


Progress on the Classification, Characteristics and Food Industry Application of Microbial Proteases and Lipases
-
摘要: 蛋白酶和脂肪酶作为两大工业水解酶,广泛应用于食品行业各个领域(如奶酪制作、酱油发酵、酒类酿造、肉质嫩化等)。蛋白酶和脂肪酶主要来源于动物、植物和微生物,微生物源蛋白酶和脂肪酶因获取便利、成本低廉、分子改造简便等优势而成为研究热点。目前国内外针对微生物源蛋白酶和脂肪酶的研究较多,包括不同来源产酶菌株筛选、产酶能力优化、理化特性研究、酶分子改造以期更好应用于食品加工等。本文总结了微生物源蛋白酶和脂肪酶相关研究,阐明了微生物源蛋白酶(丝氨酸蛋白酶、半胱氨酸蛋白酶、天冬氨酸蛋白酶和金属蛋白酶)和脂肪酶的分类、特性及在肉类、烘焙食品、乳制品、豆制品、酒类、茶类等的应用,为充分了解各种微生物源蛋白酶和脂肪酶的适用性、提高其使用效率、推动其更好应用于食品工业生产提供理论参考。Abstract: Proteases and lipases are two major kinds of industrial hydrolysis enzymes and widely used in various food industry (e.g., cheese making, soy sauce fermentation, liquor brewing, meat tenderization, etc.). Proteases and lipases are mainly obtained from animals, plants and microorganisms. The microbial proteases and lipases have attracted great interest due to easy obtaining, low cost and simple molecular modification. Currently, there is a significant amount of research on microbial proteases and lipases, including strain selection from different sources, optimization of enzyme production, physicochemical characteristics and molecular modification of enzyme for better application in food processing. This article summarizes the relevant research on microbial proteases and lipases. This elucidates the classification and characteristics of microbial proteases (serine protease, cysteine protease, aspartic protease and metalloprotease) and lipases, as well as their application in the food industry (e.g., meat, baking, milk, bean, liquor, tea, etc.). This paper provides a theoretical reference to fully understand the applicability of various microbial proteases and lipases, improve their utilization efficiency and promote their application in food industry.
-
Keywords:
- microbial proteases /
- microbial lipases /
- classification /
- enzymatic characteristics /
- application
-
酶是一种高效、可持续且广泛使用的绿色催化剂。与化学催化剂相比,它具有高特异性、高催化活性、在极端条件下的工作能力以及大规模生产的能力。蛋白酶和脂肪酶作为两大工业酶制剂,广泛应用于食品、皮革、洗涤和生物医药等多个行业[1]。在食品工业中,蛋白酶和脂肪酶常作为食品加工与检测的加工助剂,用于牛奶凝固、奶酪发酵、酱油和啤酒酿造、蛋白和多肽物质水解等,有助于改善食品品质并维持形态结构稳定[2]。凝乳酶是奶酪制作过程中的关键酶,它影响奶酪产量、质构和风味等,在奶酪成熟过程中发挥重要作用[3]。在焙烤食品中添加蛋白酶与脂肪酶,通过改善小麦面筋的溶解性、乳化性和发泡性,使得制作的面包更加柔软并延长产品货架期[4]。在啤酒发酵过程中添加蛋白酶能有效改善麦汁粘度和浊度,添加脂肪酶有利于合成芳香酯等风味物质,使口感更加丰富[5]。
酶制剂来源广泛,可以从植物茎叶、果实、动物内脏和微生物中获取,随着全球酶需求量的增大,动植物酶已无法满足其生产需求,微生物酶因具有生物技术应用所需特性而优于动植物酶,它不会产生有毒有害物质且易于进行基因操作,是一种安全高效的生物催化剂,全球酶总销售额约40%来自微生物[6]。针对部分酶的特性缺陷,可以通过酶分子体外定向进化技术,模拟自然进化进程[7],利用易错PCR等技术对编码水解酶基因进行随机突变,再由DNA重组、交错延伸过程、随机引导等进行基因突变,加快酶某种特征的进化速度,创造出具有新特征或性能有所改善的酶。通过蛋白质结构修饰(如引入共价或非共价键相互作用、改变底物亲和能力、调节活性位点等[8])提高酶活性、表达量、热稳定性、pH耐受性、底物特异性等。因此,微生物源酶制剂应用广泛。
目前已有多种蛋白酶、脂肪酶在微生物中成功表达。优势菌株发掘、产酶能力优化、理化特性探究和基因改造技术是研究的主要方向。本文对食品工业中常见的微生物源蛋白酶和脂肪酶进行分类,并阐述其部分理化特性,列举在食品工业中应用的部分实例,并对现存问题探究及发展趋势进行展望,为提升微生物源水解酶的适用性和商业价值提供一定参考。
1. 微生物源蛋白酶
蛋白酶主要作用于相邻氨基酸残基间的肽键,将其裂解成更短的肽或小分子氨基酸,是重要的商业化胞外酶之一,微生物源蛋白酶主要来源于细菌和真菌,细菌主要来源于枯草芽孢杆菌和大肠杆菌[9],真菌源蛋白酶可以从青霉菌、镰刀菌、曲霉菌和酵母菌中分离得到。它们广泛应用于肉类、乳制品、酒类、豆制品和功能性食品等[10]。
1.1 蛋白酶分类
蛋白酶有多种分类方式,按其水解多肽的方式可以分为外肽酶和内肽酶;根据其最适pH可将其分为酸性蛋白酶、中性蛋白酶和碱性蛋白酶。而学术上是根据催化结构域活性位点存在的官能团不同分为丝氨酸蛋白酶、半胱氨酸蛋白酶、天冬氨酸蛋白酶和金属蛋白酶[11]。本文就学术上的分类方式,对不同种类蛋白酶的来源菌株、部分酶学特性及其在食品工业的应用进行分类讨论,然而,关于微生物源半胱氨酸蛋白酶的相关研究并不多,本文就此部分简略说明。
1.1.1 丝氨酸蛋白酶及酶学特性
丝氨酸蛋白酶(Serine protease,Sps,EC 3.4.21)在工业中的需求量很大,因此蛋白酶生产候选物的分离和表征是一个重要的研究领域。其中芽孢杆菌属是丝氨酸蛋白酶的主要来源之一。Yilmaz等[12]表征了一种碱性蛋白酶,该酶来自Bacillus licheniformis A10,最适pH为9.0,最适温度为70 ℃,在Mg2+、Mn2+、K+存在下,酶活性显著增强。Kalwasińska等[13]从Bacillus luteus H11纯化出的碱性丝氨酸蛋白酶表现出极强的盐碱耐受性,对一些有机溶剂、表面活性剂等表现出很强的稳定性,这些性质在工业应用中是极具优势的。不仅地衣芽孢杆菌、枯草芽孢杆菌、解淀粉芽孢杆菌等细菌能产生碱性蛋白酶[14],各种真菌如黄曲霉、禾谷镰孢菌、青霉菌、黑曲霉和尖孢镰刀菌也被认为是丝氨酸蛋白酶的良好来源[15]。如Kiran[16]从土壤中分离和表征了几种黄曲霉菌株,并通过固态发酵优化其生产能力,与细菌相比其菌丝体容易分离,具有更低的成本效益,且在37 ℃下发酵3 d后获得最大的酶产量 13.73 μg/mL。丝氨酸蛋白酶的相对分子质量一般为15~40 kDa,一般在中性或偏碱性条件下发挥最大催化活性(表1),不断发掘优势菌株并探究酶的理化特性有助于提高其在食品工业中的实际应用。
表 1 微生物源丝氨酸蛋白酶的酶学性质及应用Table 1. Characteristics and application of microbial serine proteases菌株 分子量
(kDa)最适温度
(℃)最适pH 抑制剂 激活剂 应用范围 参考文献 Pediococcus
acidilactici NCDC 252937.1 37 8.5 Soybean trypsin inhibitor、Benzamide、β-ME / 具有耐酸性和抗氧化活性,用于乳制品生产 [14] Bacillus licheniformis A10 40.55 70 9.0 Fe3+、Ni2+、Zn2+、Ag+、Co2+、PMSF、EDTA、Urea、Formaldehyde、Benzene、Toluene、Xylene、Ethanol、Acetone、DMSO、SDS、Tween-20、Tween-80、H2O2 Mg2+、Mn2+、K+、Methanol、Sulfonylfluoride、β-ME 热稳定性好,用于食品工业生产中的高温环境 [12] Bacillus luteus H11 37 45 10.5 Fe2+、Cu2+、Zn2+、DFP、PMSF、Butanol、Methanol、Ethanol、Acetone、SDS 2M Methanol、Ca2+、Mg2+ 对盐和碱具有高度耐受性,用于海产品加工 [13] Geobacillus
Stearothermophilus28.3 50 8.5 Cu2+、Zn2+、Ni2+、Co2+、Mn2+、Ca2+、Na+、PMSF Mg2+ 通过蛋白酶水解、微生物发酵生产乳清蛋白水解物 [17] Planococcus bacterium 35~40 35 11.0 Ca2+、Co2+、EDTA、PMSF、SDS Mg2+、Cu2+、Methanol 水解银鱼产生大量的小分子肽,用于生物活性肽领域 [17] Bacillus amyloliquefaciens FCF-11 18.2 40 8 PMSF、EDTA Fe2+、Mn2+ 降解纤维蛋白,是有益的血栓溶解剂 [18] 注:Soybean trypsin inhibitor:大豆蛋白酶抑制剂;Benzamide:苯甲酰胺;β-ME:β-mercaptoethanol,β-巯基乙醇;PMSF:Phenylmethyl sulfonylfluoride,苯甲基磺酰氟化物;EDTA:Ethylene,乙烯; Diamine Tetraacetic Acid:乙二胺四乙酸;Urea:尿素;Formaldehyde:甲醛;Benzene:苯;Toluene:甲苯;Xylene:二甲苯;Ethanol:乙醇;Acetone:丙酮;DMSO:Dimethyl sulfoxide,二甲基亚砜;SDS:Sodium dodecyl sulfate,十二烷基硫酸钠;Tween:吐温;Sulfonylfluoride:氟化磺酰;DFP:Diisopropylfluorophosphate,二异丙基氟磷酸;PMSF:Phenylmethyl sulfonylfluoride,苯甲基磺酰氟化物;Butanol:丁醇;Methanol:甲醇;Acetone:丙酮;表2~表4同。 1.1.2 半胱氨酸蛋白酶及酶学特性
半胱氨酸蛋白酶(Cysteine protease,Cys,EC 3.4.22)广泛存在于菠萝茎、无花果、木瓜等植物中,微生物来源主要有米曲霉和孢子菌等真菌物种[19]。其相对分子量为40~55 kDa,蛋白水解活性的最适pH为6.0~8.0,最适温度为50~70 ℃,对氧化剂、p-CMB(p-chloromercuribenzene sulfonate,氯汞苯磺酸)等巯基化合物具有高度敏感性,但金属螯合剂对其活性没有显著性影响。Keshapaga等[20]从Bacillus subtilis A4中分离出的半胱氨酸蛋白酶在pH6.0~9.0显示出较高的比活性,最适pH为7.5,对pH和温度的耐受性使其在发酵过程中极为有利,可作为一种生态友好的微生物源半胱氨酸蛋白酶。
1.1.3 天冬氨酸蛋白酶及酶学特性
天冬氨酸蛋白酶(Aspartic proteases,Aps,EC 3.4.23)俗称酸性蛋白酶,是依赖天冬氨酸残基产生催化活性的内肽酶,其活性位点内具有两个天冬氨酸残基(Asp 32和Asp 215),通过激活水分子来切割肽键,这有利于提高其催化活性。微生物来源有黑曲霉、青霉、米曲霉、米黑毛霉等毛霉属、曲霉属和酵母等[21],但很少存在于细菌中,且真菌产生的酶与细菌相比具有广泛的底物特异性。Purushothaman等[22]从Aspergillus niger中分离纯化的天冬氨酸蛋白酶最适pH为3.5,最适温度为60 ℃,具有较好的耐热性,且比酶活达到40000±1800 U/mg。Wei等[23]利用Pichia pastoris异源表达了Aspergillus niger天冬氨酸蛋白酶,其总酶活和比酶活分别达到9412 U/mg和4852 U/mg。大多数天冬氨酸蛋白酶在低pH(pH3~4)下显示出最大活性,并且在pH3.0~4.5范围内具有等电点,最适温度为30~60 ℃,分子量通常为30~45 kDa,被胃蛋白酶抑制剂抑制[24](表2)。
表 2 微生物源天冬氨酸蛋白酶的酶学性质及应用Table 2. Characteristics and application of microbial aspartate proteases菌株 分子量
(kDa)最适温度
(℃)最适 pH 抑制剂 激活剂 应用范围 参考文献 Aspergillus niger 50 50 3.0 Pepstatin A / 在制备蛋白质水解物、生物活性肽方面具有应用前景 [23] Aspergillus glaucus MA0196 42.2 30 3.8 / / 对血红蛋白具有水解和脱色作用,在低水分条件下水解金枪鱼中的血红素蛋白 [25] Penicillium sp. 45 30 3.0 Co2+、Pb2+、Fe3+、PMSF、EDTA、Pepstatin A Mn2+、Cu2+ 水解肌红蛋白、血红蛋白等动物蛋白,提高肉制品中胶原蛋白的提取效率 [26] Phanerochaete chrysosporium 38 50 4.5 EDTA、IAA、PMSF、Pepstatin A、Co2+、Cu2+、AlCl3 KCl 用于奶酪生产,具有替代传统凝乳酶的潜力 [27] Aspergillus oryzae 40 60 5.0 Pepistatin A、Ni2+、Zn2+、Cu2+、COSO4 Ca2+、Mg2+、Fe2+、Mn2+、Mg2+、Mn2+ 在酸性pH下具有活性,适合奶酪生产 [28] Trichoderma reesei 40.7 50 4.0 SPH、WPH、DMF、DMSO、Cu2+、Mn2+、Pepstatin A EDTA 用于食品蛋白水解,能有效水解酸性乳清 [29] Rhizomucor miehei 37 55 5.5 Al3+、Cu2+、Co2+、Mn2+、Mg2+、Ca2+、K+、Ba2+、IAA、PMSF、EDTA、Pepstatin A SDS、CTAB、Urea、DTT 用于奶酪制作,是一种重要的凝乳酶替代物 [30] 注:Pepstatin A:胃蛋白酶抑制素;IAA:C10H9NO2,吲哚-3-乙酸;SPH:Soy protein hydrolysate,大豆蛋白水解物;WPH:Whey protein hydrolysates,乳清蛋白水解物;DMF:C3H7NO,N,N-二甲基甲酰胺;DTT:Dithiothreitol,二硫苏糖醇;表3~表4同。 1.1.4 金属蛋白酶及酶学特性
金属蛋白酶(Mental proteinase,MPs,EC 3.4.24)是一类需要金属离子(通常是锌、钴、镍和锰)作为辅助因子的酶。芽孢杆菌、放线菌、弧菌、沙雷氏菌属等都是金属酶的良好生产者[31]。不同微生物来源的金属蛋白酶酶学性质和实际应用有所差别,部分金属蛋白酶对低温、高温、有机溶剂和碱性环境具有极高的耐受性(表3)。如Zhang等[32]从Vibrio sp. LA-05菌株发酵上清液中分离纯化的金属蛋白酶VMP具有一定的耐盐性;李彬彬[33]从Thermoactinomyces vulgaris CDF得到的金属蛋白酶TvM6具有较好的热稳定性,在55 ℃高温下处理8 h仍可以保持80%活性。多数芽孢杆菌源的金属蛋白酶在凝乳方面具有独特优势,可以特异性水解酪蛋白[34],如Bacillus amyloliquefaciens GSBa-1[35]来源的金属蛋白酶主要水解κ-CN中Lys111-Lys112位点、Bacillus velezensis YH-1[36]来源的金属蛋白酶可在Lys21-Ile22和Tyr30-Val31位点水解酪蛋白、Bacillus velezensis DB219金属蛋白酶[37]主要水解κ-CN中Thr124-Ile125和Ser104-Phe105位点。这类酶主要是中性蛋白酶,最佳pH为5.0~9.0,最适温度为65~85 ℃,分子量为19~37 kDa[38]。
表 3 微生物源金属蛋白酶的酶学性质及应用Table 3. Characteristics and application of microbial metalloproteinases菌株 分子量
(kDa)最适温度
(℃)最适pH 抑制剂 激活剂 应用范围 参考文献 Marine Vibrio sp. LA-05 35 45 8.0 DTT、IAA、EDTA、EGTA、Ca2+、Mg2+、Sr2+、Ag+、Co2+、Zn2+、Mn2+、Fe2+、Ni2+、Pb2+、Al3+、Cr3+、Cu2+、Hg2+ Ba2+ 可应用于高盐海食品,多肽合成及鱼肉加工 [32] Thermoactinomycesvulgaris CDF 90 65 7.5 Na+、EDTA Ca2+ 具有较好的热稳定性,适合高温的食品加工环节 [33] Bacillus amyloliquefaciens
GSBa-138 57 5.5 EDTA、Cu2+、Cd2+、SDS、Urea、DMSO K+、Na+、Li+、Zn2+、Mg2+、Ca2+ 水解κ-CN Lys111-Lys112位点,可作为奶酪生产中的凝乳剂 [35] Bacillus velezensis YH-1 56.8 55 5 EDTA、Na+、Ca2+ / 水解κ-CN Lys21-Ile22和Tyr30-Val31位点,可增加凝乳柔韧性和粘度,加快凝固过程 [36] Bacillus velezensis DB219 36 75 5.5 EDTA、Mn2+、Na+、Al3+、Cu2+、Fe3+ Li+、Mn2+、Co2+ 水解κ-CN Thr124-Ile125和Ser104-Phe105位点,是一种
极具潜力的新型凝乳酶[37] Termitomyces clypeatus MTCC 5091 29 45 5.0 EDTA / 用于制作奶酪 [39] Paenibacillus spp.BD3526 35 65 / EDTA / 裂解κ-CN Met106-Ala107位点,奶酪生产中具备有利特质 [34] Bacillus subtilis 42 70 6.0 Zn2+、Fe2+、Cu2+、EDTA Ca2+、Al3+ 对牦牛乳酪蛋白有高度专一性,用于牦牛奶酪工业 [40] 注:EGTA:glycol-bis-(2-aminoethylether)-N,N,N′,N′-tetraacetic acid,乙二醇双α-氨基乙基醚四乙酸;κ-CN:κ-酪蛋白。 1.2 微生物源蛋白酶在食品工业中的应用
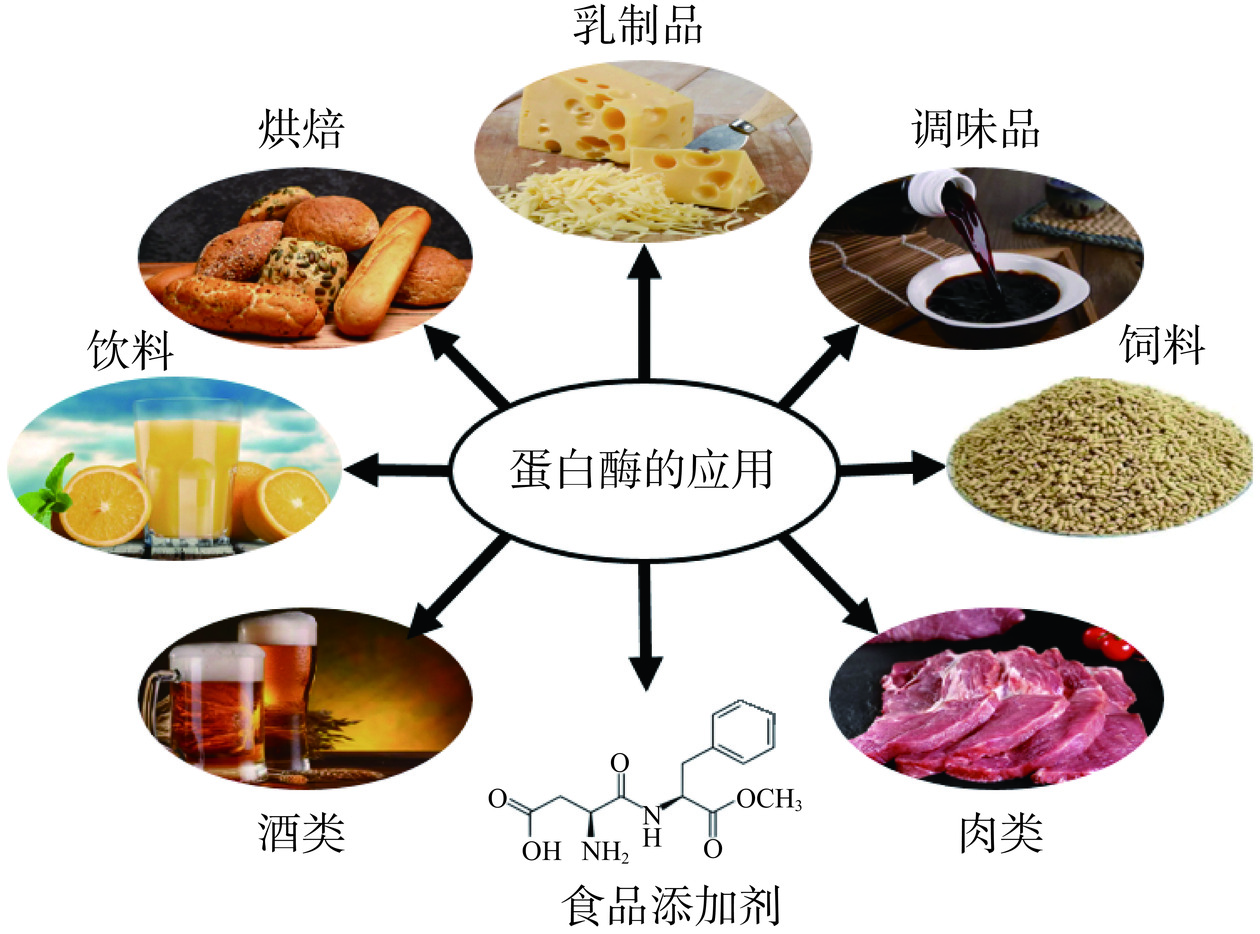
蛋白酶在食品加工中主要用于谷物糖化、酿造、生产蛋白质水解物、制作干酪和改变面团弹性等,可作为添加剂改善食品和饲料的营养品质,水解肉类中胶原蛋白使其更适合食用、减少混浊复合物果汁和酒精饮料沉淀;在酿酒工业中,蛋白酶水解大麦和麦芽产生糖和氨基酸,影响酒类的风味和香气[41]。图1列举了部分微生物源蛋白酶在肉类、乳制品、焙烤食品和豆制品等中的应用实例。
1.2.1 肉类中的应用
蛋白酶可以水解肉类结缔组织中的胶原蛋白和弹性蛋白,破坏它们分子结构,从而提高肉的嫩度,同时产生小分子肽改善肉的风味,目前微生物源蛋白酶在肉类嫩化方面研究较少[42],其中主要以天冬氨酸蛋白酶为主[43]。薛意斌等[44]在毕赤酵母中表达天冬氨酸蛋白酶基因AjproA1并探究其对猪肉嫩化的影响,经AjproA1处理的猪肉蒸煮损失率(19.55%)低于木瓜蛋白酶(22.73%)。并且其持水力增加、剪切力明显下降,表明AjproA1蛋白酶具有作为嫩肉剂的潜力。Lim等[25]从Aspergillus glaucus中获得的天冬氨酸蛋白酶在低水分条件下能水解金枪鱼干肉中的血红素蛋白,可以使鱼肉嫩化同时有脱色效果。研究发现,从Bacillus subtilis A4[20]中分离的半胱氨酸蛋白酶也可以使水牛肉嫩化而不会影响其稠度。在肉类嫩化方面,目前主要以植物源蛋白酶为主,微生物源蛋白酶研究较少,且蛋白酶对肉类嫩化机制方面的研究尚不清晰。但目前的研究显示微生物源蛋白酶在肉类嫩化方面具有较大研究潜力。
1.2.2 烘焙食品中的应用
蛋白酶通过水解蛋白质改性面包中的小麦面筋,降低面粉筋度、提高面筋可塑性、增大面团体积、改善面团韧性、缩短面团成形时间等,其降解产生的氨基酸与多肽能够促进香味物质形成。Aspergillus usamii来源的酸性蛋白酶[45]水解小麦面筋从而提高其溶解度,而小麦麸质的酶水解导致乳化活性指数(EAI)、水分和持油能力大幅提高。张依洁等[46]利用风味蛋白酶研制出具有高氨基酸含量和独特风味的鸡骨泥风味饼干。Renzetti等[47]以糙米面糊为原料制作糙米粉面包时,研究了蛋白酶(主要是中性蛋白酶)对面包质构和烘烤性能的影响,与未经处理的面包相比,无麸质米面包比体积增加30%~60%,面包硬度降低10%~30%[48],结果表明蛋白酶能够有效降低面糊粘度,提高面糊弹性和稳定性,对于面包制作具有积极影响。在焙烤食品中,添加蛋白酶使面包色泽得到改善同时还增加了面包香味,但其添加量及与其他酶的协同增效作用还需进一步探究。
1.2.3 乳制品中的应用
研究表明由芽孢杆菌产生的蛋白酶可以水解牛奶中的κ-酪蛋白,联合使用其它蛋白酶可使牛奶中的寡肽水解,加快奶酪成熟的同时产生特异风味物质,其水解产生的生物活性肽可用于开发功能性食品。在几类蛋白酶中,天冬氨酸蛋白酶因其在特定pH和温度范围内的高效性,通常被认为最适合应用于奶酪行业。且不同来源天冬氨酸蛋白酶可以特异性水解κ-酪蛋白,产生不同的小分子肽段,这些丰富的小分子肽增加了乳制品营养,有益于人体健康。如Phanerochaete chrysosporium[27]和Rhizomucor miehei[30]等产生的天冬氨酸蛋白酶可作为传统凝乳酶替代品,应用于奶酪生产。在乳制品行业,过去使用的凝乳酶均取自小牛,由于其生产周期长、产量低、价格高、人文道德等因素的制约,微生物源凝乳酶逐渐受到人们的关注。但在自然环境下筛选的微生物源凝乳酶在凝乳过程中易产生苦味物质,影响奶酪口感和得率。因此,通过分子生物学手段改造蛋白酶以适应工业化生产和实际应用是至关重要的。
1.2.4 豆制品中的应用
自古以来,蛋白酶广泛用于酱油和其他豆制品的制作,真菌来源的碱性和中性蛋白酶在酱油的加工中起关键作用。Song等[49]利用黑曲霉表达的天冬氨酸蛋白酶水解大豆分离蛋白,获得大量疏水性氨基酸,抑制了水解产物的苦味,表明蛋白酶在蛋白质水解物中具有改善风味的潜质。此外,在酱油酿造过程中,米曲霉、大豆曲霉和黑曲霉分泌的蛋白酶作用于原料大豆中的蛋白质,将其分解成小分子肽和氨基酸等风味物质,赋予酱油独特的香味[50]。Chen等[51]通过在豆制品中添加六种外源蛋白酶探究其对酱油的影响,发现中性蛋白酶与米曲霉粗蛋白酶单独和协同作用均表现很好,添加中性蛋白酶使氨基酸氮和总氮含量增加了7.45%和2.21%,提高了酱油蛋白的利用率和发酵质量。以大豆和麸皮为原料发酵酱油时,菌种质控和制曲工艺是影响原料利用率的关键,决定酱油的质量。
2. 微生物源脂肪酶
脂肪酶(Lipase,EC 3.1.1.3)是一类催化脂类水解的酶,其天然功能是水解甘油三酯中的酯键[52],催化脂质在水性或非水性条件下酯化,具有广泛的底物特异性[53]。根据脂肪酶的底物特异性分为脂肪酸特异性、位置特异性和立体特异性;依据脂肪酶来源不同分为动物脂肪酶、植物脂肪酶和微生物脂肪酶。与动植物脂肪酶相比,细菌和真菌脂肪酶的主要优点是具有较高的耐热性和有机溶剂耐受性[54],且大多数微生物源脂肪酶是胞外分泌型,方便进行分离提取,可应用于大规模生产[55]。如表4所示,脂肪酶的微生物来源包括芽孢杆菌属、伯克霍尔德氏菌属、假单胞菌属、葡萄球菌属、曲霉属、毛霉菌属、青霉属、根霉属、毕赤酵母和红酵母等[56]。作为绿色生物催化剂,脂肪酶不仅能提高产品稳定性、增加溶解度和延长保质期,还能改善产品质量,在食品工业中应用广泛。
表 4 微生物源脂肪酶的酶学性质及应用Table 4. Characteristics and application of microbial lipases菌株 分子量
(kDa)最适温度
(℃)最适pH 抑制剂 激活剂 用途 参考文献 Geobacillus kaustophilus DSM7263 43 50 8.0 Mg2+、Na+、Zn2+、PMSF、DTT、β-ME、SDS、DMSO、Ethanol、2-Propanol Ca2+ 生产生物发酵剂和风味
化合物[64] Rhizomucor miehei NRRL5282 55 40 7.0 Hg2+、Mn2+、Zn2+、Cu2+、NBS、SDS、Hexanol、Butanol、Isopropanol、butanol、Isopentanol、hexanol Mg2+、Na+ 增强食品风味 [65] Bacillus halodurans 42 60 9.0 Ba2+、Fe2+、Pb2+、DEPC、EDAC、PMSF Cd2+、Ni2+、Fe3+、DTNB 具有热稳定性,生产PUFA
食品[66] Serratia liquefaciens / 45 8.0 Zn2+、Fe2+、EDTA、SDS Ca2+、Mg2+、Tween-20 热稳定性好,用于食品中高温环节 [67] Candida viswanathii 69 45 4.0 Co2+、Hg2+、PMSF、EDTA、SDS、Tween-20、Tween-80、Sodium deoxycholate Mn2+、DTT、β-ME 在酸性pH下的活性和稳定性高,能水解卵磷脂 [68] Streptomyces bacillaris 31 45 9.0 Zn2+、Ni+、EDTA、TritonX-100、Tween-80、Tween-20、SDS Mg2+、Ba2+、Ca2+、K+、Span-80 规模化生产游离虾青素,可制备虾青素酯 [69] Bacillus
licheniformis NCUCS-527 40 8.0 CTAB、Mn2+、Cu2+、Co2+、Sn2+、Methanol、Ethanol、Acetonitrile、Acetone、Ethyl acetate、Tetrahydrofuran、Pyridine、Urea、DTT、EDTA、NaF、Na3VO4、PMSF、H2O2 Mg2+、K+、Na+、Ca2+、Al2+、Fe3+、Glycerol、Tween-20、CTAB 具有良好的耐寒性和耐碱性,用于生产中极端环境 [61] Halocynthiibacter arcticus 25.3 20 8.0 10%Ethanol、Triton X-100、SDS、Urea Na+ 通过酶促酯交换产生风味化合物 [62] L. plantarum B2-5、L. helveticus H1-6、W. confusa T1-3 / 40 6.0 Cu2+、Al3+、Fe3+、EDTA Na+、Fe2+、Mn2+ 产生的酯类物质赋予奶酪花果香 [70] W. confusa T1-5 / 35 6.0 Cu2+、Al3+、Fe3+、EDTA Na+、Fe2+、Mn2+ 丰富奶酪中优良的酸类风味物质 [70] Pseudoalteromonas lipolytica SCSIO 04301 80 45 8.0 Zn 2+、Ni 2+、Co 2+、SDS、Tween-60、Tween-80 Na+、DMSO、Ethanol、Acetone 生产EPA和优质奶酪 [71] Aeromicrobium sp. SCSIO 25071 30 35 7.5 Acetonitrile、n-Hexanol 30%DMSO、30%Ethanol、30%Acetone 具有冷活性和有机溶剂耐受性,用作食品添加剂 [72] 注:2-Propanol:异丙醇;NBS:N-Bromosuccinimide,N-溴代琥珀酰亚胺;Butanol:丁醇;Isopropanol:异丙醇;DEPC:Diethyl pyrocarbonate,焦碳酸二乙酯;EDAC:碳二亚胺盐酸盐;Sodium deoxycholate:脱氧胆酸钠;TritonX-100:聚乙二醇辛基苯基醚;Glycerol:丙三醇;Span-80:司盘80;CTAB:十六烷基三甲基溴化胺;Acetonitrile:乙腈;Acetone:丙酮;Ethyl acetate:乙酸乙酯;Tetrahydrofuran:四氢呋喃;Pyridine:吡啶;n-Hexanol:正己醇;DTNB:5,5’-Dithiobis-(2-nitrobenzoic acid),5,5’-二硫代双(2-硝基苯甲酸);PUFA:polyunsaturated fatty acid,多不饱和脂肪酸。 2.1 微生物源脂肪酶及酶学特性
在商业应用中,通常使用细菌和真菌源脂肪酶[57],它们大多数为碱性脂肪酶,分子量在25~60 kDa,最适pH为4.0~9.0,最适温度为30~60 ℃[58]。此外,由于大多数真菌源脂肪酶具有耐热性、pH稳定性、底物特异性和有机溶剂耐受性和提取成本低等优势,被认为是最好的脂肪酶来源之一[59]。
脂肪酶工业应用的一个挑战是寻找具有独特性质的新型脂肪酶,如耐有机溶剂、高盐、碱性条件和低温下的高活性酶。目前,研究人员已经从嗜极微生物中鉴定和表征了许多脂肪酶(表4),这些酶能够满足在极端反应环境的工业需求[60]。如Zhao等[61]从香樟种仁腐败物中分离出一株Bacillus licheniformis NCUCS-5,并对其胞外脂肪酶进行纯化,该脂肪酶具有良好耐寒性,在碱性条件和金属离子存在下具有较高活性,对各种有机溶剂、抑制剂和表面活性剂有较强的耐受性。Le等[62]对Halocynthiibacter arcticus分泌的一种适冷型脂肪酶进行催化特性和功能研究,发现该酶在低温下具有高活性,广泛的底物特异性,较高的稳定性和催化效率。Gao等[63]发现Paenibacillus来源的脂肪酶对一些水不溶性有机溶剂具有高度耐受性。
目前的报道主要聚焦于各种脂肪酶菌株的筛选、现有脂肪酶基因的修饰、修饰脂肪酶功能分析、广泛适用性等。天然宿主脂肪酶存在表达效率低、热稳定性差等问题,限制了其工业化生产。针对特定宿主开发出高效的遗传方法,如大肠杆菌作为目前可靠的表达系统之一,由于其高水平的重组蛋白表达能力而为大规模生产外源蛋白提供了可能。Gao等[63]从Paenibacillus pasadenensis菌株中发现了一个新的脂肪酶基因,将脂肪酶基因序列克隆到pET-28a表达载体中,用IPTG在20 ℃下诱导后,在E. coli BL21(DE3)中获得脂肪酶的高水平表达,重组脂肪酶活性比野生型高约1631倍。Ozdemir等[64]将嗜热菌Geobacillus kaustophilus的脂肪酶基因gklip克隆到pET-28a载体中,并在其N端添加6×His标签,重组gklip基因在E. coli BL21(DE3)细胞成功异源表达,且对底物特异性水解活性显著增加。这些研究为脂肪酶在食品工业生产中的应用提供有力支撑。
2.2 微生物源脂肪酶在食品工业中的应用
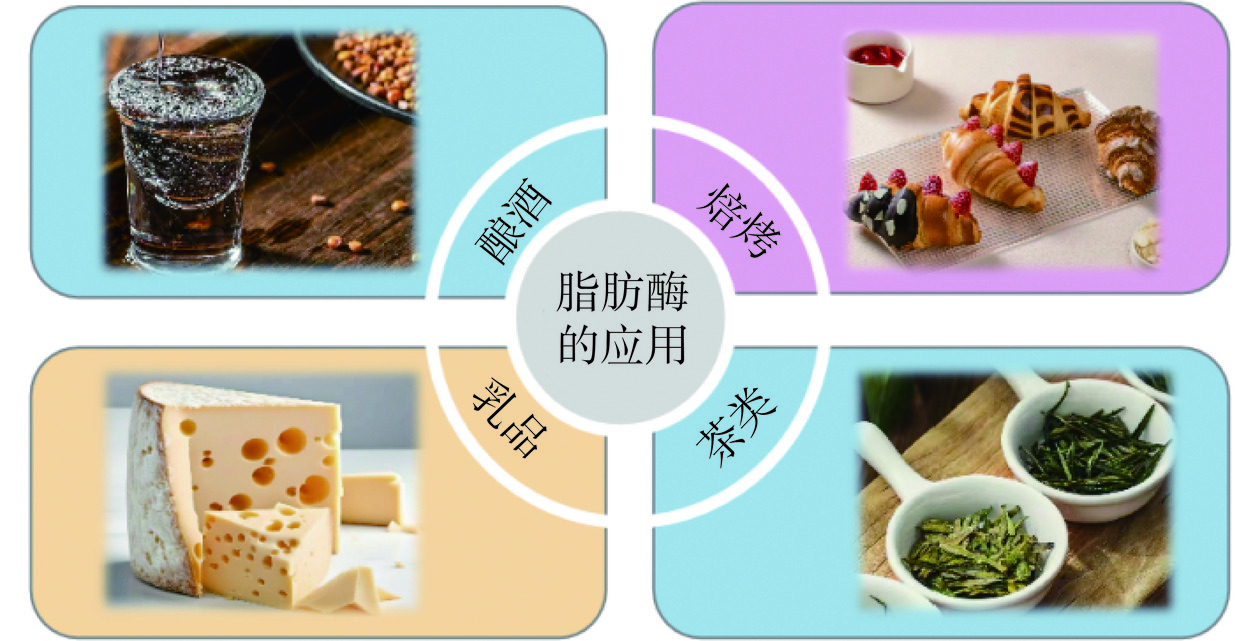
微生物源脂肪酶已深入食品工业中多个领域,广泛用于面食加工、酒类酿造、乳制品生产、茶类增香、药食同源等食品加工中[72]。通过脂肪酶酯化、酯交换或酯间反应可以获得一系列具有工业价值的风味酯,可应用于肉类、奶酪、面包、茶叶和饮料中[73],增加食品风味(图2)。随着食品工业快速发展,微生物源脂肪酶的应用也越来越广泛。以下主要论述微生物源脂肪酶在酒类酿造、面食加工、乳制品中、茶类增香中的应用。
2.2.1 酒类酿造中的应用
脂肪酶是一种同时具有酯分解、酯交换和酯合成能力的酶,在酒类酿造中,它不仅可以加快发酵进程,促进酯类化合物的合成以增强风味,还可以缩短贮存老熟的时间,提高出酒率[74]。翟磊等[75]对不同发酵阶段大曲样品进行产脂肪酶菌种的筛选与鉴定发现,产脂肪酶微生物是形成特殊风味物质的关键,这一发现为产脂肪酶提供优势菌株,同时为芝麻香型白酒香气成分解析奠定理论基础。李站胜等[76]在浓香型白酒糟中加入南极假丝酵母脂肪酶B,研究发现当酶添加量1.5‰(w/w),20 ℃下反应3 d时增酯增香的效果最好,浓香型白酒的主要呈香物质乙酸乙酯和己酸乙酯分别较空白组提高了16.38倍和16.67倍,表明脂肪酶对催化酒糟,扩大其资源化利用具有重要意义。蔡琪琪[77]通过分子生物学手段对红曲霉在黄酒酿造过程中的作用进行研究,结果发现红曲霉产生的脂肪酶可以促进酿酒酵母的酒精发酵,且能有效提高发酵前期乙酸酯类和乳酸乙酯的含量,对黄酒风味的形成起着至关重要的作用。在酒类酿造中,微生物源脂肪酶主要通过促进酯类化合物合成起到增香的作用,但具体作用机制尚不明确。
2.2.2 面食加工中的应用
脂肪酶属于面团强化用酶,能增强面团的稳定性,脂肪酶通过水解作用形成更强的极性和亲水结构,与麦谷蛋白更好地结合,形成更强的面筋网络,用于改善面包柔软度,增加面包膨胀体积等。Colakoglu等[78]通过流变特性和热分析发现,脂肪酶能提高面团稳定性、最大的抗拉伸性和硬度以及降低软化程度和粘性,极大程度改善了面团的处理性能。钱忠英等[79]探究了重组疏棉状嗜热丝孢菌脂肪酶在面包体系中的使用效果,脂肪酶在增大面包比容、强化面筋结构和提高感官整体接受度等方面仅次于商业脂肪酶,优于化学改良剂双乙酰酒石酸单(双)甘油酯,且当面包配方中加入15 mg/kg上述脂肪酶能明显增大面包比容(9.8%),降低面包硬度,气味和组织结构的感官得分最高。Huang等[80]旨在探究脂肪酶处理的乳脂对面包质量的影响,脂肪酶处理增强了面团和面包的质地,且体外消化显示,脂肪酶处理乳脂的面包的血糖负荷明显低于对照样品,说明脂肪酶不仅可以改善面包的功能,而且能降低产品的血糖反应。在焙烤食品中,添加脂肪酶可以改变面团特性。油脂产生的游离脂肪酸进一步水解、氧化生成过氧化物和氢过氧化物,过氧化物继续分解成醛类、酮类化合物、其他氧化物等,引起油脂酸败。因此,脂肪酶添加量在烘培食品制作中尤为重要。
2.2.3 乳制品中的应用
脂肪酶可以应用于乳酯水解,包括奶酪和奶粉风味增强、奶酪熟化、奶油及冰淇淋酯解改性等,脂解过程即甘油三酯酶促反应生成甘油、脂肪酸、甘油单酯或甘油二酯等,为不同风味奶酪制作与牛奶调味提供了新选择[81]。李晓楠[70]利用4株产脂肪酶乳酸菌作为发酵剂制作酸凝奶酪,结果表明实验组中奶酪氧化程度均较对照组低,且实验组奶酪中脂肪酸的种类和含量也更加丰富,质地、口感和风味都较对照组好。Zhang等[81]通过优化脂肪酶催化的水解和酯化工艺,提高复原脱脂乳制品的风味品质且降低了脂肪含量,提供了一种新的改变挥发性特征的方法,并以非合成和简化的方式改善了脱脂牛奶产品的感官质量。Omar等[82]探究了三种不同微生物源脂肪酶水解乳脂对风味的影响,结果表明这三种脂肪酶都能够产生短链脂肪酸(尤其是丁酸和己酸)和其他重要短链和中链脂肪酸(调味化合物),适用于多种乳制品生产。脂肪酶不仅可以通过酯化反应增加香气成分、丰富营养物质,对于脱脂乳生产也十分重要。
2.2.4 茶类增香中的应用
茶的质量取决于加工过程中脱水、机械切割和发酵程度,脂肪酶可减少茶叶中的脂质成分,增强多不饱和脂肪酸浓度,从而保证茶叶的优良风味[83]。儿茶素类化合物是茶汤中主要的多酚类化合物,对茶叶的品质及功效起着决定性作用,方洪枫等[84]通过混菌(冠突曲霉菌株PE-1和黑曲霉PE-4)产脂肪酶发酵绿茶,研究脂肪酶活性对酯型儿茶素的水解作用,表明该脂肪酶可以有效转化表没食子儿茶素没食子酸酯(EGCG)产生表儿茶素没食子酸酯(ECG),水解率高达94.46%。马超龙等[85]研究普洱茶叶加工过程中脂肪酶参与的后发酵过程,发现产生的不饱和脂肪酸不仅具有抗氧化活性,且脂肪酸氧化降解对茶叶香气影响至关重要。孙浩等[86]研究茶杆发酵(CETSM)和马铃薯葡萄糖发酵(CCEPDM)两种黑曲霉提取液对绿茶香气的影响,发现这些提取液中含有脂肪酶,通过感官评定、气质联用和嗅觉仪分析表明,与未经酶处理的对照相比,来自CETSM的黑曲霉发酵液对绿茶香气特征的影响较小,而CETSM的黑曲霉发酵液显著改变了绿茶的香气强度和挥发性成分。脂肪酶在茶类中的作用主要是增香,且生成的多不饱和脂肪酸可以增加营养。但发酵本身是一个复杂过程,无法确定具体香气成分,需要后续不断探究脂肪酶与香气成分的关系,以更好应用于茶类生产。
3. 结语与展望
蛋白酶和脂肪酶作为安全高效的生物酶制剂广泛应用于食品工业,且微生物源水解酶因获取便利、成本低廉、分子改造简便等优势成为研究热点。本文就食品工业中常用的微生物源水解酶-蛋白酶和脂肪酶进行综述,阐述其酶学特性及在肉类、烘焙食品、乳制品、豆制品、酒类、茶类等应用中的研究进展。通过对微生物源蛋白酶和脂肪酶分类讨论,充分了解各种蛋白酶和脂肪酶的适用性,提高其使用效率,为更好应用于食品工业生产提供理论参考。另外,随着世界人口增加和自然资源枯竭,迫切需要为食品工业化生产寻求高效、稳定和可持续使用的新酶资源,不同食品应用领域需要酶具有特定性质。因此,分子生物学技术也尤为重要,其可以使水解酶具有新的功能和稳定性,这也是未来研究的热点之一。充分挖掘优势菌株的独特性质、改造现有基因使其适用食品工业环境,不仅为优质微生物源蛋白酶和脂肪酶开发提供技术支撑,也展望了微生物源水解酶在食品工业应用中的广阔前景。
-
表 1 微生物源丝氨酸蛋白酶的酶学性质及应用
Table 1 Characteristics and application of microbial serine proteases
菌株 分子量
(kDa)最适温度
(℃)最适pH 抑制剂 激活剂 应用范围 参考文献 Pediococcus
acidilactici NCDC 252937.1 37 8.5 Soybean trypsin inhibitor、Benzamide、β-ME / 具有耐酸性和抗氧化活性,用于乳制品生产 [14] Bacillus licheniformis A10 40.55 70 9.0 Fe3+、Ni2+、Zn2+、Ag+、Co2+、PMSF、EDTA、Urea、Formaldehyde、Benzene、Toluene、Xylene、Ethanol、Acetone、DMSO、SDS、Tween-20、Tween-80、H2O2 Mg2+、Mn2+、K+、Methanol、Sulfonylfluoride、β-ME 热稳定性好,用于食品工业生产中的高温环境 [12] Bacillus luteus H11 37 45 10.5 Fe2+、Cu2+、Zn2+、DFP、PMSF、Butanol、Methanol、Ethanol、Acetone、SDS 2M Methanol、Ca2+、Mg2+ 对盐和碱具有高度耐受性,用于海产品加工 [13] Geobacillus
Stearothermophilus28.3 50 8.5 Cu2+、Zn2+、Ni2+、Co2+、Mn2+、Ca2+、Na+、PMSF Mg2+ 通过蛋白酶水解、微生物发酵生产乳清蛋白水解物 [17] Planococcus bacterium 35~40 35 11.0 Ca2+、Co2+、EDTA、PMSF、SDS Mg2+、Cu2+、Methanol 水解银鱼产生大量的小分子肽,用于生物活性肽领域 [17] Bacillus amyloliquefaciens FCF-11 18.2 40 8 PMSF、EDTA Fe2+、Mn2+ 降解纤维蛋白,是有益的血栓溶解剂 [18] 注:Soybean trypsin inhibitor:大豆蛋白酶抑制剂;Benzamide:苯甲酰胺;β-ME:β-mercaptoethanol,β-巯基乙醇;PMSF:Phenylmethyl sulfonylfluoride,苯甲基磺酰氟化物;EDTA:Ethylene,乙烯; Diamine Tetraacetic Acid:乙二胺四乙酸;Urea:尿素;Formaldehyde:甲醛;Benzene:苯;Toluene:甲苯;Xylene:二甲苯;Ethanol:乙醇;Acetone:丙酮;DMSO:Dimethyl sulfoxide,二甲基亚砜;SDS:Sodium dodecyl sulfate,十二烷基硫酸钠;Tween:吐温;Sulfonylfluoride:氟化磺酰;DFP:Diisopropylfluorophosphate,二异丙基氟磷酸;PMSF:Phenylmethyl sulfonylfluoride,苯甲基磺酰氟化物;Butanol:丁醇;Methanol:甲醇;Acetone:丙酮;表2~表4同。  下载: 导出CSV
下载: 导出CSV
表 2 微生物源天冬氨酸蛋白酶的酶学性质及应用
Table 2 Characteristics and application of microbial aspartate proteases
菌株 分子量
(kDa)最适温度
(℃)最适 pH 抑制剂 激活剂 应用范围 参考文献 Aspergillus niger 50 50 3.0 Pepstatin A / 在制备蛋白质水解物、生物活性肽方面具有应用前景 [23] Aspergillus glaucus MA0196 42.2 30 3.8 / / 对血红蛋白具有水解和脱色作用,在低水分条件下水解金枪鱼中的血红素蛋白 [25] Penicillium sp. 45 30 3.0 Co2+、Pb2+、Fe3+、PMSF、EDTA、Pepstatin A Mn2+、Cu2+ 水解肌红蛋白、血红蛋白等动物蛋白,提高肉制品中胶原蛋白的提取效率 [26] Phanerochaete chrysosporium 38 50 4.5 EDTA、IAA、PMSF、Pepstatin A、Co2+、Cu2+、AlCl3 KCl 用于奶酪生产,具有替代传统凝乳酶的潜力 [27] Aspergillus oryzae 40 60 5.0 Pepistatin A、Ni2+、Zn2+、Cu2+、COSO4 Ca2+、Mg2+、Fe2+、Mn2+、Mg2+、Mn2+ 在酸性pH下具有活性,适合奶酪生产 [28] Trichoderma reesei 40.7 50 4.0 SPH、WPH、DMF、DMSO、Cu2+、Mn2+、Pepstatin A EDTA 用于食品蛋白水解,能有效水解酸性乳清 [29] Rhizomucor miehei 37 55 5.5 Al3+、Cu2+、Co2+、Mn2+、Mg2+、Ca2+、K+、Ba2+、IAA、PMSF、EDTA、Pepstatin A SDS、CTAB、Urea、DTT 用于奶酪制作,是一种重要的凝乳酶替代物 [30] 注:Pepstatin A:胃蛋白酶抑制素;IAA:C10H9NO2,吲哚-3-乙酸;SPH:Soy protein hydrolysate,大豆蛋白水解物;WPH:Whey protein hydrolysates,乳清蛋白水解物;DMF:C3H7NO,N,N-二甲基甲酰胺;DTT:Dithiothreitol,二硫苏糖醇;表3~表4同。
下载: 导出CSV
表 3 微生物源金属蛋白酶的酶学性质及应用
Table 3 Characteristics and application of microbial metalloproteinases
菌株 分子量
(kDa)最适温度
(℃)最适pH 抑制剂 激活剂 应用范围 参考文献 Marine Vibrio sp. LA-05 35 45 8.0 DTT、IAA、EDTA、EGTA、Ca2+、Mg2+、Sr2+、Ag+、Co2+、Zn2+、Mn2+、Fe2+、Ni2+、Pb2+、Al3+、Cr3+、Cu2+、Hg2+ Ba2+ 可应用于高盐海食品,多肽合成及鱼肉加工 [32] Thermoactinomycesvulgaris CDF 90 65 7.5 Na+、EDTA Ca2+ 具有较好的热稳定性,适合高温的食品加工环节 [33] Bacillus amyloliquefaciens
GSBa-138 57 5.5 EDTA、Cu2+、Cd2+、SDS、Urea、DMSO K+、Na+、Li+、Zn2+、Mg2+、Ca2+ 水解κ-CN Lys111-Lys112位点,可作为奶酪生产中的凝乳剂 [35] Bacillus velezensis YH-1 56.8 55 5 EDTA、Na+、Ca2+ / 水解κ-CN Lys21-Ile22和Tyr30-Val31位点,可增加凝乳柔韧性和粘度,加快凝固过程 [36] Bacillus velezensis DB219 36 75 5.5 EDTA、Mn2+、Na+、Al3+、Cu2+、Fe3+ Li+、Mn2+、Co2+ 水解κ-CN Thr124-Ile125和Ser104-Phe105位点,是一种
极具潜力的新型凝乳酶[37] Termitomyces clypeatus MTCC 5091 29 45 5.0 EDTA / 用于制作奶酪 [39] Paenibacillus spp.BD3526 35 65 / EDTA / 裂解κ-CN Met106-Ala107位点,奶酪生产中具备有利特质 [34] Bacillus subtilis 42 70 6.0 Zn2+、Fe2+、Cu2+、EDTA Ca2+、Al3+ 对牦牛乳酪蛋白有高度专一性,用于牦牛奶酪工业 [40] 注:EGTA:glycol-bis-(2-aminoethylether)-N,N,N′,N′-tetraacetic acid,乙二醇双α-氨基乙基醚四乙酸;κ-CN:κ-酪蛋白。
下载: 导出CSV
表 4 微生物源脂肪酶的酶学性质及应用
Table 4 Characteristics and application of microbial lipases
菌株 分子量
(kDa)最适温度
(℃)最适pH 抑制剂 激活剂 用途 参考文献 Geobacillus kaustophilus DSM7263 43 50 8.0 Mg2+、Na+、Zn2+、PMSF、DTT、β-ME、SDS、DMSO、Ethanol、2-Propanol Ca2+ 生产生物发酵剂和风味
化合物[64] Rhizomucor miehei NRRL5282 55 40 7.0 Hg2+、Mn2+、Zn2+、Cu2+、NBS、SDS、Hexanol、Butanol、Isopropanol、butanol、Isopentanol、hexanol Mg2+、Na+ 增强食品风味 [65] Bacillus halodurans 42 60 9.0 Ba2+、Fe2+、Pb2+、DEPC、EDAC、PMSF Cd2+、Ni2+、Fe3+、DTNB 具有热稳定性,生产PUFA
食品[66] Serratia liquefaciens / 45 8.0 Zn2+、Fe2+、EDTA、SDS Ca2+、Mg2+、Tween-20 热稳定性好,用于食品中高温环节 [67] Candida viswanathii 69 45 4.0 Co2+、Hg2+、PMSF、EDTA、SDS、Tween-20、Tween-80、Sodium deoxycholate Mn2+、DTT、β-ME 在酸性pH下的活性和稳定性高,能水解卵磷脂 [68] Streptomyces bacillaris 31 45 9.0 Zn2+、Ni+、EDTA、TritonX-100、Tween-80、Tween-20、SDS Mg2+、Ba2+、Ca2+、K+、Span-80 规模化生产游离虾青素,可制备虾青素酯 [69] Bacillus
licheniformis NCUCS-527 40 8.0 CTAB、Mn2+、Cu2+、Co2+、Sn2+、Methanol、Ethanol、Acetonitrile、Acetone、Ethyl acetate、Tetrahydrofuran、Pyridine、Urea、DTT、EDTA、NaF、Na3VO4、PMSF、H2O2 Mg2+、K+、Na+、Ca2+、Al2+、Fe3+、Glycerol、Tween-20、CTAB 具有良好的耐寒性和耐碱性,用于生产中极端环境 [61] Halocynthiibacter arcticus 25.3 20 8.0 10%Ethanol、Triton X-100、SDS、Urea Na+ 通过酶促酯交换产生风味化合物 [62] L. plantarum B2-5、L. helveticus H1-6、W. confusa T1-3 / 40 6.0 Cu2+、Al3+、Fe3+、EDTA Na+、Fe2+、Mn2+ 产生的酯类物质赋予奶酪花果香 [70] W. confusa T1-5 / 35 6.0 Cu2+、Al3+、Fe3+、EDTA Na+、Fe2+、Mn2+ 丰富奶酪中优良的酸类风味物质 [70] Pseudoalteromonas lipolytica SCSIO 04301 80 45 8.0 Zn 2+、Ni 2+、Co 2+、SDS、Tween-60、Tween-80 Na+、DMSO、Ethanol、Acetone 生产EPA和优质奶酪 [71] Aeromicrobium sp. SCSIO 25071 30 35 7.5 Acetonitrile、n-Hexanol 30%DMSO、30%Ethanol、30%Acetone 具有冷活性和有机溶剂耐受性,用作食品添加剂 [72] 注:2-Propanol:异丙醇;NBS:N-Bromosuccinimide,N-溴代琥珀酰亚胺;Butanol:丁醇;Isopropanol:异丙醇;DEPC:Diethyl pyrocarbonate,焦碳酸二乙酯;EDAC:碳二亚胺盐酸盐;Sodium deoxycholate:脱氧胆酸钠;TritonX-100:聚乙二醇辛基苯基醚;Glycerol:丙三醇;Span-80:司盘80;CTAB:十六烷基三甲基溴化胺;Acetonitrile:乙腈;Acetone:丙酮;Ethyl acetate:乙酸乙酯;Tetrahydrofuran:四氢呋喃;Pyridine:吡啶;n-Hexanol:正己醇;DTNB:5,5’-Dithiobis-(2-nitrobenzoic acid),5,5’-二硫代双(2-硝基苯甲酸);PUFA:polyunsaturated fatty acid,多不饱和脂肪酸。
下载: 导出CSV
-
[1] ERMIS E. Halal status of enzymes used in food industry[J]. Trends in Food Science & Technology,2017,64:69−73.
[2] TAVANO O L, BERENGUER‐MURCIA A, SECUNDO F, et al. Biotechnological applications of proteases in food technology[J]. Comprehensive Reviews in Food Science and Food Safety,2018,17(2):412−436. doi: 10.1111/1541-4337.12326
[3] THERON L W, DIVOL B. Microbial aspartic proteases:Current and potential applications in industry[J]. Applied Microbiology and Biotechnology,2014,98(21):8853−8868. doi: 10.1007/s00253-014-6035-6
[4] HE W M, YANG R J, ZHAO W. Effect of acid deamidation-alcalase hydrolysis induced modification on functional and bitter-masking properties of wheat gluten hydrolysates[J]. Food Chemistry,2019,277:655−663. doi: 10.1016/j.foodchem.2018.11.004
[5] TAKOI K, KOIE K, ITOGA Y, et al. Biotransformation of hop-derived monoterpene alcohols by lager yeast and their contribution to the flavor of hopped beer[J]. Journal of Agricultural and Food Chemistry,2010,58(8):5050−5058. doi: 10.1021/jf1000524
[6] JERMEN M, FASSIL A. The role of microbial aspartic protease enzyme in food and beverage industries[J]. Journal of Food Quality,2018,2018:7957269.
[7] SHU L, SI X G, YANG X D, et al. Enhancement of acid protease activity of Aspergillus oryzae using atmospheric and room temperature plasma[J]. Frontiers in Microbiology,2020,11:1418−1419. doi: 10.3389/fmicb.2020.01418
[8] GAO X L, LIU E M, YIN Y Y, et al. Enhancing activities of salt-tolerant proteases secreted by Aspergillus oryzae using atmospheric and room-temperature plasma mutagenesis[J]. Journal of Agricultural and Food Chemistry,2020,68(9):2757−2764. doi: 10.1021/acs.jafc.9b08116
[9] MURTHY P S, KUSUMOTO K I. Acid protease production by Aspergillus oryzae on potato pulp powder with emphasis on glycine releasing activity:A benefit to the food industry[J]. Food and Bioproducts Processing,2015,96:180−188. doi: 10.1016/j.fbp.2015.07.013
[10] SOLANKI P, PUTATUNDA C, KUMAR A, et al. Microbial proteases:Ubiquitous enzymes with innumerable uses[J]. 3 Biotech,2021,11(10):428−428. doi: 10.1007/s13205-021-02928-z
[11] RAZZAQ A, SHAMSI S, ALI A, et al. Microbial proteases applications[J]. Frontiers in Bioengineering and Biotechnology,2019,7:110. doi: 10.3389/fbioe.2019.00110
[12] YILMAZ B, BALTACI M O, SISECIOGLU M, et al. Thermotolerant alkaline protease enzyme from Bacillus licheniformis A10:Purification, characterization, effects of surfactants and organic solvents[J]. Journal of Enzyme Inhibition and Medicinal Chemistry,2016,31(6):1241−1247. doi: 10.3109/14756366.2015.1118687
[13] KALWASIŃSKA A, JANKIEWICZ U, FELFÖLDI T, et al. Alkaline and halophilic protease production by Bacillus luteus H11 and its potential industrial applications[J]. Food Technology and Biotechnology,2018,56(4):553−561.
[14] BANSAL P, KUMAR R, SINGH J, et al. Production of extracellular alkaline serine protease from Pediococcus acidilactici NCDC 252:Isolation, purification, physicochemical and catalytic characterization[J]. Catalysis Letters,2021,151(2):324−337. doi: 10.1007/s10562-020-03331-8
[15] NOVELLI P K, BARROS M M, FLEURI L F. Novel inexpensive fungi proteases:Production by solid state fermentation and characterization[J]. Food Chemistry,2016,198:119−124. doi: 10.1016/j.foodchem.2015.11.089
[16] KIRAN ANAND K A. Fungal protease production by Aspergillus niger and Aspergillus flavus using rice bran as the substrate[J]. Academia Journal of Agricultural Research,2016,4(6):333−338.
[17] LENG W J, WU X Y, QI X H, et al. Systematic functional analysis and potential application of a serine protease from cold-adapted Planococcus bacterium[J]. Food Science and Human Wellness,2023,12(5):1751−1761. doi: 10.1016/j.fshw.2023.02.025
[18] KOTB E. Purification and partial characterization of a chymotrypsin-like serine fibrinolytic enzyme from Bacillus amyloliquefaciens FCF-11 using corn husk as a novel substrate[J]. World Journal of Microbiology & Biotechnology,2014,30:2071−2080.
[19] LIU N, REN W C, LI F J, et al. Involvement of the cysteine protease BcAtg4 in development and virulence of Botrytis cinerea[J]. Current Genetics,2019,65(1):293−300. doi: 10.1007/s00294-018-0882-0
[20] KESHAPAGA U R, JATHOTH K, SINGH S S, et al. Characterization of high-yield Bacillus subtilis cysteine protease for diverse industrial applications[J]. Brazilian Journal of Microbiology,2023,54(2):739−752. doi: 10.1007/s42770-023-00992-6
[21] XUE Y B, YAN Q J, LI X, et al. Characterization of a novel aspartic protease from Trichoderma asperellum for the preparation of duck blood peptides[J]. Applied Microbiology and Biotechnology,2024,108(1):131−131. doi: 10.1007/s00253-023-12848-y
[22] PURUSHOTHAMAN K, BHAT S K, SINGH S A, et al. Aspartic protease from Aspergillus niger:Molecular characterization and interaction with pepstatin A[J]. International Journal of Biological Macromolecules,2019,139:199−212. doi: 10.1016/j.ijbiomac.2019.07.133
[23] WEI M Y, CHEN P C, ZHENG P, et al. Purification and characterization of aspartic protease from Aspergillus niger and its efficient hydrolysis applications in soy protein degradation[J]. Microbial Cell Factories,2023,22(1):42. doi: 10.1186/s12934-023-02047-9
[24] SUMANTHA A, LARROCHE C, PANDEY A. Microbiology and industrial biotechnology of food-grade proteases:A perspective[J]. Food Technology and Biotechnology,2006,44(2):211−220.
[25] LIM L, SENBA H, KIMURA Y, et al. Influences of N-linked glycosylation on the biochemical properties of aspartic protease from Aspergillus glaucus MA0196[J]. Process Biochemistry,2019,79:74−80. doi: 10.1016/j.procbio.2018.12.017
[26] GUO Y J, LI X, JIA W, et al. Characterization of an intracellular aspartic protease (PsAPA) from Penicillium sp. XT7 and its application in collagen extraction[J]. Food Chemistry,2021,345:128834. doi: 10.1016/j.foodchem.2020.128834
[27] DA SILVA R R, DE OLIVEIRA L C G, JULIANO M A, et al. Biochemical and milk-clotting properties and mapping of catalytic subsites of an extracellular aspartic peptidase from Basidiomycete fungus Phanerochaete chrysosporium[J]. Food Chemistry,2017,225(15):45−54.
[28] MAMO J, KANGWA M, SUAREZ ORELLANA J F, et al. Purification and characterization of aspartic protease produced from Aspergillus oryzae DRDFS13 MN726447 under solid-state fermentation[J]. Catalysis Letters,2022,152(7):2033−2046. doi: 10.1007/s10562-021-03777-4
[29] AHMED U, PFANNSTIEL J, STRESSLER T, et al. Purification and characterization of a fungal aspartic peptidase from Trichoderma reesei and its application for food and animal feed protein hydrolyses[J]. Journal of the Science of Food and Agriculture,2022,102(12):5190−5199. doi: 10.1002/jsfa.11871
[30] DA SILVA R R, SOUTO T B, DE OLIVEIRA T B, et al. Evaluation of the catalytic specificity, biochemical properties, and milk clotting abilities of an aspartic peptidase from Rhizomucor miehei[J]. Journal of Industrial Microbiology and Biotechnology,2016,43(8):1059−1069. doi: 10.1007/s10295-016-1780-4
[31] 杭锋, 刘沛毅, 穆海菠, 等. 细菌来源金属蛋白酶的研究进展[J]. 工业微生物,2015,45(6):53−59. [HANG F, LIU P Y, MU H B, et al. Advance in recent research on bacterial metalloproteases[J]. Industrial Microbiology,2015,45(6):53−59.] doi: 10.3969/j.issn.1001-6678.2015.06.010 HANG F, LIU P Y, MU H B, et al. Advance in recent research on bacterial metalloproteases[J]. Industrial Microbiology, 2015, 45(6): 53−59. doi: 10.3969/j.issn.1001-6678.2015.06.010
[32] ZHANG H, LI H, LIU H, et al. The application of a halotolerant metalloprotease from marine bacterium Vibrio sp. LA-05 in liquid detergent formulations[J]. International Biodeterioration & Biodegradation,2019,142:18−25.
[33] 李彬彬. 普通高温放线菌 CDF 的金属蛋白酶 TvM6 的活化机制及性质研究[D]. 武汉:武汉大学, 2019. [LI B B, Activation and enzymatic property of a metalloproteinase (TvM6) from Thermoactinomyces vulgaris CDF[D]. Wuhan:Wuhan University, 2019.] LI B B, Activation and enzymatic property of a metalloproteinase (TvM6) from Thermoactinomyces vulgaris CDF[D]. Wuhan: Wuhan University, 2019.
[34] HANG F, WANG Q B, HONG Q, et al. Purification and characterization of a novel milk-clotting metalloproteinase from Paenibacillus spp. BD3526[J]. International Journal of Biological Macromolecules,2016,85:547−554. doi: 10.1016/j.ijbiomac.2016.01.028
[35] ZHAO X, CAI M, YANG Z J, et al. Purification and characterization of a novel milk-clotting enzyme produced by Bacillus amyloliquefaciens GSBa-1[J]. European Food Research and Technology,2019,245(11):2447−2457. doi: 10.1007/s00217-019-03361-6
[36] WANG Y H, AZIZ T, HU G G, et al. Production, purification, and characterization of a novel milk-clotting metalloproteinase produced by Bacillus velezensis YH-1 isolated from traditional Jiuqu[J]. LWT,2023,184:115080. doi: 10.1016/j.lwt.2023.115080
[37] ZHANG Y, HU J Y, WANG J X, et al. Purification and characteristics of a novel milk-clotting metalloprotease from Bacillus velezensis DB219[J]. Journal of Dairy Science,2023,106(10):6688−7000. doi: 10.3168/jds.2023-23450
[38] 李鑫淳. 向日葵锈菌金属蛋白酶基因的生物信息学分析及其原核表达[D]. 呼和浩特:内蒙古农业大学, 2019. [LI X C, Bioinformatics analysis and prokaryotic expression of mentaiioproteinnase gene from Pucciniahelianthi schw[D]. Hohhot:Inne Mongolia Agricultural University, 2019.] LI X C, Bioinformatics analysis and prokaryotic expression of mentaiioproteinnase gene from Pucciniahelianthi schw[D]. Hohhot: Inne Mongolia Agricultural University, 2019.
[39] MAJUMDER R, BANIK S P, KHOWALA S. Purification and characterisation of κ-casein specific milk-clotting metalloprotease from Termitomyces clypeatus MTCC 5091[J]. Food Chemistry,2015,173:441−448. doi: 10.1016/j.foodchem.2014.10.027
[40] LI Y, LIANG S, ZHI D J, et al. Purification and characterization of Bacillus subtilis milk-clotting enzyme from Tibet Plateau and its potential use in yak dairy industry[J]. European Food Research and Technology,2012,234(4):733−741. doi: 10.1007/s00217-012-1663-5
[41] NAVEED M, NADEEM F, MEHMOOD T, et al. Protease—a versatile and ecofriendly biocatalyst with multi-industrial applications:An updated review[J]. Catalysis Letters,2021,151(2):307−323. doi: 10.1007/s10562-020-03316-7
[42] MADHUSANKHA G, THILAKARATHNA R C N. Meat tenderization mechanism and the impact of plant exogenous proteases:A review[J]. Arabian Journal of Chemistry,2021,14(2):102967. doi: 10.1016/j.arabjc.2020.102967
[43] WILKESMAN J. Cysteine protease zymography:Brief review[J]. Zymography:Methods and Protocols, 2017:25−31.
[44] 薛意斌, 李雪, 李延啸, 等. 日本曲霉酸性蛋白酶的分泌表达、性质及其在猪肉嫩化中的应用[J]. 食品科学技术学报,2022,40(5):82−92. [XUE Y B, LI X, LI Y X, et al. Secretory expression and properties of acid protease from Aspergillus japonicus and its application in pork tenderization[J]. Journal of Food Science and Technology,2022,40(5):82−92.] doi: 10.12301/spxb202100910 XUE Y B, LI X, LI Y X, et al. Secretory expression and properties of acid protease from Aspergillus japonicus and its application in pork tenderization[J]. Journal of Food Science and Technology, 2022, 40(5): 82−92. doi: 10.12301/spxb202100910
[45] DENG L, WANG Z, YANG S, et al. Improvement of functional properties of wheat gluten using acid protease from Aspergillus usamii[J]. PLOS ONE,2016,11(7):101−101.
[46] 张依洁, 汪兰, 吴文锦, 等. 鸡骨泥风味饼干的研制[J]. 农产品加工,2014(21):10−12. [ZHANG Y J, WANG L, WU W J. et al. The preparation of mashed chicken-bone flavor biscuit[J]. Academic Periodical of Farm Products,2014(21):10−12.] ZHANG Y J, WANG L, WU W J. et al. The preparation of mashed chicken-bone flavor biscuit[J]. Academic Periodical of Farm Products, 2014(21): 10−12.
[47] RENZETTI S, ARENDT E. Effect of protease treatment on the baking quality of brown rice bread:From textural and rheological properties to biochemistry and microstructure[J]. Journal of Cereal Science,2009,50(1):22−28. doi: 10.1016/j.jcs.2009.02.002
[48] HATTA E, MATSUMOTO K, HONDA Y. Bacillolysin, papain, and subtilisin improve the quality of gluten-free rice bread[J]. Journal of Cereal Science,2015,61:41−47. doi: 10.1016/j.jcs.2014.10.004
[49] SONG P, CHENG L, TIAN K, et al. Biochemical characterization of two new Aspergillus niger aspartic proteases[J]. 3 Biotech,2020,10(7):1−9.
[50] LIU Y, SUN G, LI J, et al. Starter molds and multi-enzyme catalysis in koji fermentation of soy sauce brewing:A review[J]. Food Research International, 2024:114273.
[51] CHEN C, HOU S, WU C, et al. Improving protein utilization and fermentation quality of soy sauce by adding protease[J]. Journal of Food Composition and Analysis,2023,121:105399. doi: 10.1016/j.jfca.2023.105399
[52] FACIN B R, MELCHIORS M S, VALÉRIO A, et al. Driving immobilized lipases as biocatalysts:10 years state of the art and future prospects[J]. Industrial & Engineering Chemistry Research,2019,58(14):5358−5378.
[53] MATSUMOTO T, YAMADA R, OGINO H. Chemical treatments for modification and immobilization to improve the solvent-stability of lipase[J]. World Journal of Microbiology and Biotechnology,2019,35:1−8. doi: 10.1007/s11274-018-2566-9
[54] CASTILLA A, GIORDANO S R, IRAZOQUI G. Extremophilic lipases and esterases:Characteristics and industrial applications[J]. Microbial Extremozymes, 2022:207−222.
[55] YAO W, LIU K, LIU H, et al. A valuable product of microbial cell factories:Microbial lipase[J]. Frontiers in Microbiology,2021,12:743377. doi: 10.3389/fmicb.2021.743377
[56] SALGADO C A, DOS SANTOS C I A, VANETTI M C D. Microbial lipases:Propitious biocatalysts for the food industry[J]. Food Bioscience,2022,45:101509. doi: 10.1016/j.fbio.2021.101509
[57] ADETUNJI A I, OLANIRAN A O. Production strategies and biotechnological relevance of microbial lipases:A review[J]. Brazilian Journal of Microbiology,2021,52(3):1257−1269. doi: 10.1007/s42770-021-00503-5
[58] 史程风, 贾冉冉, 阎振丽, 等. 一株产低温脂肪酶酵母菌的鉴定及酶学性质[J]. 化工进展,2022,41(10):5541−5548. [SHI C F, JIA R R, YAN Z L, et al. Identification of a cold-adapted lipase-producing yeast and its enzyme characterization[J]. Chemical Industry and Engineering Progress,2022,41(10):5541−5548.] SHI C F, JIA R R, YAN Z L, et al. Identification of a cold-adapted lipase-producing yeast and its enzyme characterization[J]. Chemical Industry and Engineering Progress, 2022, 41(10): 5541−5548.
[59] ALI S, KHAN S A, HAMAYUN M, et al. The recent advances in the utility of microbial lipases:A review[J]. Microorganisms,2023,11(2):510−513. doi: 10.3390/microorganisms11020510
[60] VIVEK K, SANDHIA G, SUBRAMANIYAN S. Extremophilic lipases for industrial applications:A general review[J]. Biotechnology Advances,2022,60:108002. doi: 10.1016/j.biotechadv.2022.108002
[61] ZHAO J X, MA M M, YAN X H, et al. Characterization of a novel lipase from Bacillus licheniformis NCU CS-5 for applications in detergent industry and biodegradation of 2, 4-D butyl ester[J]. International Journal of Biological Macromolecules,2021,176(1):126−136.
[62] LE L T H L, YOO W, JEON S, et al. Biodiesel and flavor compound production using a novel promiscuous cold-adapted SGNH-type lipase (Ha SGNH1) from the psychrophilic bacterium Halocynthiibacter arcticus[J]. Biotechnology for Biofuels,2020,13(1):1−13. doi: 10.1186/s13068-019-1642-1
[63] GAO J X, OU X Y, XU P, et al. Cloning, overexpression, and characterization of a novel organic solvent-tolerant lipase from Paenibacillus pasadenensis CS0611[J]. Chinese Journal of Catalysis,2018,39(5):937−945. doi: 10.1016/S1872-2067(18)63033-5
[64] ÖZDEMIR F İ, TÜLEK A, ERDOĞAN D. Identification and heterologous production of a lipase from Geobacillus kaustophilus DSM 7263T and tailoring its N-terminal by a His-tag epitope[J]. The Protein Journal,2021,40(3):436−437. doi: 10.1007/s10930-021-09987-4
[65] TAKÓ M, KOTOGÁN A, PAPP T, et al. Purification and properties of extracellular lipases with transesterification activity and 1, 3-regioselectivity from Rhizomucor miehei and Rhizopus oryzae[J]. Journal of Microbiology and Biotechnology,2017,27(2):277−288. doi: 10.4014/jmb.1608.08005
[66] DUA A, GUPTA R. Functional characterization of hormone sensitive-like lipase from Bacillus halodurans:Synthesis and recovery of p NP-laurate with high yields[J]. Extremophiles,2017,21(5):871−889. doi: 10.1007/s00792-017-0949-8
[67] 韩生义, 赵淑琴, 刘晓丽, 等. 一株碱性脂肪酶产生菌的筛选, 鉴定及酶学性质研究[J]. 甘肃农业大学学报,2017,52(1):119−125. [HAN S Y, ZHAO S Q, LIU X L, et al. Screening, identification and enzymatic properties of an alkaline lipase producing strain[J]. Journal of Gansu Agricultural University,2017,52(1):119−125.] HAN S Y, ZHAO S Q, LIU X L, et al. Screening, identification and enzymatic properties of an alkaline lipase producing strain[J]. Journal of Gansu Agricultural University, 2017, 52(1): 119−125.
[68] DE ALMEIDA A F, TERRASAN C R F, TERRONE C C, et al. Biochemical properties of free and immobilized Candida viswanathii lipase on octyl-agarose support:Hydrolysis of triacylglycerol and soy lecithin[J]. Process Biochemistry,2018,65:71−80. doi: 10.1016/j.procbio.2017.10.019
[69] GAO K P, WANG X F, JIANG H, et al. Identification of a GDSL lipase from Streptomyces bacillaris and its application in the preparation of free astaxanthin[J]. Journal of Biotechnology,2021,325:280−287. doi: 10.1016/j.jbiotec.2020.10.009
[70] 李晓楠. 产脂肪酶乳酸菌的筛选及其对奶酪风味的影响[D]. 石河子:石河子大学, 2021. [LI X N. Screening of lipase-producing lactic acid bacteria and its effect on cheese flavor[D]. Shihezi:Shihezi University, 2021.] LI X N. Screening of lipase-producing lactic acid bacteria and its effect on cheese flavor[D]. Shihezi: Shihezi University, 2021.
[71] SU H F, MAI Z M, ZHANG S. Cloning, expression and characterization of a lipase gene from marine bacterium Pseudoalteromonas lipolytica SCSIO 04301[J]. Journal of Ocean University of China,2016,15(6):1051−1058. doi: 10.1007/s11802-016-3071-0
[72] SU H F, MAI Z M, YANG J, et al. Cloning, expression, and characterization of a cold-active and organic solvent-tolerant lipase from Aeromicrobium sp. SCSIO 25071[J]. Journal of Microbiology and Biotechnology,2016,26(6):1067−1076. doi: 10.4014/jmb.1511.11068
[73] CHANDRA P, ENESPA, SINGH R, et al. Microbial lipases and their industrial applications:A comprehensive review[J]. Microbial Cell Factories,2020,19:1−42. doi: 10.1186/s12934-019-1269-8
[74] MUNIANDY M, LASEKAN O, GHAZALI H M, et al. Lipase-catalyzed formation of pentyl nonanoate using screened immobilized lipase from Rhizomucor meihei[J]. Brazilian Journal of Chemical Engineering,2019,36(3):1089−1097. doi: 10.1590/0104-6632.20190363s20180419
[75] 翟磊, 信春晖, 许玲, 等. 芝麻香型白酒高温大曲产脂肪酶菌株的筛选与鉴定[J]. 食品与发酵工业,2015,41(12):36−39. [ZHAI L, XIN C H, XU L. Screening and identification of lipase-producing strains from high-temperature Daqu in sesame flavor liquor[J]. Food and Fermentation Industries,2015,41(12):36−39.] ZHAI L, XIN C H, XU L. Screening and identification of lipase-producing strains from high-temperature Daqu in sesame flavor liquor[J]. Food and Fermentation Industries, 2015, 41(12): 36−39.
[76] 李站胜, 颜晨麟, 江宁, 等. 脂肪酶处理白酒丢糟提升复糟酒的品质[J]. 现代食品科技,2019,25(5):191−197. [LI Z S, YAN C L, JIANG N, et al. Lipase treatment of distiller's grains wine to promote its quality[J]. Modern Food Science and Technology,2019,25(5):191−197.] LI Z S, YAN C L, JIANG N, et al. Lipase treatment of distiller's grains wine to promote its quality[J]. Modern Food Science and Technology, 2019, 25(5): 191−197.
[77] 蔡琪琪. 红曲霉在红曲黄酒酿造过程中的作用研究[D]. 福州:福州大学, 2015. [CAI Q Q. Effect of Monascus on the fermentation of Hong Qu glutinous rice wine[D]. Fuzhou:Fuzhou University, 2015.] CAI Q Q. Effect of Monascus on the fermentation of Hong Qu glutinous rice wine[D]. Fuzhou: Fuzhou University, 2015.
[78] COLAKOGLU A S, ÖZKAYA H. Potential use of exogenous lipases for DATEM replacement to modify the rheological and thermal properties of wheat flour dough[J]. Journal of Cereal Science,2012,55(3):397−404. doi: 10.1016/j.jcs.2012.02.001
[79] 钱忠英, 刘滔, 杨环毓, 等. 重组疏棉状嗜热丝孢菌脂肪酶对面包烘焙品质的影响[J]. 食品工业科技,2020,41(9):74−80. [QIAN Z Y, LIU T, YANG H Y. Effect of recombinant Thermomyces lanuginosus lipase on bread baking quality[J]. Science and Technology of Food Industry,2020,41(9):74−80.] QIAN Z Y, LIU T, YANG H Y. Effect of recombinant Thermomyces lanuginosus lipase on bread baking quality[J]. Science and Technology of Food Industry, 2020, 41(9): 74−80.
[80] HUANG Z G, BRENNAN C S, ZHENG H T, et al. The effects of fungal lipase-treated milk lipids on bread making[J]. LWT,2020,128:109455. doi: 10.1016/j.lwt.2020.109455
[81] ZHANG X M, AI N S, WANG J, et al. Lipase-catalyzed modification of the flavor profiles in recombined skim milk products by enriching the volatile components[J]. Journal of Dairy Science,2016,99(11):8665−8679. doi: 10.3168/jds.2015-10773
[82] OMAR K A, GOUNGA M E, LIU R, et al. Effects of microbial lipases on hydrolyzed milk fat at different time intervals in flavour development and oxidative stability[J]. Journal of Food Science & Technology,2016,53(2):1035−1046.
[83] NAVVABI A, RAZZAGHI M, FERNANDES P, et al. Novel lipases discovery specifically from marine organisms for industrial production and practical applications[J]. Process Biochemistry,2018,70:61−70. doi: 10.1016/j.procbio.2018.04.018
[84] 方洪枫, 张慧霞, 王国红, 等. 混菌发酵绿茶产脂肪酶及其对酯型儿茶素的水解作用分析[J]. 茶叶科学,2019,39(1):88−97. [FANG H F, ZHANG H X, WANG G H, et al. Fungal mixed fermentation for the production of lipase and its activity analysis in galloylated catechin hydrolysis[J]. Journal of Tea Science,2019,39(1):88−97.] doi: 10.3969/j.issn.1000-369X.2019.01.010 FANG H F, ZHANG H X, WANG G H, et al. Fungal mixed fermentation for the production of lipase and its activity analysis in galloylated catechin hydrolysis[J]. Journal of Tea Science, 2019, 39(1): 88−97. doi: 10.3969/j.issn.1000-369X.2019.01.010
[85] 马超龙, 李小嫄, 岳翠男, 等. 茶叶中脂肪酸及其对香气的影响研究进展[J]. 食品研究与开发,2017,38(4):220−224. [MA C L, LI X Y, YUE C N, et al. Progress on fatty acids in tea and their influences on tea aroma[J]. Food Research and Development,2017,38(4):220−224.] doi: 10.3969/j.issn.1005-6521.2017.04.052 MA C L, LI X Y, YUE C N, et al. Progress on fatty acids in tea and their influences on tea aroma[J]. Food Research and Development, 2017, 38(4): 220−224. doi: 10.3969/j.issn.1005-6521.2017.04.052
[86] 孙浩, 倪辉, 陈峰. 黑曲霉粗酶液对绿茶产品风味及挥发性成分的影响[C]. 中国食品科学技术学会第十三届年会论文摘要集. 北京:中国食品科学技术学会, 2016:2. [SUN H, NI H, CHEN F. Effects of two enzyme extract of Aspergillus niger on odour characteristic and volatile compounds of green tea[C]. The 13th Annual Meeting of CIFST. Beijing:Chinese Institute of Food Science and Technology, 2016:2.] SUN H, NI H, CHEN F. Effects of two enzyme extract of Aspergillus niger on odour characteristic and volatile compounds of green tea[C]. The 13th Annual Meeting of CIFST. Beijing: Chinese Institute of Food Science and Technology, 2016: 2.
 下载:
下载:


计量
- 文章访问数: 28
- HTML全文浏览量: 32
- PDF下载量: 4



