


Extraction Process Optimization and Bioactivity Analysis of Andrias davidianus Liver Metallothionein
-
摘要: 为提高大鲵加工副产物利用价值,探索两栖动物金属硫蛋白结构、活性和功能,本研究采用匀浆离心法从大鲵肝脏提取金属硫蛋白(AdMT),通过单因素和正交试验优化提取工艺,利用离子交换层析分离纯化,以自由基清除能力检测抗氧化活性,乳酸脱氢酶释放率测试细胞毒性,脂多糖诱导RAW264.7细胞炎症模型评价抗炎活性,CCK-8法分析细胞增殖能力和免疫原性,流式细胞术检测细胞周期,划痕实验测试细胞迁移能力,CdCl2诱导L929和HaCaT细胞中毒模型评价重金属解毒能力。结果表明:最佳提取工艺为液料比5:1、提取液浓度0.02 mol/L、提取温度40 ℃,提取时间1 h,提取含量为0.362 mg/g。AdMT对ABTS+自由基的清除率达到95.01%,对L929和HaCaT细胞乳酸脱氢酶释放率及RAW264.7细胞存活率均无显著影响。AdMT处理48 h后,L929和HaCaT细胞存活率分别达到192.63%和207.92%,细胞增殖指数分别上升至0.56和0.53。同时,细胞迁移能力随着AdMT浓度增加不断增强,划痕愈合率分别为63.38%和76.23%。此外,AdMT能显著(P<0.05,P<0.01,P<0.001)降低细胞炎症模型中炎症因子的表达水平,将镉中毒细胞存活率恢复至对照组水平。研究结果为大鲵金属硫蛋白的进一步开发和大鲵副产物加工利用提供了参考。Abstract: In this study, the by-products processed from Andrias davidianus were evaluated, and the structure, activity, and functionality of the metallothionein from this amphibian species (AdMT) were investigated. AdMT was extracted from the liver using a centrifugal homogenization method, with the extraction process optimized by means of single-factor and orthogonal experiments. Subsequently, the protein was purified using ion-exchange chromatography. The antioxidative activity of AdMT was assessed by measuring its free radical-scavenging ability, and its cytotoxicity was evaluated using the lactate dehydrogenase release assay. The anti-inflammatory activity of the protein was determined by RAW264.7 cell model of lipopolysaccharide-induced inflammation, and its immunogenicity and effect on cell proliferation were analyzed using the CCK-8 assay. Its effects on the cell cycle and cell migration ability were assessed using flow cytometric and scratch assays, respectively. Its heavy metal detoxification capacity was evaluated using L929 and HaCaT cell models of CdCl2-induced toxicity. Results showed that, the optimal AdMT extraction conditions were determined to be as follows: Liquid-to-material ratio of 5:1, extraction solution concentration of 0.02 mol/L, extraction temperature of 40 ℃, and extraction time of 1 h, yielding 0.362 mg/g of the protein. AdMT exhibited a 95.01% scavenging rate against ABTS+ free radicals and had no significant impact on the lactate dehydrogenase release rates of L929 and HaCaT cells or on the viability of RAW264.7 cells. After 48 h of AdMT treatment, the viability of L929 and HaCaT cells improved to 192.63% and 207.92%, respectively, and their cell proliferation indices increased to 0.56 and 0.53, respectively. Additionally, the migration ability of the L929 and HaCaT cells increased with increasing AdMT concentration, showing scratch healing rates were 63.38% and 76.23%, respectively. In the cell inflammation model, AdMT significantly (P<0.05, P<0.01, P<0.001) reduced the expression levels of inflammatory factors. Furthermore, the survival rate of Cd-exposed cells was restored to that of the control group. These results would provide a reference for the development of AdMT and utilization of by-products processed from Andrias davidianus.
-
大鲵(Andrias davidianus)是现存寿命最长、体型最大的两栖动物,具有悠久的食药两用历史,各部位中共含有70余种生物活性成分[1],是一种高利用价值的生物资源[2]。随着人工繁育技术的突破,人工养殖子二代大鲵已经成为我国陕西、贵州、重庆、湖南、江西等地区的特色产业[3],但目前的销售仍局限于餐桌消费,产业发展受到严重约束。大鲵体长0.5~1.5 m,重2~20 kg,其肝脏约占总体质量的3%~5%,但由于腥味重,在加工中常作为副产物被直接丢弃[4],若能充分研究和利用这一废弃资源,加大综合开发力度,精深加工高附加值产品,对保护环境、提升资源利用率和提高大鲵产业经济效益有着重要意义。
金属硫蛋白(Metallothionein,MT)是一种富含半胱氨酸,参与机体金属稳态和氧化应激调节的低分子量蛋白质家族,具有重金属解毒、神经系统保护、调节生长和发育、抗癌、抗氧化、参与炎症和免疫应答等多种活性和功能[5−7],其发展和进步越来越受到研究者的关注。MT广泛存在于生物体中,已有文献报道从鼠、猪等陆生动物[8−9],口虾蛄、牡蛎、扇贝等水生动物[10−12]中提取分离MT。研究发现,MT在长寿的物种中表达量更高[13]。大鲵作为现存寿命最长的两栖动物,其肝脏中富含MT。然而,从两栖动物组织中提取MT的研究还较少,其结构、活性和功能等方面仍不清楚,对大鲵肝脏金属硫蛋白的提取和活性研究是一种有利于产业发展的生物资源加工利用方式。
本研究以大鲵肝脏为原料,采用匀浆离心法提取MT,通过正交试验优化提取工艺,利用离子交换层析分离纯化,检测其抗氧化、抗炎、促进细胞增殖和迁移活性及重金属解毒能力。研究结果有利于了解两栖动物MT的特点和生物活性,为其生物医药开发提供实验基础,同时也为大鲵副产物加工利用提供参考。
1. 材料与方法
1.1 材料与仪器
大鲵肝脏 由贵州黔梦农业科技有限公司提供;金属硫蛋白标准品 源于兔肝脏,纯度>98%,上海颖心实验室设备有限公司;三羟甲基氨基甲烷(Tris)、苯甲基磺酰氟(PMSF)、二硫苏糖醇(DTT)、DEAE-琼脂糖凝胶FF、葡聚糖凝胶G-15、考马斯亮蓝R-250、ABTS(2,2-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐,纯度≥98%)、DPPH(1,1-二苯基-2-三硝基苯肼,纯度>97%)、CCK-8试剂盒、彩虹245plus广谱蛋白Marker(5~245 kD) 北京索莱宝科技有限公司;过硫酸钾(>99.5%)、5,5'二硫代双(2-硝基苯甲酸)(98%) 上海麦克林生化科技有限公司;HaCaT细胞株、RAW264.7细胞株、L929细胞株、十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)凝胶制备试剂盒 武汉博士德生物工程有限公司;MEM培养基、DMEM培养基、RPMI 1640培养基 美国Gibco公司;RNA提取试剂盒、RNA反转录试剂盒、qPCR Mix 南京诺唯赞生物科技股份有限公司;0.25%胰蛋白酶(含EDTA)、乳酸脱氢酶细胞毒性检测试剂盒 上海碧云天生物科技有限公司;LPS(脂多糖) 美国Sigma公司;引物合成由生工生物工程(上海)股份有限公司完成;其他试剂均为分析纯。
JJ-2组织捣碎匀浆机 常州国华电器有限公司;HWCC-3集热式恒温磁力搅拌器 郑州长城科工贸有限公司;Allegra V-15R高速冷冻离心机 美国BECKMAN COULTER公司;T100 PCR仪、CHemiDoc-XRS+凝胶成像系统 美国Bio-Rad公司;DYY-6C电泳仪 北京六一生物科技有限公司;Triacl 2.5 L真空冷冻干燥机 美国Labconco公司;311 CO2细胞培养箱、Evolution 200紫外可见分光光度计 美国Thermo Fisher公司;Spectra Max M2多功能酶标仪 美国Molecular Devices公司;NovoCyte 2040R流式细胞仪 美国艾森生物科学公司。
1.2 实验方法
1.2.1 大鲵金属硫蛋白提取
参照文献[12]描述的方法稍作修改。取50 g大鲵肝脏解冻后加一定量的Tris-HCl提取液(含1 mmol/L DTT防止氧化,pH8.6),组织匀浆,恒温水浴振荡提取。4 ℃下离心(11800×g)20 min,收集上清,80 ℃加热5 min去除热不稳定蛋白,冰浴冷却,离心取上清,加入3倍体积−20 ℃预冷的无水乙醇,4 ℃下过夜沉淀。离心收集沉淀,加10 mL Tris-HCl室温溶解5 h,离心收集上清即为大鲵金属硫蛋白(AdMT)粗提液。在−40 ℃冷冻干燥保存备用。
1.2.2 金属硫蛋白提取含量测定
1.2.2.1 标准溶液配制
参照吴云辉等[14]的方法,先配制质量浓度为 1 μg/μL的MT标准品溶液,分别取不同体积的MT标准溶液加入4 mL离心管中,依次加入10 μL HCl(1.2 mol/L)和200 μL EDTA(0.1 mol/L),于暗处反应10 min脱去金属;再加入5 mmol/L DTNB充分涡旋混匀3 min,使MT与DTNB形成黄色络合物,最后用Tris-HCl缓冲液(pH7.5)稀释至4 mL,在412 nm波长下通过比色法测定显色产物硫代硝基苯甲酸阴离子(TNBA)的量来确定MT含量。
1.2.2.2 标准曲线绘制
根据不同含量MT反应后所测得的吸光度值不同,以MT标准品浓度为横坐标,吸光度值为纵坐标绘制标准曲线。得到线性回归方程为y=0.004878x+0.0580(R2=0.9994)。
1.2.2.3 样品测定
取1.2.1中收集的AdMT粗提液1 mL,按1.2.2.1方法测定吸光度,根据标准曲线确定提取含量。
1.2.3 单因素实验
按照1.2.1提取方法,称取50 g大鲵肝脏,在Tris-HCl提取液体积与肝脏质量液料比3:1 mL:g、Tris-HCl提取液浓度0.02 mol/L、提取温度40 ℃、提取时间1.5 h条件下,研究液料比(2:1、3:1、4:1、5:1、6:1、7:1 mL:g)、提取液浓度(0.01、0.02、0.05、0.10、0.15、0.20 mol/L)、提取温度(20、30、40、50、60、70 ℃)及提取时间(0.5、1.0、1.5、2.0、2.5、3.0 h)对AdMT提取含量的影响。
1.2.4 正交试验
根据单因素实验结果,确定因素最优范围,以AdMT提取含量为指标,采用L9(34)正交试验对液料比(A)、提取液浓度(B)、提取温度(C)和提取时间(D)四个因素进行优化,具体因素和水平设计见表1。
表 1 正交试验因素与水平Table 1. Factors and levels of orthogonal tests水平 A液料比
(mL:g)B提取液浓度
(mol/L)C提取温度
(℃)D提取时间
(h)1 3:1 0.01 30 1.0 2 4:1 0.02 40 1.5 3 5:1 0.05 50 2.0 1.2.5 大鲵金属硫蛋白鉴定
1.2.5.1 SDS-PAGE
参考何蜀峰等[15]的方法略作修改。配制15%的分离胶和4%的浓缩胶进行SDS-PAGE电泳,电泳条件如下:样品浓度为3 mg/mL,上样体积10 μL,浓缩胶电压80 V、时间20 min,分离胶电压120 V、约1.5 h待溴酚蓝指示带到达分离胶底部时结束电泳。电泳后用考马斯亮蓝R-250染色30 min,然后脱色液脱色至背景清晰,使用凝胶成像系统采集成像。
1.2.5.2 红外光谱分析
取一定量AdMT样品,与溴化钾混匀压片,采用傅里叶红外光谱仪测定,分辨率4 cm−1,扫描32次,扫描范围为4000~400 cm−1。
1.2.6 金属硫蛋白分离纯化
1.2.6.1 阴离子交换层析
参考李停停[16]的方法并作改进。选用离子交换填料DEAE-琼脂糖凝胶FF填装层析柱(2.6×30 cm)对AdMT粗提物进行分离纯化。将样品溶于0.02 mol/L Tris-HCl中,过0.45 μm滤膜后上样,使用0.02、0.2、0.3、0.4、0.5 mol/L NaCl溶液,以0.02 mol/L Tris为缓冲成分,梯度洗脱,流速为2.0 mL/min,收集254 nm滤光片下有明显吸收峰的组分,采用SDS-PAGE分析各组分蛋白分子量,根据组分条带单一性和分子量范围(6~7 kDa)确定目标峰组分。
1.2.6.2 脱盐
将选择的目标峰组分进行Sephadex G-15(2.6×40 cm)脱盐,使用超纯水进行洗脱,流速为1 mL/min,设置自动收集3 mL/管,254 nm滤光片下将收集到的目的组分冷冻干燥,−40 ℃保存。
1.2.7 抗氧化活性
1.2.7.1 ABTS+自由基清除能力
根据文献[17]描述的方法略作修改。等体积7.4 mmol/L ABTS溶液和2.6 mmol/L K2S2O8溶液混匀,黑暗条件下室温反应14~16 h形成ABTS+自由基储备溶液,用无水乙醇稀释到在734 nm处吸光度为0.70±0.01,将AdMT溶液与ABTS+自由基液1:19混匀室温避光反应30 min后于734 nm处测定吸光度。按照公式(1)计算ABTS+自由基清除率。
(1) 式中:A为AdMT组的吸光度;A0为去离子水代替AdMT溶液的吸光度。
1.2.7.2 DPPH自由基清除能力
DPPH自由基清除能力测试按照文献[18]方法进行。将一定体积的0.1 mmol/L DPPH溶液与等体积的AdMT溶液混合均匀,于室温下避光反应20 min。反应后,于517 nm下测定吸光度。DPPH自由基的清除率根据公式(2)计算。
(2) 式中,A为AdMT组的吸光度值;A0为相同条件下去离子水代替AdMT的吸光度值;B为95%乙醇溶液代替DPPH的吸光度值。
1.2.8 细胞培养
L929、HaCaT和RAW264.7细胞株用完全培养基(含10%胎牛血清)于37 ℃、5% CO2培养箱进行孵育培养,期间每1~2 d更换培养基,待细胞密度生长至约80%~90%时传代,L929和HaCaT用无菌PBS清洗后使用含EDTA的胰蛋白酶消化液消化收集细胞,RAW264.7使用细胞刮刀收集细胞,1000 r/min离心4 min后弃去上清,加入新鲜完全培养基重悬细胞,按照后续实验要求的细胞数量传代或接种至细胞培养板。
1.2.9 细胞毒性及免疫原性
1.2.9.1 细胞毒性
通过乳酸脱氢酶(LDH)释放率检测细胞膜损伤,测试AdMT的细胞毒性[19]。将L929和HaCaT细胞以1×104个/孔接种于96孔板,待细胞贴壁后加入AdMT使其至不同终浓度(0、25、100、250、500、1000 μg/mL),24 h后进行测定,根据乳酸脱氢酶细胞毒性检测试剂盒步骤进行实验,在490 nm处测定样品吸光度值,LDH释放率根据公式(3)计算。
(3) 式中,A为AdMT组的吸光度值;A0为对照组的吸光度值;A1为细胞最大酶活性对照孔的吸光度值,即加入LDH释放剂的分组。
1.2.9.2 免疫原性
选择RAW264.7细胞,采用CCK-8法检测AdMT的免疫原性。将RAW264.7细胞以5×104个/孔接种于96孔板,贴壁后加入AdMT至不同终浓度(0、25、100、250、500、1000 μg/mL)共培养12/24 h,加入1/10体积的CCK-8试剂,在37 ℃避光孵育1~2 h,于450 nm波长下检测吸光度,细胞存活率根据公式(4)计算。
(4) 式中,As为AdMT组的吸光度值;Ab为空白组的吸光度值,即只加入培养基和CCK-8试剂;Ac为对照组的吸光度值。
1.2.10 细胞增殖能力
1.2.10.1 细胞活力测定
采用CCK-8法,测定AdMT对L929和HaCaT细胞活力的影响。细胞以1×104个/孔接种于96孔板,待细胞贴壁后加入AdMT使其至不同终浓度(0、25、100、250、500、1000 μg/mL),24/48 h后参照1.2.9.2测定细胞活力。
1.2.10.2 细胞周期
取对数生长期的L929和HaCaT细胞以5×105个/孔接种于6孔板,培养至贴壁后,加入AdMT使其至不同终浓度(0、25、100、250、500、1000 μg/mL),24 h后收集细胞并用冰浴预冷的PBS洗涤,于4 ℃下在70%乙醇中固定12 h,离心去除上清,加入0.5 mL碘化丙啶染色液,缓慢并充分重悬细胞沉淀,37 ℃避光温浴30 min,使用流式细胞仪进行检测。分析处于G0/G1期、S期、G2/M期各阶段细胞占比,细胞增殖指数(PI)根据公式(5)计算。
(5) 式中:A1为G0/G1期细胞;A2为S期细胞;A3为G2/M期细胞。
1.2.11 细胞迁移能力
取对数生长期的L929和HaCaT细胞铺于6孔板,以第2 d正好长满为宜,用20 μL枪头制造划痕,加入AdMT(终浓度0、25、100、250、500 μg/mL)的含1%血清培养基继续培养24 h,拍照观察,利用Image J计算划痕面积,根据公式(6)计算细胞迁移率。
(6) 式中:A0为起始划痕面积;A1为培养24 h后划痕面积。
1.2.12 抗炎活性
参照朱彦彬等[20]的方法,用100 ng/μL LPS建立RAW264.7细胞炎症模型,3 h后加入AdMT使终浓度为500 μg/mL,培养12 h,收集细胞。采用Trizol法提取总RNA,逆转录为cDNA,以GAPDH为内参,RT-qPCR检测炎症因子IL-1β、IL-6、TNF-α的表达,其中引物序列见表2。
表 2 RT-qPCR引物Table 2. Primers sequences used for RT-qPCR引物名称 引物序列 IL-1β F:CCAGGATGAGGACCCAAGCA;
R:TCCCGACCATTGCTGTTTCCIL-6 F:GAGTCCTTCAGAGAGATACAG;
R:CTGTGACTCCAGCTTATCTGTNF-α F:GTCTCAGCCTCTTCTCATTC;
R:CATAGAACTGATGAGAGGGAGAPDH F:GTGGCAAAGTGGAGATTGTTG;
R:CTCCTGGAAGATGGTGATGG1.2.13 重金属解毒能力
1.2.13.1 重金属中毒模型构建
参照文献[21]方法,采用CdCl2构建L929和HaCaT细胞重金属中毒模型。将L929和HaCaT细胞以1×104个/孔细胞接种于96孔板,培养至细胞贴壁。用培养基配制不同浓度(0、10、15、20、25、50、100 μmol/L)的CdCl2溶液,加入细胞中共培养24 h。以CCK-8法检测细胞存活率,选择存活率为65%~75%时的CdCl2浓度为模型诱导剂量。
1.2.13.2 AdMT的重金属解毒能力
加入一定浓度的CdCl2构建重金属中毒模型,3 h后,加入AdMT使其为不同的终浓度(0、25、100、250、500 μg/mL),共培养24 h后,以CCK-8法检测细胞存活率。
1.3 数据处理
本文中所涉及实验至少平行重复3次,数据以平均值±标准差(SD)表示。使用SPSS 27.0单因素方差分析(one-way ANOVA)中的Tukey's HSD检验确认是否存在显著统计学差异,P<0.05表示存在显著差异,ns表示P>0.05,*表示P<0.05,**或##表示P<0.01,***或###表示P<0.001,不同字母表示各组间存在显著差异。
2. 结果与分析
2.1 单因素实验
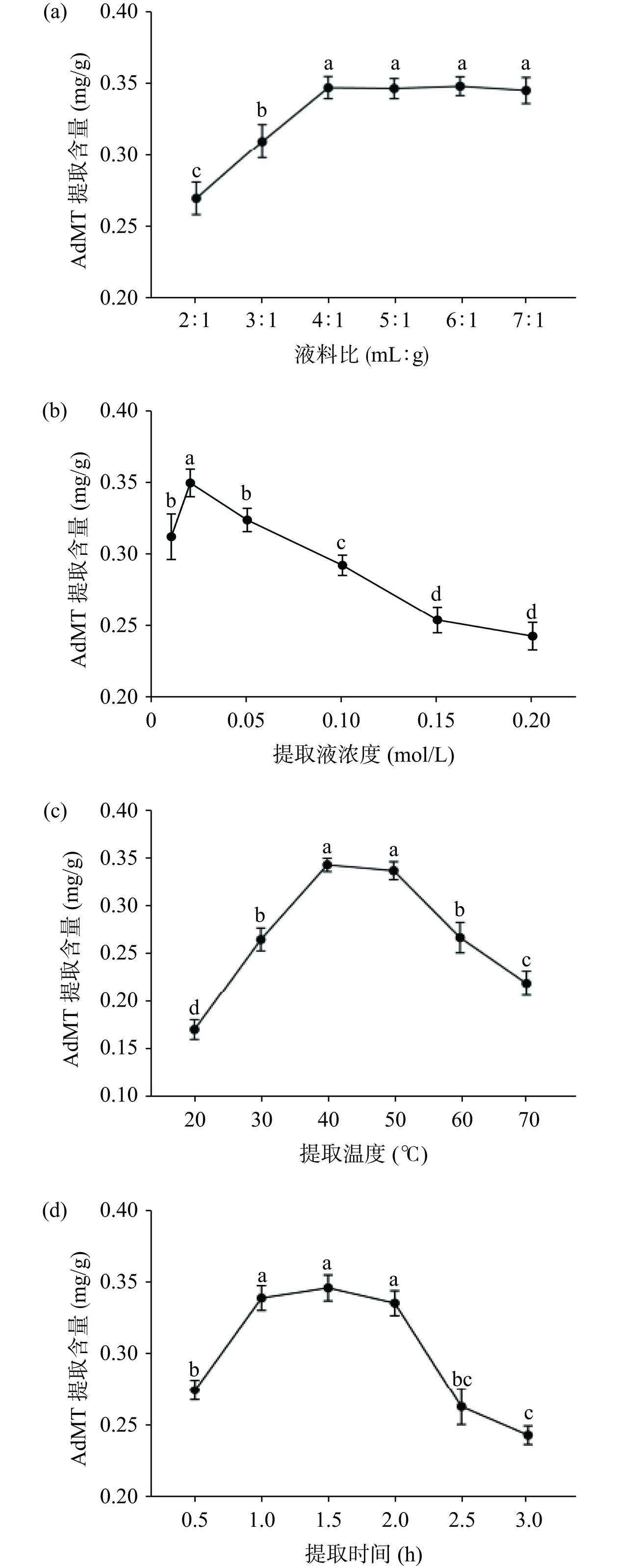
不同因素对AdMT提取含量的影响存在差异(图1)。随着液料比的增大,提取含量先增加后保持不变,对于提取液浓度、提取温度和提取时间,则先增加后降低。在一定范围内,增大液料比可以增加肝脏组织与提取液的接触率,提高AdMT溶解量。当液料比为4:1时,提取液已将肝脏中AdMT充分溶解,提取含量在此水平上稳定。提取液浓度通过离子强度影响AdMT溶解量,在0.02 mol/L时,提取含量达到最高。然而,过高的离子强度可能会导致蛋白发生沉淀[16]。提取温度影响溶剂分子扩散,随着温度的升高,AdMT分子扩散速率增大,提取率快速增加,在40 ℃时达到最大,但较高温度下的长时间提取过程可能会导致部分AdMT变性。提取时间对溶解过程产生影响,时间的延长使AdMT溶解更充分。然而,过长的提取时间(超过1.5 h)会使AdMT中巯基氧化[22],导致在利用DTNB比色法检测AdMT提取含量时,原有与DTNB反应生成5-巯基-2-硝基苯甲酸巯基减少,从而被检测到的AdMT提取含量降低。因此,各因素的单因素结果为:液料比4:1、提取液浓度0.02 mol/L、提取温度40 ℃、提取时间1.5 h。
![]()
2.2 正交试验
四个因素对AdMT提取含量的影响程度不同(表3),从极差R可知,影响程度按大小排序为A>C>D>B。液料比影响最显著,其次是提取温度和提取时间,提取液浓度影响最弱。AdMT的提取是一个溶解过程,提取液(溶剂)的量决定了能溶解的AdMT(溶质)的量。当肝脏组织的量一定时,其中含有的AdMT量不变,液料比就决定了最大能溶解的AdMT量,因此影响最显著。由于AdMT具有很好的溶解性,在不影响结构的前提下,提取液(Tris-HCl)中离子强度对溶解过程的影响并不明显。对于提取温度和时间,分别影响AdMT的溶解速率和效率,在提取过程中影响程度适中。根据最佳水平得到AdMT的最优提取工艺A3B2C2D1,即液料比5:1、提取液浓度0.02 mol/L、提取温度40 ℃,提取时间1 h。进一步的验证实验表明,在该条件下的提取含量为0.362 mg/g,高于正交试验中的各组数据,说明结果可靠。本研究从人工饲养的大鲵肝脏中提取AdMT,提取含量高于口虾蛄[10](0.232 mg/g)、牡蛎[11](0.221 mg/g)、扇贝[12](0.115 mg/g)等水生动物,且明显高于植物鹰嘴豆[23](0.16 mg/g),这表明大鲵肝脏富含金属硫蛋白。大鲵作为现存体型最大的两栖动物,肝脏占总质量的3%~5%,且常作为加工副产物未被充分利用。因此,大鲵肝脏是一种含量丰富、组织体积大且价格低廉的优质金属硫蛋白提取原料。
表 3 正交试验结果Table 3. Results of orthogonal tests实验号 液料比 提取液浓度 提取温度 提取时间 AdMT提取
含量(mg/g)1 1 1 1 1 0.334 2 1 2 3 2 0.344 3 1 3 2 3 0.339 4 2 1 3 3 0.348 5 2 2 2 1 0.357 6 2 3 1 2 0.341 7 3 1 2 2 0.351 8 3 2 3 3 0.346 9 3 3 1 1 0.352 K 1 1.02 1.03 1.02 1.04 2 1.05 1.05 1.05 1.04 3 1.05 1.03 1.04 1.03 k 1 0.34 0.34 0.34 0.35 2 0.35 0.35 0.35 0.35 3 0.35 0.34 0.35 0.34 最佳水平 3 2 2 1 R 0.54 0.02 0.25 0.11 2.3 AdMT鉴定
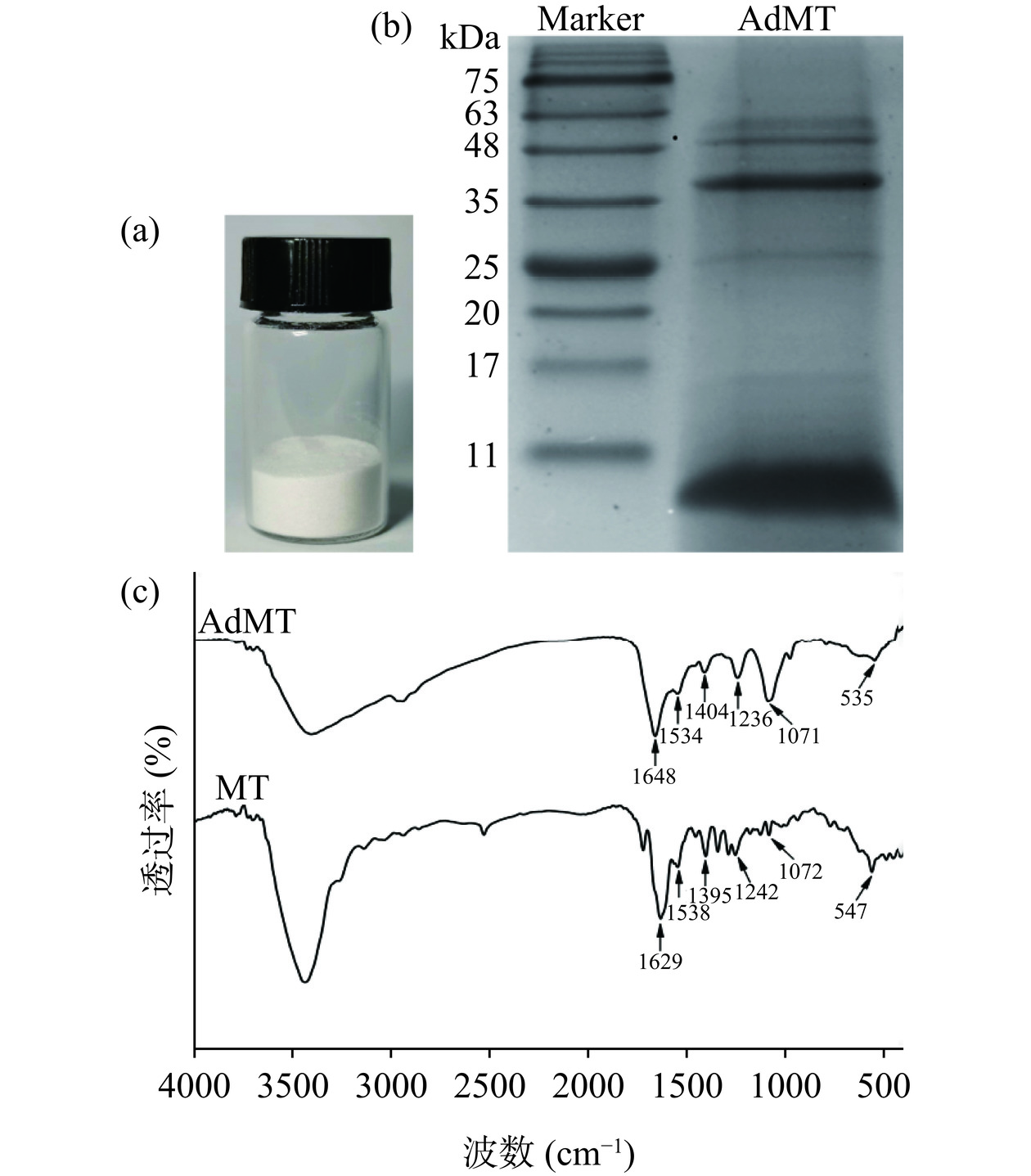
匀浆离心法提取的AdMT经冷冻干燥后,为白色粉末(图2a),易溶于水,与口虾蛄MT外观一致[24]。SDS-PAGE分析表明,AdMT粗提物样品中有4条明显条带(图2b),其中在6 kDa处条带最明显(含量最高),与MT理论分子量大小一致[25]。AdMT和兔肝MT标准品红外光谱如图2c,AdMT中位于1648 cm−1处吸收峰归属于酰胺Ⅰ带羰基的C=O伸缩振动(1700~1600 cm−1),1534 cm−1处吸收峰来源于酰胺Ⅱ带N-H弯曲振动和C-N伸缩振动(1600~1500 cm−1),1236 cm−1处吸收峰对应酰胺Ⅲ带(1330~1220 cm−1)。这些酰胺基团的特征吸收峰位置与标准品基本一致,且与文献报道MT特征峰相符[26]。这些结果表明,从大鲵肝脏中提取的粗提物具有分子量和特征基团与MT一致的目标蛋白,但由于存在杂蛋白,后续需要进一步纯化。
![]() 图 2 AdMT提取物(a)、15% SDS-PAGE图(b)及红外光谱扫描图(c)Figure 2. AdMT extract (a), 15% SDS-PAGE patterns (b) and infrared spectral scanning (c) of AdMT
图 2 AdMT提取物(a)、15% SDS-PAGE图(b)及红外光谱扫描图(c)Figure 2. AdMT extract (a), 15% SDS-PAGE patterns (b) and infrared spectral scanning (c) of AdMT2.4 AdMT分离纯化
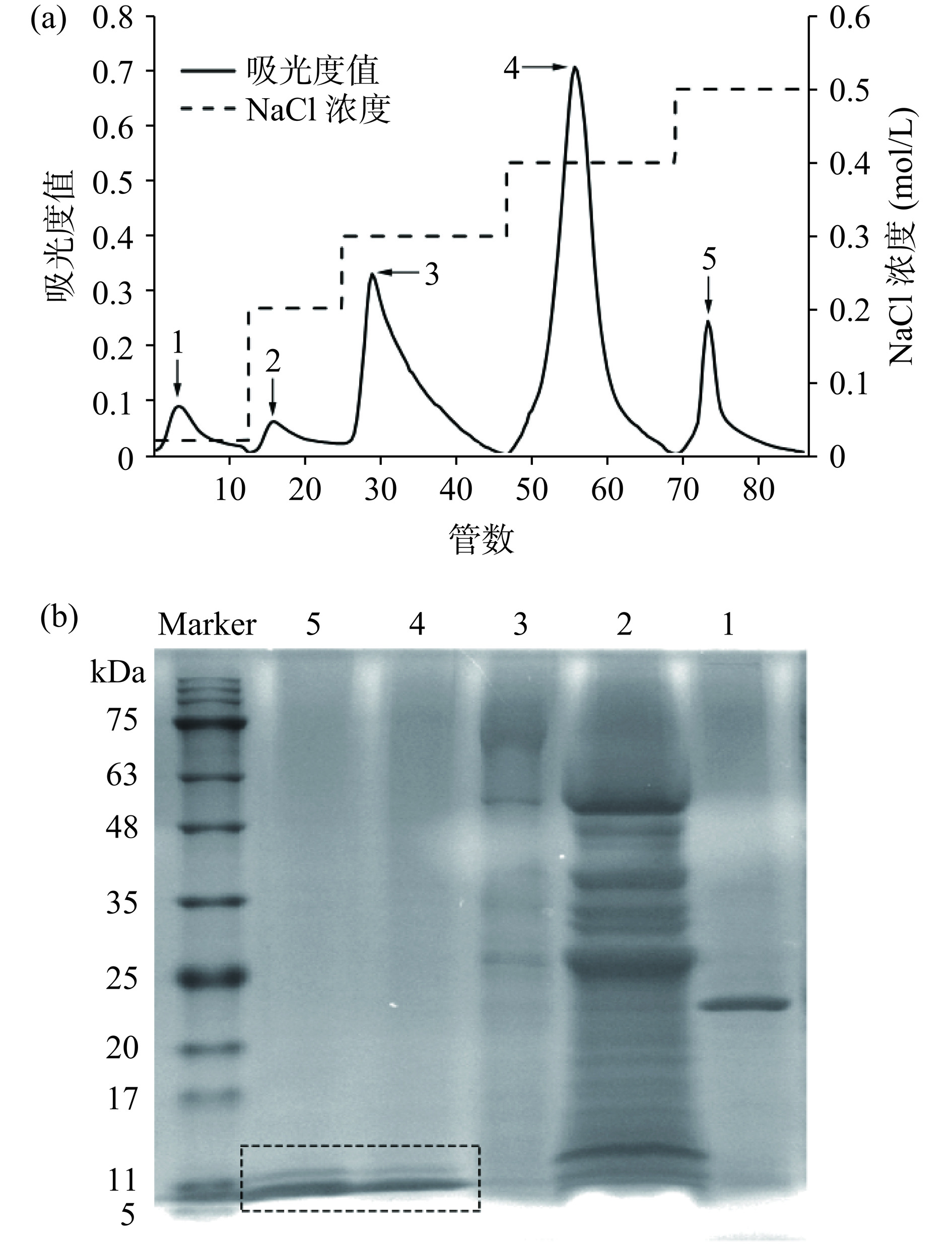
将AdMT粗提物进行DEAE阴离子交换层析(图3a),在0.02~0.3 mol/L较低NaCl浓度洗脱下,收集到吸附力不强的3个峰组分,在NaCl浓度为0.4 mol/L和0.5 mol/L时,洗脱下2个峰组分。对收集到的5个峰组分进行SDS-PAGE分析(图3b),低NaCl浓度洗脱下的蛋白组分(峰1~峰3)中具有较多杂蛋白,未检测到MT。经高浓度NaCl洗脱后,蛋白组分(峰4、峰5)中杂蛋白被清除,在分子量6 kDa处呈现明显条带,根据分子量判断为AdMT目标蛋白。同时,在12 kDa左右处出现一条微弱条带,这可能是Cys残基形成分子间二硫键,导致分子聚合形成AdMT二聚体[27]。由于两个组分中均具有目标蛋白,后续研究将其合并收集。这些结果表明,采用DEAE阴离子交换层析分离纯化可得到电泳纯的AdMT。
![]() 图 3 AdMT的DEAE-琼脂糖凝胶FF柱层析洗脱曲线(a)及15% SDS-PAGE图(b)Figure 3. DEAE sepharose fast flow column elution curve (a) and 15% SDS-PAGE patterns (b) of AdMT
图 3 AdMT的DEAE-琼脂糖凝胶FF柱层析洗脱曲线(a)及15% SDS-PAGE图(b)Figure 3. DEAE sepharose fast flow column elution curve (a) and 15% SDS-PAGE patterns (b) of AdMT2.5 生物活性分析
2.5.1 抗氧化活性
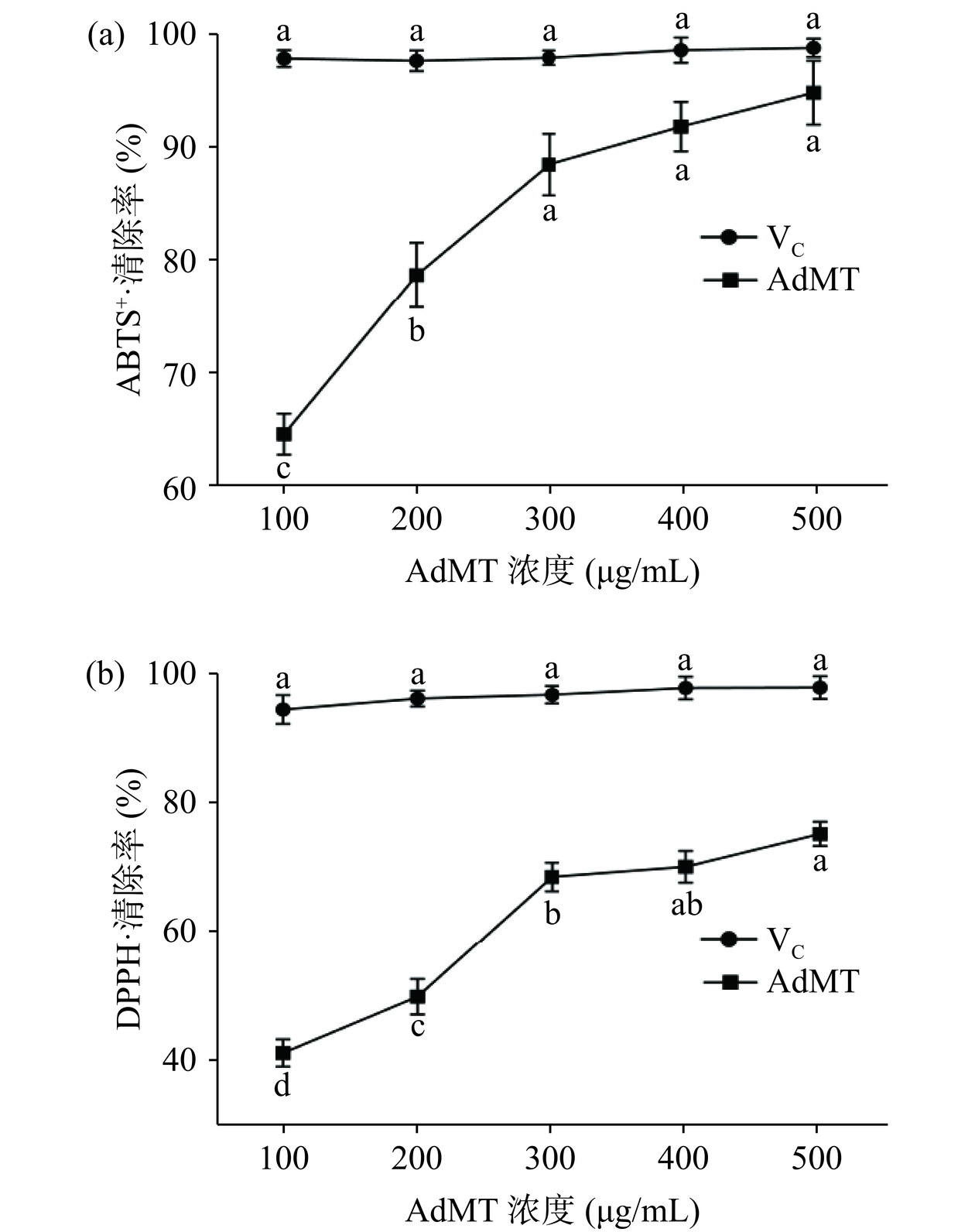
ABTS+和DPPH自由基清除率测定被广泛应用于活性物质的抗氧化能力检测,AdMT对该两种自由基的清除率如图4。由图4可知,AdMT对ABTS+和DPPH自由基清除率均随着浓度的升高而增强,呈剂量依赖特征。在浓度为100~300 μg/mL时,清除率增幅明显,在浓度为400~500 μg/mL时,增幅减弱。当AdMT浓度为500 μg/mL时,对ABTS+自由基的清除率达到95.01%,接近于对照组VC(99.03%),对DPPH自由基的最大清除率为75.25%,虽低于对照组(98.17%),但仍表现良好的抗氧化能力,与重组人金属硫蛋白-Ⅲα对DPPH自由基清除能力相当[28]。一般情况下,机体的调节能力可以使自由基保持动态平衡,但当体内产生的自由基数量超出自身的清除能力时,会引起细胞和组织发生氧化损伤,导致营养代谢和免疫调节能力降低,这与衰老及许多慢性疾病密切相关[29]。结果表明,AdMT具有良好的抗氧化能力,在机体抗氧化损伤方面具有一定潜力。
![]() 图 4 AdMT对ABTS+(a)和DPPH(b)自由基的清除率Figure 4. ABTS+ (a) and DPPH (b) scavenging activities of AdMT
图 4 AdMT对ABTS+(a)和DPPH(b)自由基的清除率Figure 4. ABTS+ (a) and DPPH (b) scavenging activities of AdMT2.5.2 细胞毒性及免疫原性
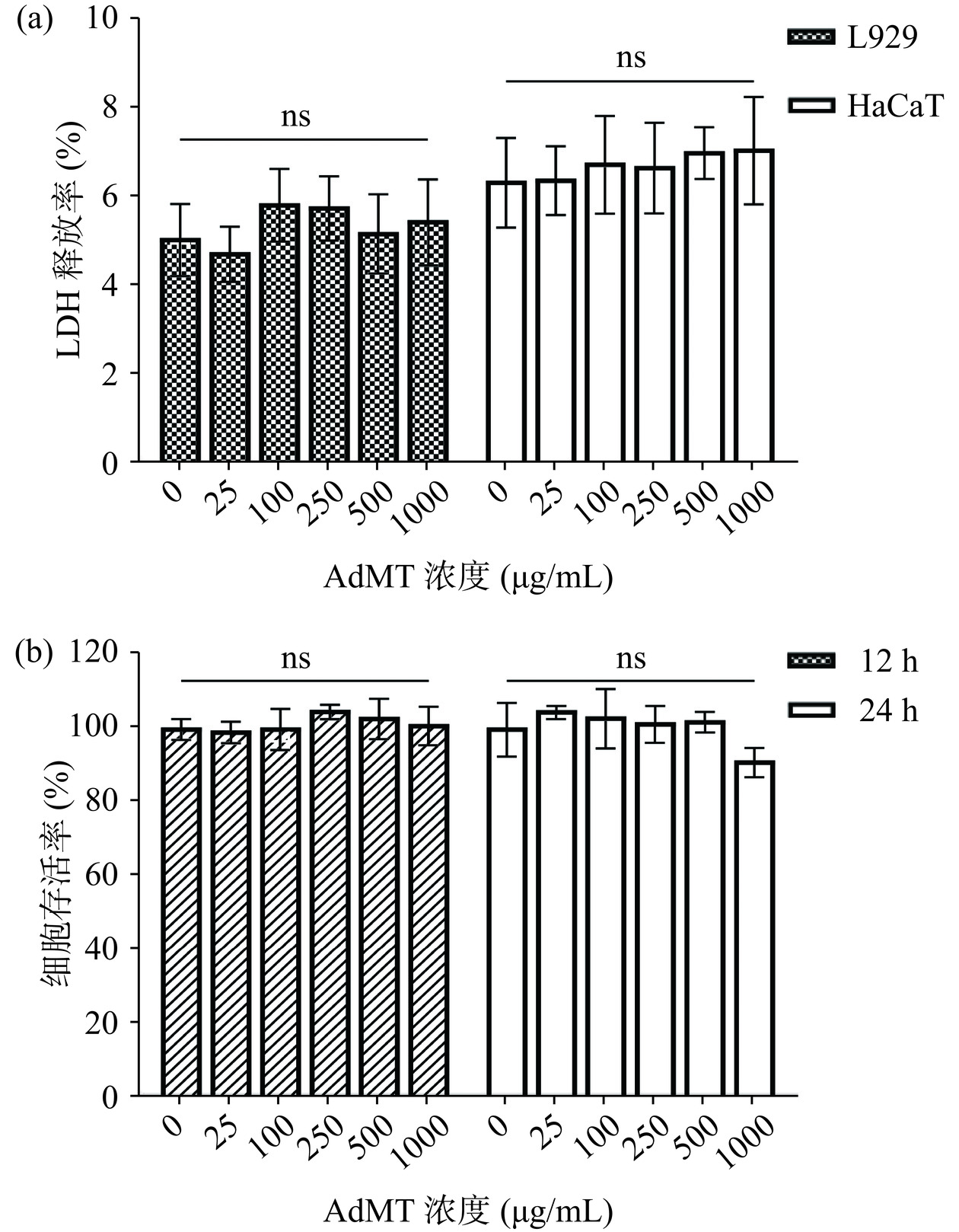
LDH释放率是基于细胞受损时,细胞膜发生损伤释放出乳酸脱氢酶而进行的检测[30]。不同浓度AdMT暴露对L929和HaCaT两种细胞的LDH释放水平如图5a。在1000 μg/mL AdMT范围内,两种细胞的LDH释放率均低于10%,且与对照组相比未呈现出显著差异(P>0.05),这表明AdMT对两种细胞的膜结构无损伤效应。外来病原体的入侵能够激活宿主的自身免疫反应,引起免疫细胞的异常增殖[31],若AdMT具有较高的免疫原性则会引起RAW264.7细胞的增殖。CCK-8检测不同浓度AdMT暴露后RAW264.7细胞的存活率,结果如图5b。AdMT既不引起RAW264.7细胞的增殖,也无显著致其死亡的作用,这说明AdMT不具有免疫原性。有研究表明,蛋白质的分子量大于10 kDa或含大量芳香族氨基酸时,可能具有较强的免疫原性[32],而AdMT分子量较小(6 kDa),且金属硫蛋白普遍不含有芳香族氨基酸,这也可能是其不具有免疫原性的原因。AdMT的无细胞毒性和免疫原性为其在食品、保健品、生物医药等领域的应用提供了基础。
![]() 图 5 AdMT的细胞毒性(a)和免疫原性(b)注:ns表示与对照组不具有显著统计学差异(P>0.05);图8同。Figure 5. Cytotoxicity (a) and immunogenicity (b) of AdMT
图 5 AdMT的细胞毒性(a)和免疫原性(b)注:ns表示与对照组不具有显著统计学差异(P>0.05);图8同。Figure 5. Cytotoxicity (a) and immunogenicity (b) of AdMT2.5.3 细胞增殖能力
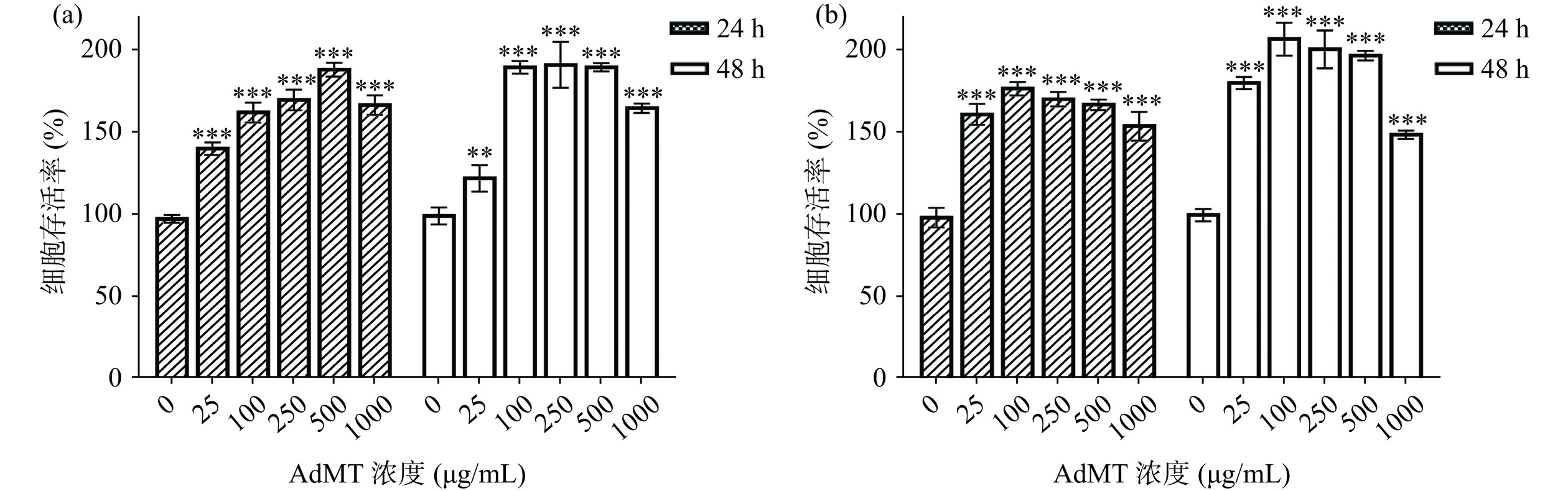
HaCaT细胞和L929细胞保持良好的活力和增殖能力,对表皮屏障功能及损伤修复功能具有十分重要的意义。以CCK-8法检测L929和HaCaT细胞活力(图6),结果表明,在不同浓度AdMT处理24或48 h后,与对照组相比,两种细胞的细胞存活率得到显著的提升(P<0.01,P<0.001),L929在250 μg/mL AdMT处理48 h后,细胞存活率达到192.63%,HaCaT在100 μg/mL AdMT处理48 h后,细胞存活率达到207.92%,均在100~500 μg/mL范围内无显著差异(P>0.05)。
![]()
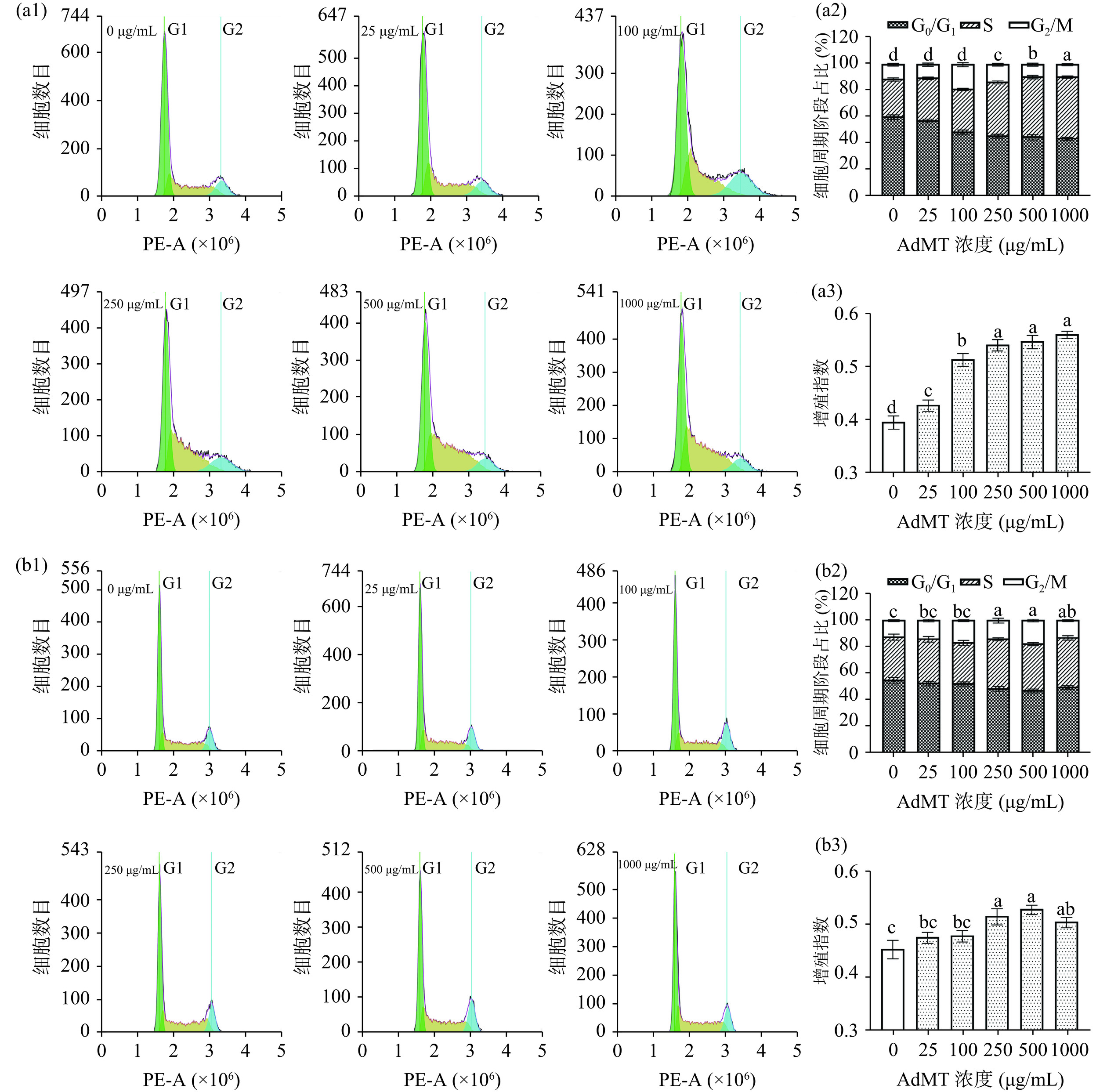
CCK-8实验反映细胞增殖的速度,而细胞周期则通过检测细胞中DNA的合成来检测细胞增殖能力,反映了AdMT对细胞所处周期阶段和细胞状态的直接影响。细胞周期中S期和G2/M期细胞的占比决定了细胞增殖的能力[33]。由图7可知,随着AdMT浓度的增加,L929处于G0/G1期的细胞占比逐渐降低,细胞增殖指数上升至0.56。对于HaCaT细胞,当浓度大于250 μg/mL时,其增殖指数与对照组相比显著增大(P<0.05),在500 μg/mL时达到0.53。这些结果表明,AdMT可有效提升L929和HaCaT细胞的活力,并通过增加S期和G2/M期的细胞占比促进细胞增殖。
![]() 图 7 AdMT对L929和HaCaT细胞周期及增殖指数的影响注:(a1)~(a3)为细胞L929;(b1)~(b3)为HaCaT细胞。Figure 7. Effects of AdMT on the cell cycle and proliferation index of L929 and HaCaT cells
图 7 AdMT对L929和HaCaT细胞周期及增殖指数的影响注:(a1)~(a3)为细胞L929;(b1)~(b3)为HaCaT细胞。Figure 7. Effects of AdMT on the cell cycle and proliferation index of L929 and HaCaT cells2.5.4 细胞迁移能力
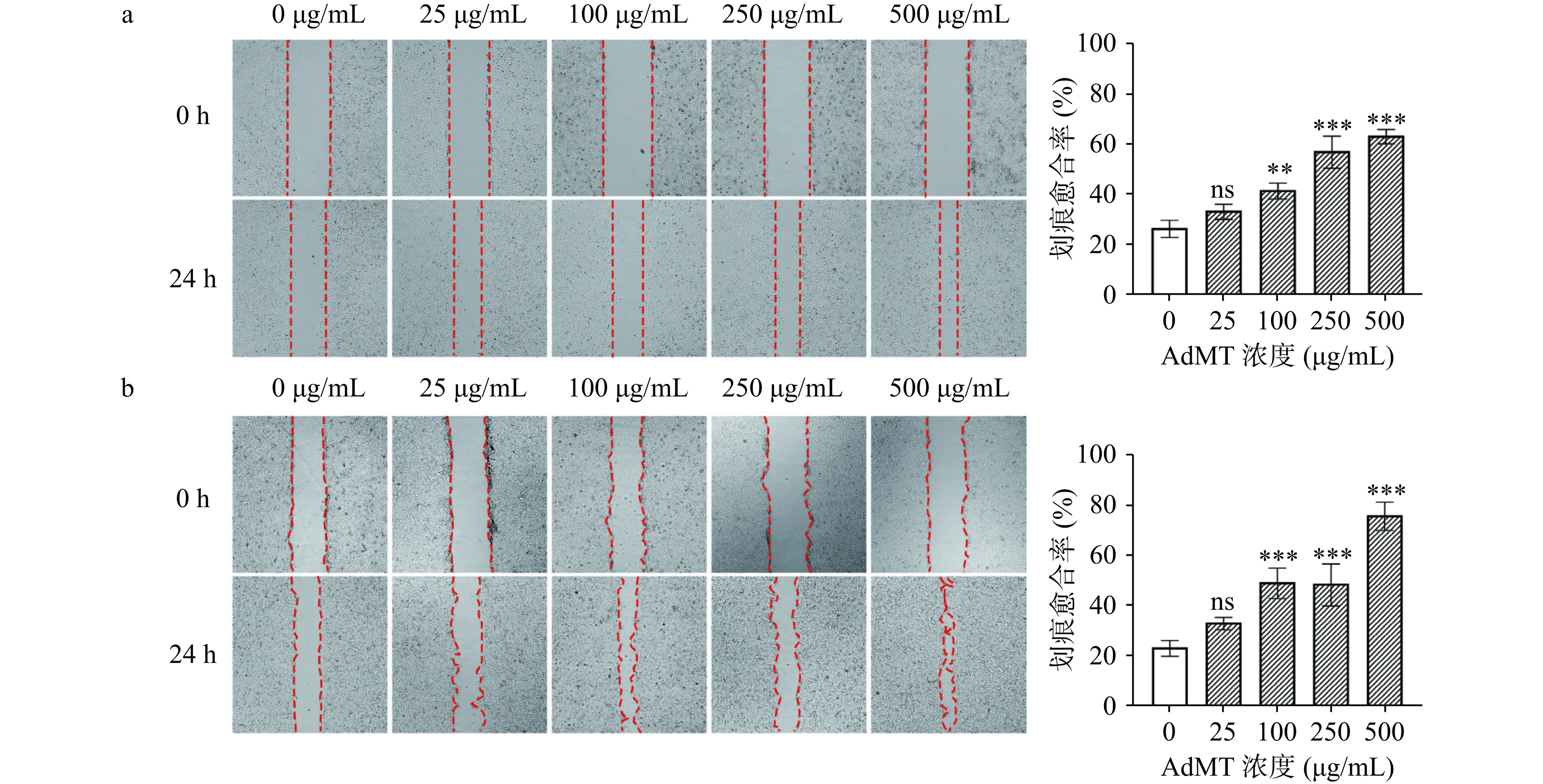
采用体外细胞划痕实验模拟皮肤伤口,测定细胞迁移运动与修复能力[34]。在用AdMT处理24 h后观察划痕愈合情况,如图8所示,AdMT促使细胞向划痕内迁移,随着AdMT浓度增加,细胞迁移能力增强,在500 μg/mL时,L929的划痕愈合率为63.38%,HaCaT的划痕愈合率达到76.23%,相较于对照组(分别为26.41%和23.15%)具有极显著差异(P<0.001)。AdMT对表皮细胞(HaCaT)和成纤维细胞(L929)的促进增殖和迁移作用表明,其在皮肤修复方面具有潜在应用价值。
![]() 图 8 AdMT对L929(a)和HaCaT(b)细胞迁移能力的影响Figure 8. Effects of AdMT on the migration ability of L929 (a) and HaCaT (b) cells
图 8 AdMT对L929(a)和HaCaT(b)细胞迁移能力的影响Figure 8. Effects of AdMT on the migration ability of L929 (a) and HaCaT (b) cells2.5.5 抗炎活性
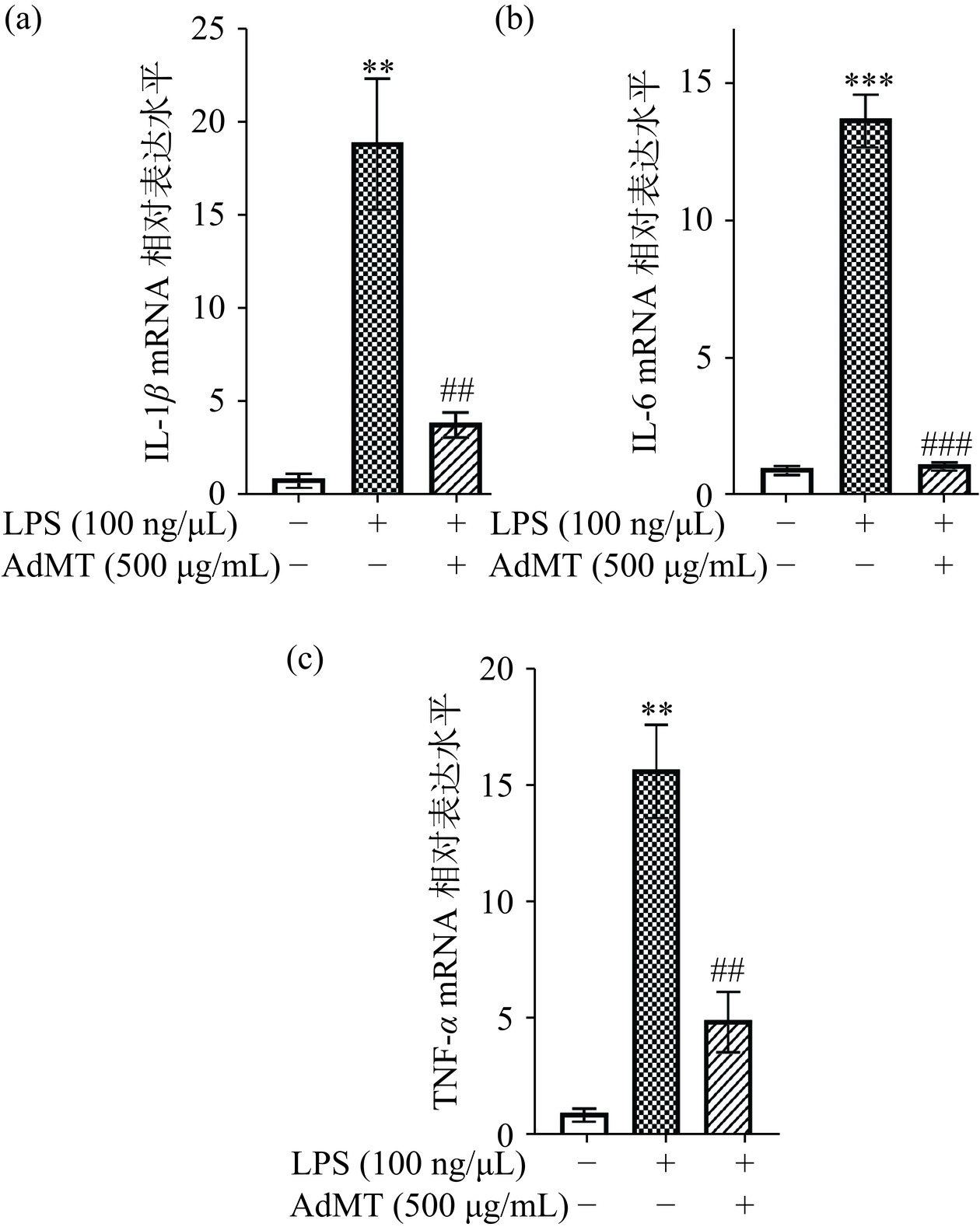
炎症反应是机体对有害因素入侵产生的一种生理防御反应,通过释放炎症因子和细胞因子来引起机体的炎症反应,但其过度释放会导致器官损伤和急性炎症性疾病的发生[35]。AdMT对LPS诱导的RAW264.7细胞中IL-1β、IL-6和TNF-α表达量的影响如图9所示。与对照组比较,LPS处理后,RAW264.7细胞中炎症因子IL-1β、IL-6、TNF-α的相对表达水平极显著升高(P<0.01),炎症模型构建成功,而后加入500 μg/mL AdMT共培养12 h,治疗炎症模型,结果显示,AdMT可高度显著降低炎症因子的相对表达水平(P<0.01),对LPS诱导的RAW264.7细胞炎症反应有良好的改善作用。
![]() 图 9 AdMT对LPS诱导的 RAW264.7 细胞中IL-1β(a)、IL-6(b)、TNF-α(c)表达量的影响注:*表示对照组与模型组统计学差异(显著*P<0.05,高度显著**P<0.01,极显著***P<0.001),#表示样品组与模型组统计学差异(显著#P<0.05,高度显著##P<0.01,极显著###P<0.001);图11同。Figure 9. Effects of AdMT on LPS-induced expression of IL-1β (a), IL-6 (b) and TNF-α (c) of RAW264.7 cells
图 9 AdMT对LPS诱导的 RAW264.7 细胞中IL-1β(a)、IL-6(b)、TNF-α(c)表达量的影响注:*表示对照组与模型组统计学差异(显著*P<0.05,高度显著**P<0.01,极显著***P<0.001),#表示样品组与模型组统计学差异(显著#P<0.05,高度显著##P<0.01,极显著###P<0.001);图11同。Figure 9. Effects of AdMT on LPS-induced expression of IL-1β (a), IL-6 (b) and TNF-α (c) of RAW264.7 cells2.5.6 重金属解毒能力
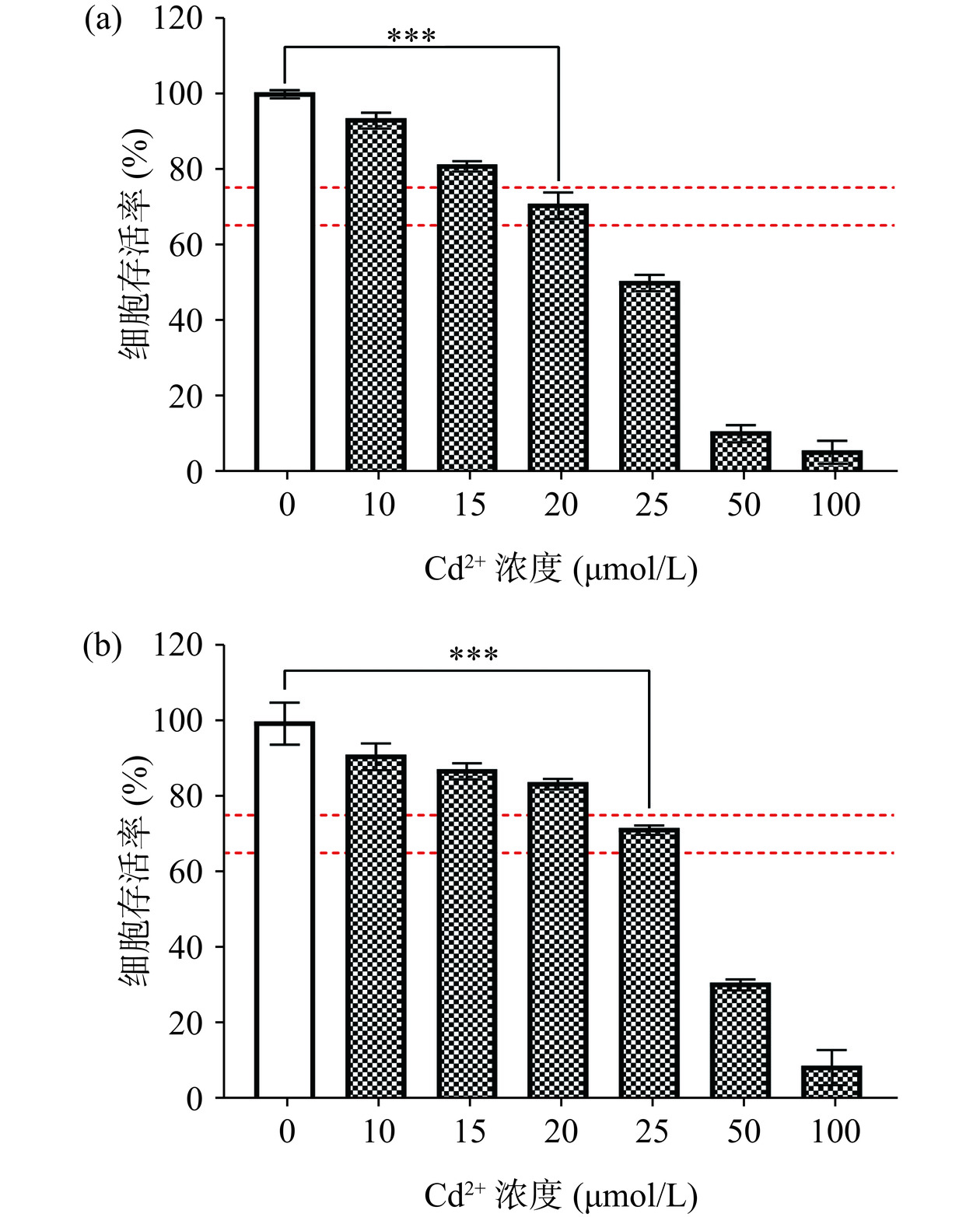
镉是工业上广泛使用的重金属,皮肤是镉暴露的重要靶器官[21]。细胞活力是细胞毒性的重要指标,可以反应细胞凋亡和死亡的情况。选用CdCl2构建L929和HaCaT细胞的重金属中毒模型,通过检测细胞存活率选择Cd2+暴露浓度,由图10可知,随着Cd2+浓度的升高,两种细胞的存活率均呈剂量依赖式下降。过高浓度Cd2+会造成细胞损伤严重,难以修复,因此选择细胞存活率极显著下降(P<0.001),且在65%~75%时的Cd2+浓度为模型暴露浓度,即分别为20 μmol/L(L929)和25 μmol/L(HaCaT),这个差异可能是由于不同细胞对重金属的耐受能力不同所致[36]。
![]() 图 10 L929(a)和HaCaT(b)细胞Cd2+中毒损伤模型Figure 10. Cd2+ toxic injury model of L929 (a) and HaCaT (b) cells
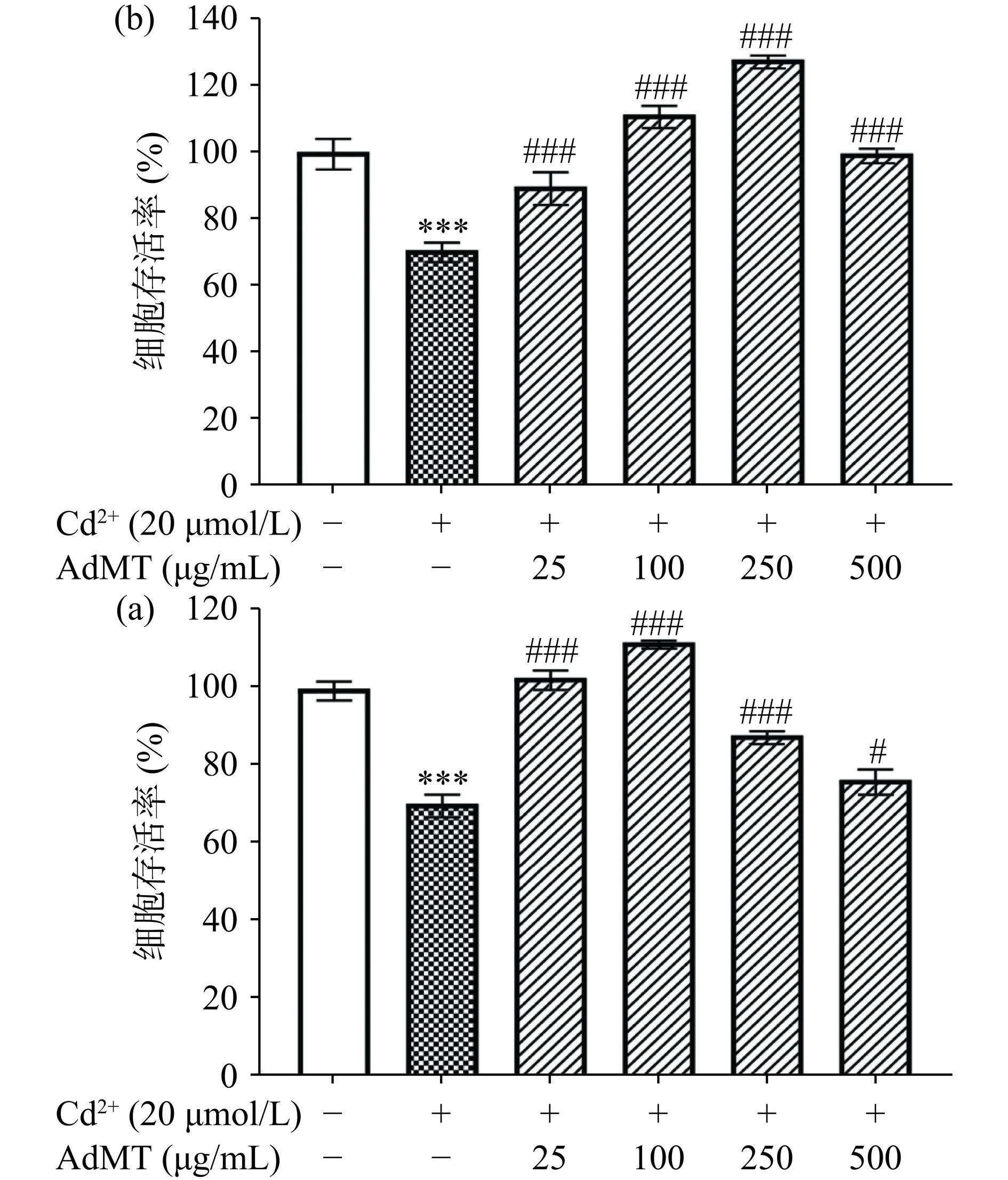
图 10 L929(a)和HaCaT(b)细胞Cd2+中毒损伤模型Figure 10. Cd2+ toxic injury model of L929 (a) and HaCaT (b) cells建立Cd2+重金属中毒模型后,加入不同浓度AdMT进行处理,结果显示(图11),AdMT在100 μg/mL时表现出最优异的修复L929的Cd2+中毒模型能力,而HaCaT细胞Cd2+中毒模型的最佳AdMT修复作用浓度为250 μg/mL,均可将细胞中毒模型的存活率修复至对照组水平,与Wahyudi等[37]使用姜黄素通过金属硫蛋白2A修复Cd2+诱导的HaCaT细胞中毒模型的结果一致。这些结果表明,AdMT对Cd2+诱导的L929和HaCaT细胞中毒损伤具有修复作用。
![]() 图 11 AdMT对L929(a)和HaCaT(b)细胞Cd2+中毒损伤的修复作用Figure 11. Repair effects of AdMT on Cd2+ toxic injury of L929 (a) and HaCaT (b) cells
图 11 AdMT对L929(a)和HaCaT(b)细胞Cd2+中毒损伤的修复作用Figure 11. Repair effects of AdMT on Cd2+ toxic injury of L929 (a) and HaCaT (b) cells3. 结论
本研究采用正交试验优化了大鲵金属硫蛋白的提取工艺,获得最优提取条件为液料比5:1、提取液浓度0.02 mol/L、提取温度40 ℃,提取时间1 h,在该条件下提取含量为0.362 mg/g,工艺有效可行,且含量高于常见水生动物;采用SDS-PAGE和FTIR,通过分子量和特征基团鉴定目标产物;随后采用DEAE阴离子交换层析分离纯化获得电泳纯AdMT。本实验条件下获得的AdMT具有良好的生物相容性,表现出较好的ABTS+和DPPH自由基清除能力,可以有效促进L929和HaCaT细胞增殖和迁移,降低炎症模型中IL-1β、IL-6和TNF-α的表达,同时显示对重金属Cd中毒模型的修复能力,说明AdMT具有优秀的抗氧化、抗炎、促进细胞增殖和迁移及重金属解毒生物活性,在抗氧化、皮肤修复、重金属解毒等食品、保健品、生物医药多方面具有一定开发利用潜力。
大鲵作为现存体型最大的两栖动物,其肝脏质量占比高且作为加工副产物未被充分利用,本研究结果表明,大鲵肝脏是一种含量丰富且价格低廉的优质金属硫蛋白提取原料,开发利用大鲵金属硫蛋白,不仅有利于减少资源浪费,同时可以有效提高大鲵产品的附加值,是大鲵产业可持续发展的新途径。但是,由于蛋白在体内的吸收代谢受多种因素影响,后续还需研究阐明具体的作用机制,进一步探究其在体内的吸收利用情况,推动大鲵金属硫蛋白的实际应用。
-
![]()
![]()
图 2 AdMT提取物(a)、15% SDS-PAGE图(b)及红外光谱扫描图(c)
Figure 2. AdMT extract (a), 15% SDS-PAGE patterns (b) and infrared spectral scanning (c) of AdMT
![]()
图 3 AdMT的DEAE-琼脂糖凝胶FF柱层析洗脱曲线(a)及15% SDS-PAGE图(b)
Figure 3. DEAE sepharose fast flow column elution curve (a) and 15% SDS-PAGE patterns (b) of AdMT
![]()
图 4 AdMT对ABTS+(a)和DPPH(b)自由基的清除率
Figure 4. ABTS+ (a) and DPPH (b) scavenging activities of AdMT
![]()
图 5 AdMT的细胞毒性(a)和免疫原性(b)
注:ns表示与对照组不具有显著统计学差异(P>0.05);图8同。
Figure 5. Cytotoxicity (a) and immunogenicity (b) of AdMT
![]()
![]()
图 7 AdMT对L929和HaCaT细胞周期及增殖指数的影响
注:(a1)~(a3)为细胞L929;(b1)~(b3)为HaCaT细胞。
Figure 7. Effects of AdMT on the cell cycle and proliferation index of L929 and HaCaT cells
![]()
图 8 AdMT对L929(a)和HaCaT(b)细胞迁移能力的影响
Figure 8. Effects of AdMT on the migration ability of L929 (a) and HaCaT (b) cells
![]()
图 9 AdMT对LPS诱导的 RAW264.7 细胞中IL-1β(a)、IL-6(b)、TNF-α(c)表达量的影响
注:*表示对照组与模型组统计学差异(显著*P<0.05,高度显著**P<0.01,极显著***P<0.001),#表示样品组与模型组统计学差异(显著#P<0.05,高度显著##P<0.01,极显著###P<0.001);图11同。
Figure 9. Effects of AdMT on LPS-induced expression of IL-1β (a), IL-6 (b) and TNF-α (c) of RAW264.7 cells
![]()
图 10 L929(a)和HaCaT(b)细胞Cd2+中毒损伤模型
Figure 10. Cd2+ toxic injury model of L929 (a) and HaCaT (b) cells
![]()
图 11 AdMT对L929(a)和HaCaT(b)细胞Cd2+中毒损伤的修复作用
Figure 11. Repair effects of AdMT on Cd2+ toxic injury of L929 (a) and HaCaT (b) cells
表 1 正交试验因素与水平
Table 1 Factors and levels of orthogonal tests
水平 A液料比
(mL:g)B提取液浓度
(mol/L)C提取温度
(℃)D提取时间
(h)1 3:1 0.01 30 1.0 2 4:1 0.02 40 1.5 3 5:1 0.05 50 2.0  下载: 导出CSV
下载: 导出CSV
表 2 RT-qPCR引物
Table 2 Primers sequences used for RT-qPCR
引物名称 引物序列 IL-1β F:CCAGGATGAGGACCCAAGCA;
R:TCCCGACCATTGCTGTTTCCIL-6 F:GAGTCCTTCAGAGAGATACAG;
R:CTGTGACTCCAGCTTATCTGTNF-α F:GTCTCAGCCTCTTCTCATTC;
R:CATAGAACTGATGAGAGGGAGAPDH F:GTGGCAAAGTGGAGATTGTTG;
R:CTCCTGGAAGATGGTGATGG
下载: 导出CSV
表 3 正交试验结果
Table 3 Results of orthogonal tests
实验号 液料比 提取液浓度 提取温度 提取时间 AdMT提取
含量(mg/g)1 1 1 1 1 0.334 2 1 2 3 2 0.344 3 1 3 2 3 0.339 4 2 1 3 3 0.348 5 2 2 2 1 0.357 6 2 3 1 2 0.341 7 3 1 2 2 0.351 8 3 2 3 3 0.346 9 3 3 1 1 0.352 K 1 1.02 1.03 1.02 1.04 2 1.05 1.05 1.05 1.04 3 1.05 1.03 1.04 1.03 k 1 0.34 0.34 0.34 0.35 2 0.35 0.35 0.35 0.35 3 0.35 0.34 0.35 0.34 最佳水平 3 2 2 1 R 0.54 0.02 0.25 0.11
下载: 导出CSV
-
[1] 蒋万胜, 兰香英, 王金秀, 等. 中国大鲵种质资源保护与利用研究进展[J]. 水产学报,2022,46(4):683−705. [JIANG W S, LAN X Y, WANG J X, et al. Research progress on the conservation and utilization of germplasm resources of Giant Salamander in China[J]. Journal of Fisheries of China,2022,46(4):683−705.] JIANG W S, LAN X Y, WANG J X, et al. Research progress on the conservation and utilization of germplasm resources of Giant Salamander in China[J]. Journal of Fisheries of China, 2022, 46(4): 683−705.
[2] ZHANG P, DONG X, GRENOUILLET G, et al. Species range shifts in response to climate change and human pressure for the world's largest amphibian[J]. The Science of the Total Environment,2020,735:139543. doi: 10.1016/j.scitotenv.2020.139543
[3] 易凡智, 刘金洋, 翟兴月, 等. 大鲵水解胶原的制备、生物活性及护肤作用研究进展[J]. 广东化工,2023,50(16):89−91. [YI F Z, LIU J Y, ZHAI X Y, et al. Advanced on preparation, biological activities and skin care effects of hydrolyzed collagen from Andrias Davidianus[J]. Guangdong Chemical Industry,2023,50(16):89−91.] YI F Z, LIU J Y, ZHAI X Y, et al. Advanced on preparation, biological activities and skin care effects of hydrolyzed collagen from Andrias Davidianus[J]. Guangdong Chemical Industry, 2023, 50(16): 89−91.
[4] 曾允灏, 苏文, 郑小莉, 等. 大鲵肝肽酒制备及其对小鼠酒后肝脏保护作用研究[J]. 中国酿造,2023,42(2):120−125. [ZENG Y H, SU W, ZHENG X L, et al. Preparation of Andrias davidianus liver peptide wine and its protective effect on mouse liver after drinking[J]. Chinese Brewing,2023,42(2):120−125.] ZENG Y H, SU W, ZHENG X L, et al. Preparation of Andrias davidianus liver peptide wine and its protective effect on mouse liver after drinking[J]. Chinese Brewing, 2023, 42(2): 120−125.
[5] MERLOS R M A, JIMENEZ J A M, HADDAD Y, et al. Metallothionein isoforms as double agents-Their roles in carcinogenesis, cancer progression and chemoresistance[J]. Drug Resistance Updates:Reviews and Commentaries in Antimicrobial and Anticancer Chemotherapy,2020,52:100691.
[6] LIU Y, WU Z, GUO K, et al. Metallothionein-1 gene from Exopalaemon carinicauda and its response to heavy metal ions challenge[J]. Marine Pollution Bulletin,2022,175:113324. doi: 10.1016/j.marpolbul.2022.113324
[7] DAI H, WANG L, LI L, et al. Metallothionein 1:A new spotlight on inflammatory diseases[J]. Frontiers in Immunology,2021,12:739918. doi: 10.3389/fimmu.2021.739918
[8] 周小清, 戴玉洁, 黄林芳, 等. 大体积样品堆积-毛细管电泳-紫外检测分析鼠肝中不同亚型金属硫蛋白[J]. 分析科学学报,2017,33(5):593−599. [ZHOU X Q, DAI Y J, HUANG L F, et al. Large volume sample stacking-capillary electrophoresis-ultraviolet detection for analysis of metallothionein isoforms in rats' liver[J]. Journal of Analytical Science,2017,33(5):593−599.] ZHOU X Q, DAI Y J, HUANG L F, et al. Large volume sample stacking-capillary electrophoresis-ultraviolet detection for analysis of metallothionein isoforms in rats' liver[J]. Journal of Analytical Science, 2017, 33(5): 593−599.
[9] 李丽立. 一种猪肝金属硫蛋白的提取方法:中国,200410013212. X[P]. [LI L L. A method for extracting metallothionein from pig liver:China,200410013212.X[P].] LI L L. A method for extracting metallothionein from pig liver: China, 200410013212.X[P].
[10] 严国, 梅光明, 顾捷, 等. 口虾蛄金属硫蛋白提取工艺的优化[J]. 食品与发酵工业,2019,45(15):108−114. [YAN G, MEI G M, GU J, et al. Optimization of metallothionein extraction process for Oratosquilla oratoria[J]. Food and Fermentation Industries,2019,45(15):108−114.] YAN G, MEI G M, GU J, et al. Optimization of metallothionein extraction process for Oratosquilla oratoria[J]. Food and Fermentation Industries, 2019, 45(15): 108−114.
[11] 姚冬儿, 杨慧琳, 斯兴开, 等. 基于响应面优化的褶牡蛎中金属硫蛋白提取工艺研究[J]. 现代食品科技,2018,34(2):189−196,245. [YAO D E, YANG H L, SI X K, et al. Optimization for the extraction process of metallothionein from oyster (Crassostrea plicatula) based on response surface methodology[J]. Modern Food Science and Technology,2018,34(2):189−196,245.] YAO D E, YANG H L, SI X K, et al. Optimization for the extraction process of metallothionein from oyster (Crassostrea plicatula) based on response surface methodology[J]. Modern Food Science and Technology, 2018, 34(2): 189−196,245.
[12] 孟春英, 李停停, 黄丽英, 等. 虾夷扇贝内脏源金属硫蛋白的提取工艺优化[J]. 食品工业科技,2019,40(5):176−181. [MENG C Y, LI T T, HUANG L Y, et al. Optimization of extraction technology of visceral metallothionein from Patinopecten yess-oensis[J]. Science and Technology of Food Industry,2019,40(5):176−181.] MENG C Y, LI T T, HUANG L Y, et al. Optimization of extraction technology of visceral metallothionein from Patinopecten yess-oensis[J]. Science and Technology of Food Industry, 2019, 40(5): 176−181.
[13] PABIS K, CHIARI Y, SALA C, et al. Elevated metallothionein expression in long-lived species mediates the influence of cadmium accumulation on aging[J]. Gero Science,2021,43(4):1975−1993.
[14] 吴云辉, 王俊坤, 孙继鹏, 等. DTNB比色法测定金属硫蛋白含量[J]. 食品科学,2013,34(16):196−199. [WU Y H, WANG J K, SUN J P, et al. Colormetric determination of metallothionein using DTNB[J]. Food Science,2013,34(16):196−199.] WU Y H, WANG J K, SUN J P, et al. Colormetric determination of metallothionein using DTNB[J]. Food Science, 2013, 34(16): 196−199.
[15] 何蜀峰, 李孟孟, 孙杨赢. 水解对低盐鸭肉肌原纤维蛋白结构和功能特性的影响[J]. 食品工业科技,2024,45(2):84−91. [HE S F, LI M M, SUN Y Y. Effect of hydrolysis on structure and properties of duck myofibrillar protein inlow-salt condition[J]. Science and Technology of Food Industry,2024,45(2):84−91.] HE S F, LI M M, SUN Y Y. Effect of hydrolysis on structure and properties of duck myofibrillar protein inlow-salt condition[J]. Science and Technology of Food Industry, 2024, 45(2): 84−91.
[16] 李停停. 毛蚶中金属硫蛋白的提取纯化及抗氧化活性分析[D]. 舟山:浙江海洋大学, 2019. [LI T T. The extraction and purification and antioxidant activity analysis of metallothionein from Scapharca subcrenata[D]. Zhoushan:Zhejiang Ocean University, 2019.] LI T T. The extraction and purification and antioxidant activity analysis of metallothionein from Scapharca subcrenata[D]. Zhoushan: Zhejiang Ocean University, 2019.
[17] PARK S K, LEE Y K. Antioxidant activity in rheum emodi wall (Himalayan Rhubarb)[J]. Molecules (Basel, Switzerland),2021,26(9):2555. doi: 10.3390/molecules26092555
[18] HAO W, LI K, MA Y, et al. Preparation and antioxidant activity of chitosan dimers with different sequences[J]. Marine drugs,2021,19(7):366. doi: 10.3390/md19070366
[19] 孙左义, 原海亮, 秦建新, 等. 重组人金属硫蛋白-Ⅲ对细胞和秀丽隐杆线虫毒性效应的研究[J]. 中国细胞生物学学报,2022,44(7):1339−1348. [SUN Z Y, YUAN H L, QIN J X, et al. Toxic effects of genetically engineered recombinant human metallothionein-I on cells and Caenorhabditis elegans[J]. Chinese Journal of Cell Biology,2022,44(7):1339−1348.] SUN Z Y, YUAN H L, QIN J X, et al. Toxic effects of genetically engineered recombinant human metallothionein-I on cells and Caenorhabditis elegans[J]. Chinese Journal of Cell Biology, 2022, 44(7): 1339−1348.
[20] 朱彦彬, 徐鑫婷, 黎燕冰, 等. 桦褐孔菌醇抑制LPS诱导RAW264.7细胞炎症反应[J]. 食品工业科技,2023,44(19):401−409. [ZHU Y B, XU X T, LI Y B, et al. Obliquin inhibits LPS-induced inflammatory response in RAW264.7 cells[J]. Science and Technology of Food Industry,2023,44(19):401−409.] ZHU Y B, XU X T, LI Y B, et al. Obliquin inhibits LPS-induced inflammatory response in RAW264.7 cells[J]. Science and Technology of Food Industry, 2023, 44(19): 401−409.
[21] MOON S H, LEE C M, NAM M J. Cytoprotective effects of taxifolin against cadmium-induced apoptosis in human keratinocytes[J]. Human & Experimental Toxicology,2019,38(8):992−1003.
[22] 黄楚森, 朱维平, 徐玉芳, 等. 蛋白质巯基及其氧化性修饰的化学检测方法[J]. 药学学报,2012,47(3):280−290,258. [HUANG C S, ZHU W P, XU Y F, et al. Chemical approaches for trapping protein thiols and their oxidative modification[J]. Acta Pharmaceutica Sinica,2012,47(3):280−290,258.] HUANG C S, ZHU W P, XU Y F, et al. Chemical approaches for trapping protein thiols and their oxidative modification[J]. Acta Pharmaceutica Sinica, 2012, 47(3): 280−290,258.
[23] 于立博, 刘继文. 维药鹰嘴豆金属硫蛋白的提取检测[J]. 时珍国医国药,2013,24(5):1095−1096. [YU L B, LIU J W. Extraction and detection of metallothionein in chickpea[J]. Lishizhen Medicine and Materia Medica Research,2013,24(5):1095−1096.] YU L B, LIU J W. Extraction and detection of metallothionein in chickpea[J]. Lishizhen Medicine and Materia Medica Research, 2013, 24(5): 1095−1096.
[24] 严国. 口虾蛄金属硫蛋白的提取与纯化制备工艺研究[D]. 舟山:浙江海洋大学, 2019. [YAN G. Study on extraction and purification of metallothionein from Oratosquilla oratoria[D]. Zhoushan:Zhejiang Ocean University, 2019.] YAN G. Study on extraction and purification of metallothionein from Oratosquilla oratoria[D]. Zhoushan: Zhejiang Ocean University, 2019.
[25] LI X, REN Z, CRABBE M J C, et al. Genetic modifications of metallothionein enhance the tolerance and bioaccumulation of heavy metals in Escherichia coli[J]. Ecotoxicology and Environmental Safety,2021,222:112512. doi: 10.1016/j.ecoenv.2021.112512
[26] MENG C, WANG K, ZHANG X, et al. Purification and structure analysis of zinc-binding protein from Mizuhopecten yess-oensis[J]. Journal of Food Biochemistry,2021,45(6):e13756.
[27] 黄婧筠. 小球藻锌结合类金属硫蛋白的分离提取及其对重金属镉的螯合作用研究[D]. 厦门:集美大学, 2017. [HUANG J J. Extraction of zinc-binding metallothionein-like proteins induced in C. vugularis and its chelation to cadmium[D]. Xiamen:Jimei University, 2017.] HUANG J J. Extraction of zinc-binding metallothionein-like proteins induced in C. vugularis and its chelation to cadmium[D]. Xiamen: Jimei University, 2017.
[28] 甘俊英, 孙左义, 秦建新, 等. 重组人金属硫蛋白-Ⅲα短肽对UVB致HaCaT细胞氧化损伤的缓解作用[J]. 生态毒理学报,2021,16(4):240−249. [GAN J Y, SUN Z Y, QIN J X, et al. Allievation effects of recombinant human metallothionein Ⅲα peptide on oxidative damage of HaCaT cells induced by UVB[J]. Asian Journal of Ecotoxicology,2021,16(4):240−249.] GAN J Y, SUN Z Y, QIN J X, et al. Allievation effects of recombinant human metallothionein Ⅲα peptide on oxidative damage of HaCaT cells induced by UVB[J]. Asian Journal of Ecotoxicology, 2021, 16(4): 240−249.
[29] HU Q, LIU Z, GUO Y, et al. Antioxidant capacity of flavonoids from Folium Artemisiae argyi and the molecular mechanism in Caenorhabditis elegans[J]. Journal of Ethnopharmacology,2021,279:114398. doi: 10.1016/j.jep.2021.114398
[30] VAN D B S, VANDEPLASSCHE E, OSTYN L, et al. Bacterial interference with lactate dehydrogenase assay leads to an underestimation of cytotoxicity[J]. Frontiers in Cellular and Infection Microbiology,2020,10:494. doi: 10.3389/fcimb.2020.00494
[31] NING X, WANG Y, JING M, et al. Apoptotic caspases suppress type i interferon production via the cleavage of cGAS, MAVS, and IRF3[J]. Molecular Cell, 2019, 74(1):19−31. e7.
[32] GREENFIELD E A, DECAPRIO J, BRAHMANDAM M. Selecting the Antigen[J]. Cold Spring Harbor Protocols, 2021, 2021(12).
[33] BERTOLI C, SKOTHEIM J M, DE B R A. Control of cell cycle transcription during G1 and S phases[J]. Nature Reviews Molecular Cell Biology,2013,14(8):518−528. doi: 10.1038/nrm3629
[34] DING C, PENG X, YANG J, et al. Rg3-loaded P407/CS/HA hydrogel inhibits UVB-induced oxidative stress, inflammation and apoptosis in HaCaT cells[J]. Biomedicine & Pharmacotherapy,2023,165:115177.
[35] HUANG C, DONG L, ZHAO B, et al. Anti-inflammatory hydrogel dressings and skin wound healing[J]. Clinical and Translational Medicine,2022,12(11):e1094. doi: 10.1002/ctm2.1094
[36] LI J Y, CUI D L, XIE Y M, et al. Mechanisms of Cd-induced cytotoxicity in normal human skin keratinocytes:Implication for human health[J]. International Journal of Molecular Sciences,2022,23(19):11767. doi: 10.3390/ijms231911767
[37] WAHYUDI L D, YU S H, CHO M K. The effect of curcumin on the cadmium-induced mitochondrial apoptosis pathway by metallothionein 2A regulation[J]. Life Sciences,2022,310:121076. doi: 10.1016/j.lfs.2022.121076
-
期刊类型引用(1)
1. 章紫英 ,周汉昌 ,黄思远 ,李婷 ,余林 ,况小宝 . 烟熏对食品风味及安全性影响研究进展. 食品工业. 2024(11): 270-276 .  百度学术
百度学术
其他类型引用(0)
 下载:
下载:











计量
- 文章访问数: 59
- HTML全文浏览量: 10
- PDF下载量: 9
- 被引次数: 1



