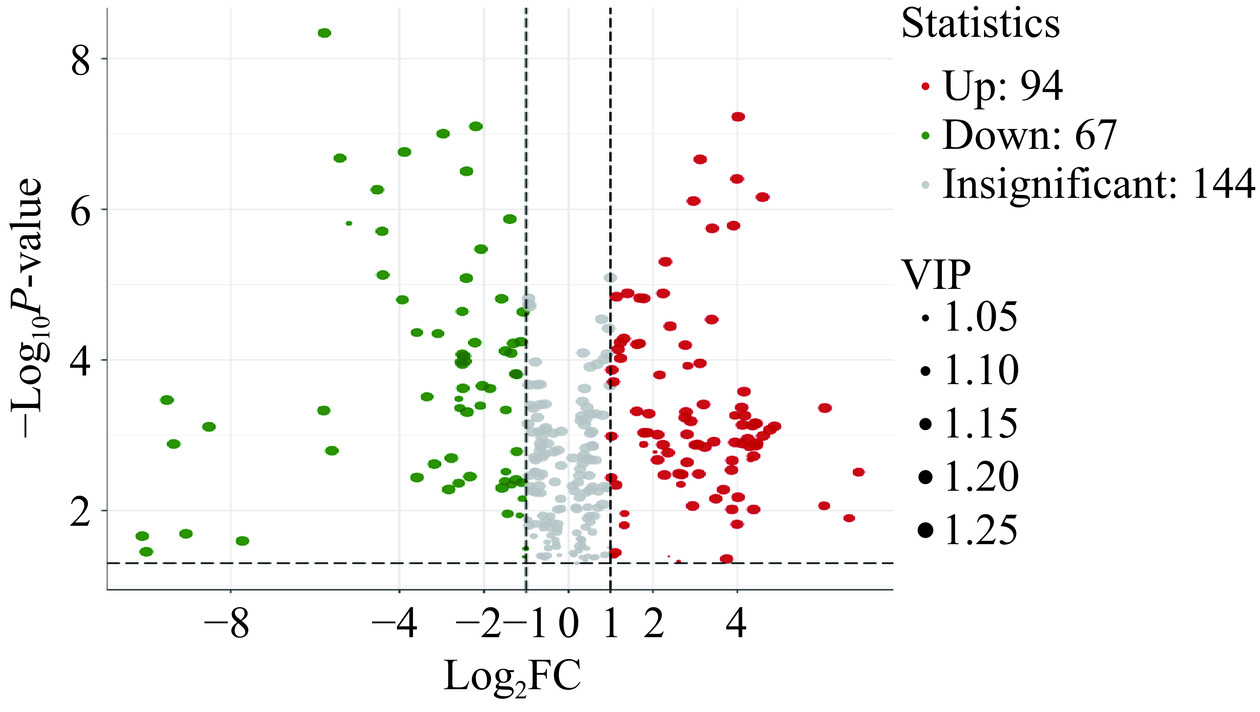
在PB和PPB组中共检测到305种代谢物质,以VIP>1、P<0.05、FC≥2或FC≤0.5作为筛选标准,对样品进行检验得到,PB组和PPB组差异代谢物质共有161种(附表1,详见本刊官网http://www.spgykj.com/文章的网络版)。相比于PB组,PPB组有94种代谢物质呈显著上调,67个代谢物质呈显著下调(图11)。
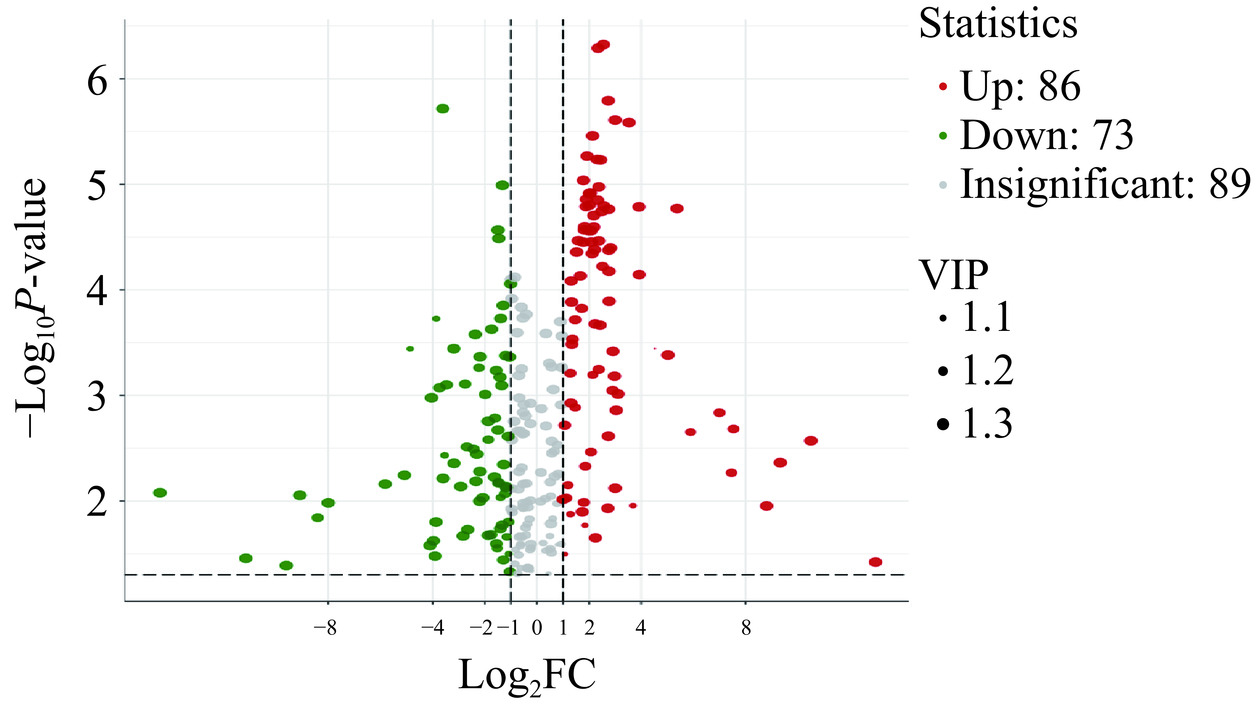
PP组和PPB组共检测出248种代谢物质,以VIP>1、P<0.05和FC≥2或FC≤0.5作为筛选标准,PP组和PPB组差异代谢物共有159种(附表2,详见本刊官网http://www.spgykj.com/文章的网络版);相比于PP组,在PPB组中有86种差异代谢物显著(P<0.05)上调,73个差异代谢物显著下调(图12)。
2.2.4 复合益生菌后生元代谢通路富集分析
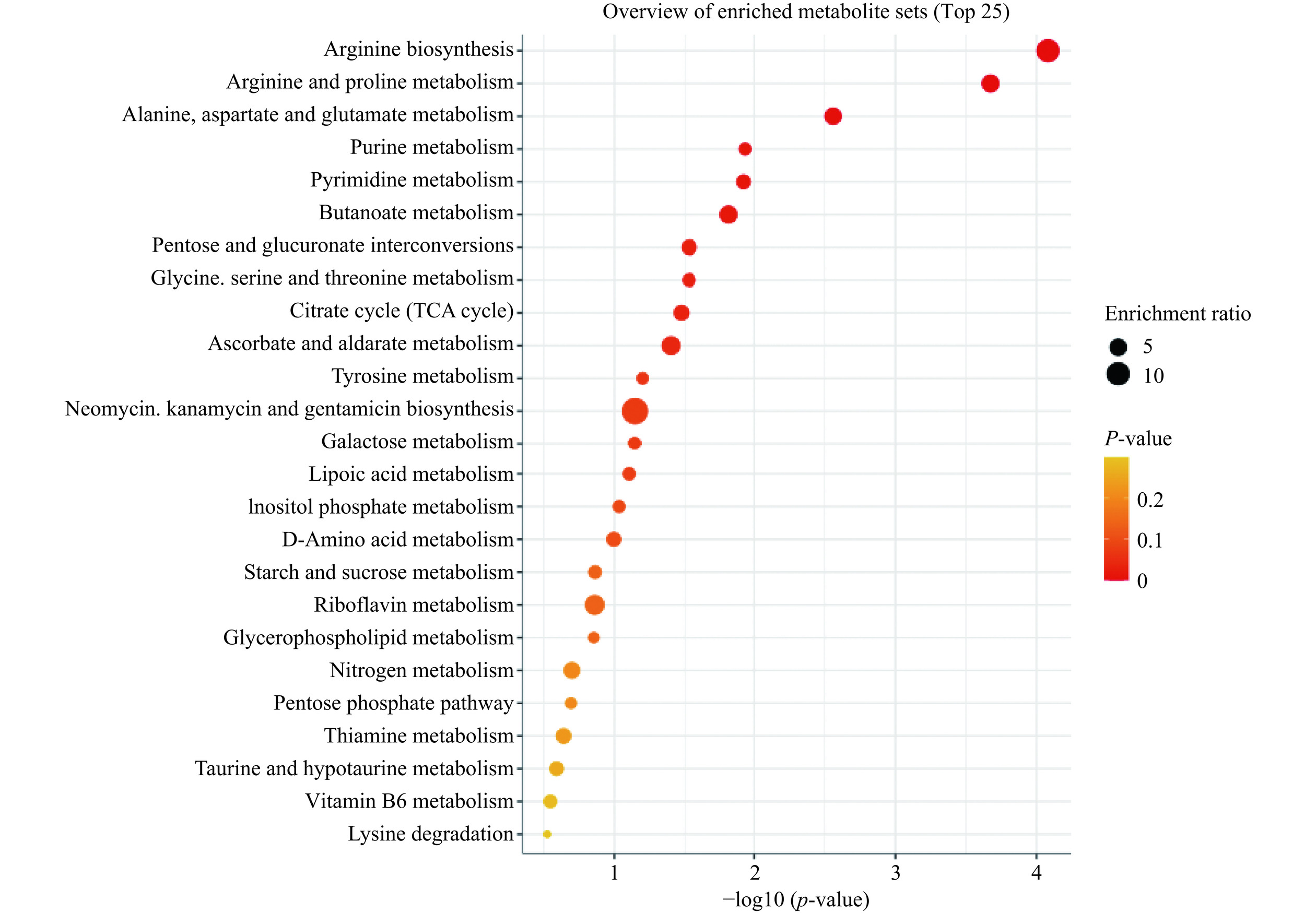
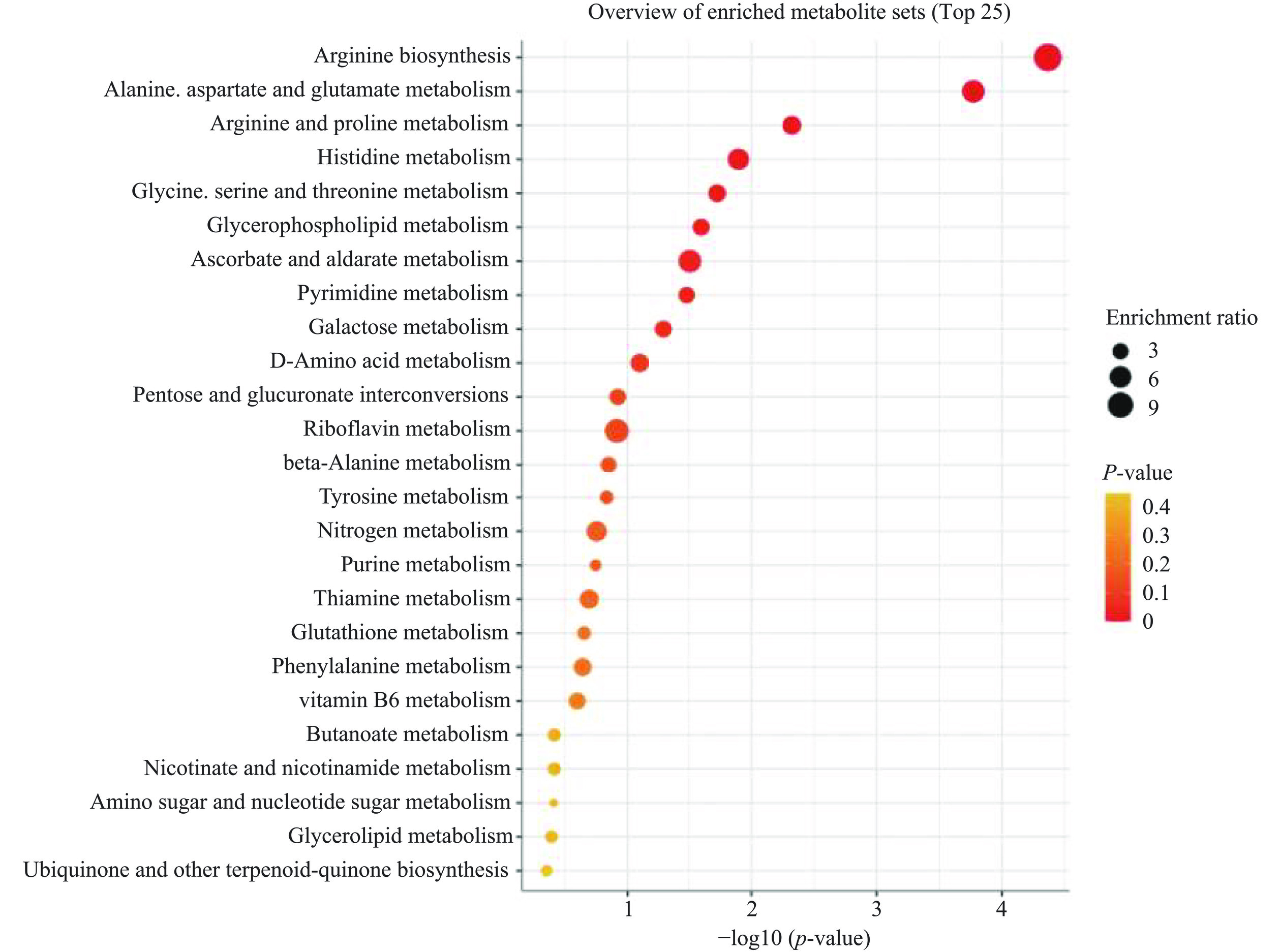
通过KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库对差异代谢物质进行通路富集分析,分析了PB组和PPB组中161种差异显著的代谢产物,确定了25条主要代谢通路(图13),其中显著的富集通路有精氨酸生物合成、精氨酸和脯氨酸代谢、丙氨酸,天冬氨酸和谷氨酸代谢、嘌呤代谢、嘧啶代谢、丁酸代谢、戊糖和葡萄糖醛酸的相互转化、柠檬酸循环(TCA循环)等。
![]() 图 13 PB组和PPB组样品差异代谢物的代谢通路富集图Figure 13. Metabolic pathway enrichment plots of differential metabolites between PB and PPB group samples
图 13 PB组和PPB组样品差异代谢物的代谢通路富集图Figure 13. Metabolic pathway enrichment plots of differential metabolites between PB and PPB group samplesPP组与PPB组中差异显著的代谢产物进行代谢通路富集分析,其显著或较显著的通路有精氨酸生物合成、丙氨酸,天冬氨酸和谷氨酸代谢、精氨酸和脯氨酸代谢、组氨酸代谢、甘氨酸,丝氨酸和苏氨酸代谢、甘油磷脂代谢、半乳糖代谢、氨基酸代谢、酪氨酸代谢(图14)。
![]() 图 14 PP组和PPB组样品差异代谢物的代谢通路富集图Figure 14. Metabolic pathway enrichment plots of differential metabolites between PP and PPB group samples
图 14 PP组和PPB组样品差异代谢物的代谢通路富集图Figure 14. Metabolic pathway enrichment plots of differential metabolites between PP and PPB group samples2.2.5 复合益生菌后生元有益代谢物分析
代谢物差异分析表明复合益生菌后生元显著提高了氨基酸的代谢水平,特别是精氨酸和脯氨酸的代谢。其中,L-精氨酸是精氨酸生物合成代谢通路中主要的差异物质,L-精氨酸是一种阳离子半必需氨基酸,是人体及动物体内含量最丰富的含氮化合物[20]。L-精氨酸在人体营养和代谢活动中发挥着多种重要作用[21]。此外,L-精氨酸还具有较强的自由基清除能力,能够提高动物机体的抗氧能力,这为前期复合益生菌后生元的抗氧化能力优于单菌株后生元提供分子水平证据支持[22]。脯氨酸是蛋白质合成的重要部分,也是DNA合成的重要前体物质,参与细胞与分裂和增殖等生理功能[23]。这些氨基酸在复合益生菌后生元中富集,更多地参与精氨酸与脯氨酸代谢,从而使复合益生菌后生元的营养价值和生理功能得到提升。
甘油磷脂是复合益生菌后生元中另外一种富集的代谢物。磷脂代谢是脂质代谢中的关键路径,甘油磷脂是人体内最常见的磷脂类型,含量丰富,对细胞的活跃状态、新陈代谢以及基本代谢活动至关重要[24]。此外,甘油磷脂还有助于调节脂肪代谢,改善血液循环,并有助于预防心血管疾病[25−27]。在PPB组中甘油磷脂相关代谢物如PC(18:1(11Z)/18:0)、PI(16:1(9Z)/18:1(11Z))、PE(18:0/14:1(9Z))、PS(16:0/18:1(11Z))等水平均高于PB组和PP组,提示PPB组可以调控甘油磷脂代谢[28]。此外,PS已被证明可以缓解情绪、修复大脑损伤和提高记忆力等[29],还具有一定的抗炎和抗氧化能力,能够调节肠道菌群紊乱等[30]。
同时研究发现PPB组中包括生物碱、黄酮、有机酸和酚类化合物等代谢物显著增加。黄酮通常以糖苷形式存在,黄酮类化合物在多个行业中有着广泛的利用价值,如抗肿瘤、抗氧化、糖尿病防治、抗炎、抗病毒和抗感染等,黄酮类化合物因其出色的自由基清除和抗氧化能力,能够改善脂质代谢,还有助于降低心血管疾病的发生率和死亡率,在本研究中也观察到富含黄酮类化合物的复合益生菌后生元在体外表现出较强的抗氧化能力和优异的抑菌效果[31];此外,天然的黄酮类化合物具有血管舒张作用,能有效改善心脏血管平滑肌的收缩和舒张功能[32]。有机酸类物质能降低消化系统的pH,促进胃蛋白酶原转化为胃蛋白酶,并抑制有害细菌的繁殖,为胃肠道提供能量,帮助食物在胃中的排空,以及增加内源酶的分泌,还有助于矿物质的螯合[33]。
代谢组学分析结果表明,经ProSci-101和B8589复合发酵可赋予复合益生菌后生元更为丰富的益生特性,且也发现可提高其抗氧化活性和抑菌能力的代谢众多代谢产物,如L-精氨酸和黄酮类化合物。在抗氧化实验和抑菌实验中,与PP组和PB组相比,PPB组均表现出优异的效果。结合代谢组学与表型实验结果,复合益生菌后生元通过丰富代谢产物和代谢途径从而提高抗氧化和抑菌能力来发挥潜在益生特性。
3. 结论
本研究以副干酪乳酪杆菌ProSci-101和青春双歧杆菌B8589复合发酵的后生元为研究对象,通过与单菌株发酵的后生元作比较,发现复合益生菌后生元具有更强的抑菌能力和抗氧化活性。同时,非靶向代谢组学分析显示,复合益生菌后生元中的差异代谢通路主要为氨基酸和甘油磷脂代谢等,富集了生物碱、黄酮、有机酸、酚类等有益代谢物。研究为以副干酪乳酪杆菌ProSci-101和青春双歧杆菌B8589复合发酵复合益生菌后生元的开发和应用奠定了基础。
-
![]()
图 1 PPB组、PP组、PB组的样品
注:A为PPB样品;B为PP样品;C为PB样品。
Figure 1. Samples from PPB, PP and PB groups
![]()
图 2 样品组和对照组pH及滴定酸度的测定结果
注:柱状图上不同小写字母代表不同组间差异显著(P<0.05),图3同。
Figure 2. Results of pH values and titration acidity in the sample and control groups
![]()
图 4 不同浓度的三组样品抑菌活性的测定结果
注:图中X轴1%、5%、50%分别为复合益生菌后生元复溶后的浓度;柱状图上不同大写字母代表不同浓度组间差异显著(P<0.05);不同小写字母代表同一浓度组内差异显著(P<0.05)。
Figure 4. Results of the antibacterial activity of three groups of samples at different concentrations
![]()
![]()
图 6 不同浓度的三组样品羟自由基的清除能力
Figure 6. Hydroxyl radical scavenging capacity of the three groups of samples at different concentrations
![]()
图 7 不同浓度的三组样品超氧阴离子的含量
Figure 7. Content of superoxide anion in three sets of samples at different concentrations
![]()
图 8 不同浓度的三组样品总抗氧化能力
Figure 8. Total antioxidant capacity of the three groups of samples at different concentrations
![]()
图 10 PB、PPB、PP三组样品的OPLS-DA图和置换检验图
注:A、B分别代表复合益生菌后生元PB组与PPB组的OPLS-DA模型及置换检验图;C和D分别代表复合益生菌后生元PP组与PPB组的OPLS-DA模型及置换检验图。
Figure 10. OPLS-DA plots and displacement test plots of the three samples of PB, PPB and PP
![]()
图 13 PB组和PPB组样品差异代谢物的代谢通路富集图
Figure 13. Metabolic pathway enrichment plots of differential metabolites between PB and PPB group samples
-
[1] SALMINEN S, COLLADO M C, ENDO A, et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics[J]. Nature Reviews Gastroenterology & Hepatology,2021,18(9):649−667.
[2] 胡成文, 张国芳, 王伟军, 等. 后生元的制备方法及其应用[J]. 乳品与人类,2023(3):31−38. [HU Chengwen, ZHANG Guofang, WANG Weijun, et al. Preparation method of epigenetic element and its application[J]. Dairy and Humankind,2023(3):31−38.] doi: 10.3969/j.issn.1671-5071.2023.03.005 HU Chengwen, ZHANG Guofang, WANG Weijun, et al. Preparation method of epigenetic element and its application[J]. Dairy and Humankind, 2023(3): 31−38. doi: 10.3969/j.issn.1671-5071.2023.03.005
[3] ZHONG Y J, WANG T, LUO R L, et al. Recent advances and potentiality of postbiotics in the food industry:Composition, inactivation methods, current applications in metabolic syndrome, and future trends[J]. Critical Reviews in Food Science and Nutrition, 2022:1−25.
[4] 胡文锋, 郭增雄, 方祥, 等. 热灭活嗜酸乳杆菌及其多肽类抗菌物质特性研究[J]. 饲料博览,1999(10):7−8. [HU Wenfeng, GUO Zengxiong, FANG Xiang, et al. Characterization of heat inactivatedLactobacillus acidophilus and its polypeptide antibacterial substances[J]. Feed Review,1999(10):7−8.] HU Wenfeng, GUO Zengxiong, FANG Xiang, et al. Characterization of heat inactivated Lactobacillus acidophilus and its polypeptide antibacterial substances[J]. Feed Review, 1999(10): 7−8.
[5] FAN X K, DU L, XU J, et al. Effect of single probiotics Lacticaseibacillus casei CGMCC1. 5956 and Levilactobacillus brevis CGMCC1. 5954 and their combination on the quality of yogurt as fermented milk[J]. LWT, 2022, 163:113530.
[6] LUO J Y, JIANG L, LIANG S, et al. Fermentation of Lonicera caerulea by complex probiotics and its effect on hypolipidemic[J]. Food Bioscience,2024,57:103502. doi: 10.1016/j.fbio.2023.103502
[7] HOMAYOUNI R A, AGHEBATI M L, SAMADI K H, et al. Postbiotics:A novel strategy in food allergy treatment[J]. Critical Reviews in Food Science and Nutrition,2021,61(3):492−499. doi: 10.1080/10408398.2020.1738333
[8] HUANG Y Y, ZHAO S, YAO K, et al. Physicochemical, microbiological, rheological, and sensory properties of yoghurts with new polysaccharide extracts from Lactarius volemus Fr. using three probiotics[J]. International Journal of Dairy Technology,2020,73(1):168−181. doi: 10.1111/1471-0307.12653
[9] MORADI M, MARDANI K, TAJIK H. Characterization and application of postbiotics of Lactobacillus spp. on Listeria monocytogenes in vitro and in food models[J]. LWT,2019,111:457−464. doi: 10.1016/j.lwt.2019.05.072
[10] SUN Y R, GUO S, WU T, et al. Bifidobacterium adolescentis B8589-and Lacticaseibacillus paracasei PC-01-co-fermented milk has more γ-aminobutyric acid and short-chain fatty acids than Lacticaseibacillus paracasei PC-01-fermented milk[J]. LWT,2023,179:114645. doi: 10.1016/j.lwt.2023.114645
[11] 杨阳. 副干酪乳酪杆菌PC-01和青春双歧杆菌B8589在活性乳酸菌饮料中的应用研究[D]. 呼和浩特:内蒙古农业大学, 2022. [YANG Y. Application of Lacticaseibacillus paracasei PC-01 and Bifidobacterium adolescentis B8589 in active lactobacillus beverage[D]. Hohhot:Inner MongoliaAgricultural University, 2022.] YANG Y. Application of Lacticaseibacillus paracasei PC-01 and Bifidobacterium adolescentis B8589 in active lactobacillus beverage[D]. Hohhot: Inner MongoliaAgricultural University, 2022.
[12] GANDHI A, SHAH N P. Effect of salt on cell viability and membrane integrity of Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium longum as observed by flow cytometry[J]. Food Microbiology,2015,49:197−202. doi: 10.1016/j.fm.2015.02.003
[13] 瞿茜楠, 彭传涛, 黄天, 等. 副干酪乳酪杆菌PC-724蛋白类抑菌活性代谢产物的研究[J/OL]. 中国食品学报, 1−12[2024-05-10]. http://kns.cnki.net/kcms/detail/11.4528.TS.20230417.1904.004.html. [QU Xinan, PENG Chuantao, HUANG Tian, et al. Study on antibacterial active metabolites of Lacticaseibacillus paracasei PC-724 protein[J/OL]. Journal of Chinese Institute of Food Science and, 1−12[2024-05-10]. http://kns.cnki.net/kcms/detail/11.4528.TS.20230417.1904.004.html.] QU Xinan, PENG Chuantao, HUANG Tian, et al. Study on antibacterial active metabolites of Lacticaseibacillus paracasei PC-724 protein[J/OL]. Journal of Chinese Institute of Food Science and, 1−12[2024-05-10]. http://kns.cnki.net/kcms/detail/11.4528.TS.20230417.1904.004.html.
[14] SINGH N, SHARMA C, GULHANE R D, et al. Inhibitory effects of lactobacilli of goat's milk origin against growth and biofilm formation by pathogens:An in vitro study[J]. Food Bioscience,2018,22:129−138. doi: 10.1016/j.fbio.2018.02.001
[15] BAJPAI V K, HAN J-H, RATHER I A, et al. Characterization and antibacterial potential of lactic acid bacterium Pediococcus pentosaceus 4I1 isolated from freshwater fish Zacco koreanus[J]. Frontiers in Microbiology,2016,7:2037.
[16] GULCIN İ, ALWASEL S H. DPPH radical scavenging assay[J]. Processes,2023,11(8):2248. doi: 10.3390/pr11082248
[17] PĂCULARU-BURADA B, BAHRIM G E. Extraction and antioxidant activity assessment of postbiotic exopolysaccharides produced by selected lactic acid bacteria[J]. Innovative Romanian Food Biotechnology, 2021(20).
[18] 荣梦杰. 棉酚的亚临界提取及抗氧化活性研究[D]. 北京:中国农业科学院, 2020. [RONG M J. Study on subcritical extraction and antioxidant activity of gossypol[D]. Beijing:Chinese Academy of Agricultural Sciences, 2020.] RONG M J. Study on subcritical extraction and antioxidant activity of gossypol[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020.
[19] JIE Z S, LIU J, SHU M C, et al. Detection strategies for superoxide anion:A review[J]. Talanta,2022,236:122892. doi: 10.1016/j.talanta.2021.122892
[20] WU G, MEININGER C J, KNABE D A, et al. Arginine nutrition in development, health and disease[J]. Current Opinion in Clinical Nutrition & Metabolic Care,2000,3(1):59−66.
[21] WU G, MORRIS JR S M. Arginine metabolism:nitric oxide and beyond[J]. Biochemical Journal,1998,336(1):1−17. doi: 10.1042/bj3360001
[22] ALIMOHAMMADI S, ZENDEHDEL M, BABAPOUR V. Modulation of opioid-induced feeding behavior by endogenous nitric oxide in neonatal layer-type chicks[J]. Veterinary Research Communications,2015,39:105−113. doi: 10.1007/s11259-015-9631-8
[23] 刘玉霞, 窦世娟, 王秀伶. 细菌中脯氨酸的生物合成、降解及功能[J]. 微生物学报,2021,61(11):3351−3362. [LIU Yuxia, DOU Shijuan, WANG Xiuling. Research progress in proline biosynthesis, degradation and function in bacteria[J]. Acta Microbiologica Sinica,2021,61(11):3351−3362.] LIU Yuxia, DOU Shijuan, WANG Xiuling. Research progress in proline biosynthesis, degradation and function in bacteria[J]. Acta Microbiologica Sinica, 2021, 61(11): 3351−3362.
[24] CAO P, WU Y, LI Y P, et al. The important role of glycerophospholipid metabolism in the protective effects of polyphenol-enriched Tartary buckwheat extract against alcoholic liver disease [J]. Food & Function, 2022,13(20): 10415-10425.
[25] 赵琳儒, 何盼, 李杰, 等. 柴芩宁神颗粒改善失眠大鼠睡眠作用的海马代谢组学研究[J]. 中国中药杂志,2022,47(7):1921−1931. [ZHAO Linru, HE Pan, LI Jie, et al. Sleep-improving mechanism of Chaiqin Ningshen Granules in insomnia rats:Based on hippocampal metabonomics[J]. China Journal of Chinese Materia Medica,2022,47(7):1921−1931.] ZHAO Linru, HE Pan, LI Jie, et al. Sleep-improving mechanism of Chaiqin Ningshen Granules in insomnia rats: Based on hippocampal metabonomics[J]. China Journal of Chinese Materia Medica, 2022, 47(7): 1921−1931.
[26] HUI D Y. Intestinal phospholipid and lysophospholipid metabolism in cardiometabolic disease[J]. Current opinion in lipidology,2016,27(5):507−512. doi: 10.1097/MOL.0000000000000334
[27] CHEN H, WANG Z X, QIN M, et al. Comprehensive metabolomics identified the prominent role of glycerophospholipid metabolism in coronary artery disease progression[J]. Frontiers in Molecular Biosciences, 2021,8:632950.
[28] 佟颖, 黄慧丽, 项敬, 等. 基于LC-MS研究痹宁汤对痛风性肾病小鼠肾脏代谢产物的影响[J]. 中医药学报,2023,51(11):47−54. [TONG Ying, HUANG Huili, XIANG Jing, et al. Study on the effect of bining decoction on metabolites of kidney tissues in mice with gouty nephropathy based on LC-MS[J]. Acta Chinese Medicine and Pharmacology,2023,51(11):47−54.] TONG Ying, HUANG Huili, XIANG Jing, et al. Study on the effect of bining decoction on metabolites of kidney tissues in mice with gouty nephropathy based on LC-MS[J]. Acta Chinese Medicine and Pharmacology, 2023, 51(11): 47−54.
[29] DINKLA S, PEPPELMAN M, VAN DER RAADT J, et al. Phosphatidylserine exposure on stored red blood cells as a parameter for donor-dependent variation in product quality[J]. Blood Transfusion,2014,12(2):204.
[30] ZHOU Y F, TIAN S S, QIAN L, et al. DHA-enriched phosphatidylserine ameliorates non-alcoholic fatty liver disease and intestinal dysbacteriosis in mice induced by a high-fat diet[J]. Food & Function,2021,12(9):4021−4033.
[31] 马洁桃, 张岭, 王茵. 黄酮类化合物的降脂活性及其作用机制的研究进展[J]. 中国预防医学杂志,2011,12(4):370−372. [MA Jietao, ZHANG Ling, WANG Yin. Research progress on lipid lowering activity and mechanism of flavonoid compounds[J]. Chinese Preventive Medicine,2011,12(4):370−372.] MA Jietao, ZHANG Ling, WANG Yin. Research progress on lipid lowering activity and mechanism of flavonoid compounds[J]. Chinese Preventive Medicine, 2011, 12(4): 370−372.
[32] 严啸东, 郭美丽. 黄酮类化合物抗心脑血管缺血性疾病作用的研究进展[J]. 药学实践杂志,2022,40(2):97−102. [YAN Xiaodong, GUO Meili. Research progress of the effect of flavonoids on cardiovascular and cerebrovascular ischemic diseases[J]. Journal of Pharmaceutical Practice and Service,2022,40(2):97−102.] YAN Xiaodong, GUO Meili. Research progress of the effect of flavonoids on cardiovascular and cerebrovascular ischemic diseases[J]. Journal of Pharmaceutical Practice and Service, 2022, 40(2): 97−102.
[33] 旷义文. 复合有机酸替代抗生素或氧化锌对断奶仔猪生长性能的影响及机理研究[D]. 雅安:四川农业大学, 2018. [KUANG Y W. Effects and underlying mechanisms of compoundorganic acids utilized as alternatives to antibioticsor zinc oxide in weaned piglet diets[D]. Yaan:Sichuan Agricultural University, 2018.] KUANG Y W. Effects and underlying mechanisms of compoundorganic acids utilized as alternatives to antibioticsor zinc oxide in weaned piglet diets[D]. Yaan: Sichuan Agricultural University, 2018.
-
期刊类型引用(3)
1. 高畅. 基于电子鼻分析发酵条件对酸樱桃风味的影响. 食品安全导刊. 2024(20): 95-98 .  百度学术
百度学术
2. 张轶,丁同仁,王光强. 益生菌冷冻干燥保护剂的研究进展. 工业微生物. 2024(06): 91-96 . 百度学术
3. 邓露芳,辛娜,信丽凤,黄玉海,宗文丽,王东,郭亮. 复合乳酸菌制剂对蛋鸡产蛋后期产蛋性能和蛋品质的影响. 饲料博览. 2024(04): 31-35 . 百度学术
其他类型引用(0)
-
其他相关附件
-
DOCX格式
(附表1、附表2)2024040295-复合益生菌后生元的功效及有益代谢产物的研究 51KB -
PDF格式
EI收录证明 42KB -
DOCX格式
(附表1、附表2)2024040295-复合益生菌后生元的功效及有益代谢产物的研究 51KB
-
 下载:
下载:

 下载:
下载:













计量
- 文章访问数: 176
- HTML全文浏览量: 107
- PDF下载量: 35
- 被引次数: 3



