


Gene Expression and Activity Change Analysis of Cell Wall Degrading Enzyme in Postharvest MelonInfected by Alternaria alternata
-
摘要: 目的:探讨链格孢菌侵染采后甜瓜果实过程中细胞壁降解酶活性及基因表达的变化规律,揭示链格孢菌的致病机理。方法:以伽师瓜和86-1甜瓜果实为实验材料,损伤接种链格孢菌(Alternaria alternata),室温(25 ℃),相对湿度35%~40%贮藏,测定病斑直径,观察病斑组织显微结构,测定果胶裂解酶(Pectin lyase,PL)、纤维素酶(Cellulase,Cx)、β-葡萄糖苷酶(β-Glucosidase,CB)、多聚半乳糖醛酸酶(Polygalacturonase,PG),采用荧光定量聚合酶链式反应(Polymerase chain reaction,PCR)的绝对定量方法对AaPL、AaCx、AaPG、AaCB表达量的变化进行分析,并对四种细胞壁降解酶的活性与其对应基因的表达量进行相关性分析。结果:在相同的侵染天数下,在86-1甜瓜果肉、果皮组织的病斑直径大于伽师瓜;观察病斑组织显微结构,对比两种甜瓜,链格孢菌在86-1甜瓜细胞中繁殖更快,对86-1甜瓜细胞结构的破坏能力更强;比较四种细胞壁降解酶在两种甜瓜上活性和基因表达量的变化规律,相同天数下总体来说在86-1甜瓜接种组中的酶活性和基因表达量更高,在伽师瓜接种组中活性、表达量达到峰值的天数更晚。结论:在侵染过程中由于甜瓜自身抗病机制,使细胞壁降解酶活性及基因表达的变化趋势在6~10 d有所波动;链格孢菌在86-1甜瓜中的侵染能力更强,也体现了伽师瓜对链格孢菌侵染的抵抗能力更强。Abstract: Objective: To investigate the changes of cell wall degrading enzyme activity and gene expression during the infection of postharvest melon fruit by Alternaria alternata, and to reveal the pathogenic mechanism of Alternaria alternata. Methods: Jiashi melon and 86-1 melon fruits were used as experimental materials. Alternaria alternata was inoculated and stored at room temperature (25 ℃) and relative humidity of 35%~40%. The diameter of lesions was measured, and the microstructure of lesions was observed. Pectin lyase (PL), cellulase (Cx), β-glucosidase (CB) and polygalacturonase (PG) were determined. The expression changes of AaPL, AaCx, AaPG and AaCB were analyzed by absolute quantitative method of fluorescence quantitative polymerase chain reaction (PCR), and the correlation between the activity of four cell wall degrading enzymes and the expression of their corresponding genes was analyzed. Results: Under the same infection days, the lesion diameter of 86-1 melon pulp and peel tissue was larger than that of Jiashi melon. The microstructure of the lesion tissue was observed. Compared with the two melons, A. alternata propagated faster in 86-1 melon cells and had a stronger ability to destroy the cell structure of 86-1 melon. The changes of activity and gene expression of four cell wall degrading enzymes on two melons were compared. Under the same number of days, the enzyme activity and gene expression were higher in the 86-1 melon inoculation group, and the number of days when the activity and expression reached the peak in the Jiashi melon inoculation group was later. Conclusion: During the infection process, due to the melon's own disease resistance mechanism, the change trend of cell wall degrading enzyme activity and gene expression fluctuated from 6 to 10 days. The infection ability of Alternaria alternata in 86-1 melon was stronger, which also reflected that Jiashi melon had stronger resistance to Alternaria alternata infection.
-
Keywords:
- Alternaria alternata /
- melon /
- cell wall degrading enzyme /
- gene expression /
- enzyme activity
-
甜瓜(Cucumis melo L.)富含糖、维生素、抗坏血酸、叶酸等多种营养物质,广受世界各地消费者的喜爱[1]。2020年,我国甜瓜栽培面积为38.78万公顷、产量达138.65万吨,种植面积及产量均列世界前茅(数据来源为联合国粮农组织统计数据库[2])。病原真菌的侵染是引起甜瓜病害的主要原因之一[3]。链格孢菌作为常见的植物病原真菌,能够引起伽师瓜等厚皮甜瓜的黑斑病,严重影响果实品质,在贮运过程中增加了残次果的产生,最终造成巨大的经济损失[4]。病原微生物在侵染植物宿主时会被植物细胞壁所抵挡。病原微生物为顺利侵入寄主细胞,便在侵染过程中合成的用以分解植物细胞壁的一系列的酶,统称为细胞壁降解酶,主要含有果胶酶、纤维素酶、半纤维素酶、多聚半乳糖醛酸等,细胞壁降解酶的活性与病原菌致病力的关系也呈现正相关[5−6]。实时荧光定量PCR(real time flurescent quantitive polymerase chain reaction,qPCR),作为一种成熟的测定基因表达量的方式,已被应用到医药、农学等各个领域[7],根据定量形式可以分为绝对定量和相对定量,相较于相对定量而言,绝对定量具有可以准确获取样本中所测基因的拷贝数[8−9]。
目前,对于甜瓜黑斑病的发病机理主要集中在甜瓜自身抗病能力的研究,如甜瓜果皮网纹大小对抗病能力的影响[10]、不同组织抗病能力的差异[11]、抗菌物质(黄酮、总酚)的变化规律[12]、病程相关蛋白及苯丙烷代谢酶的基因表达及活性变化规律[13−14],以链格孢菌的致病机制为主要视角的研究较少;而对于作物病原菌致病基因表达变化的测定多数采取real-time PCR相对定量的方法,鲜有采用绝对定量的方法。
实验以伽师瓜与86-1甜瓜作为试验对象,链格孢菌损伤侵染甜瓜,测定果实硬度和病斑直径、观察甜瓜组织显微结构;测定侵染过程中果胶裂解酶(Pectin lyase,PL)[EC 4.2.2.10]、纤维素酶(Cellulase,Cx)[EC 3.2.1.91]、β-葡萄糖苷酶(β-Glucosidase,CB)[EC 3.2.1.21]、多聚半乳糖醛酸酶(Polygalacturonase,PG)[EC 3.2.1.15]的活性;并通过本课题组前期对链格孢菌一些细胞壁降解酶基因克隆测序结果作为参考设计引物,采用荧光定量聚合酶链式反应(polymerase chain reaction,PCR)绝对定量的方法对AaPL、AaCx、AaPG、AaCB表达量变化进行分析,寻找酶活性变化规律与基因表达变化之间的规律,进一步揭示链格孢菌的致病机理。
1. 材料与方法
1.1 材料与仪器
86-1甜瓜、伽师瓜 均来自新疆伽师县,果实发育良好、未发病、无损伤伽师瓜质量4.0±0.5 kg,硬度0.55±0.05 kg/cm2,边缘可溶性固形物(10±1)°Brix;86-1甜瓜质量3.5±0.5 kg,硬度0.40±0.05 kg/cm2,边缘可溶性固形物(11.5±1)°Brix;链格孢菌 分离于自然发病的伽师瓜黑斑病组织,纯化后经ITS鉴定确定为链格孢菌(Alternaria alternata),马铃薯葡萄糖琼脂培养基(potato dextrose agar,PDA)28 ℃培养,4 ℃保藏;无水乙醇、硫酸镁、3,5-二硝基水杨酸、结晶酚、酒石酸钾钠、L-抗坏血酸、无水乙酸钠、浓盐酸 分析纯,天津市致远化学试剂有限公司;PDA培养基 北京奥博星生物技术有限责任公司;六胺银染液 武汉谷歌生物科技有限公司;真菌总RNA快速抽提试剂盒、无内毒素质粒DNA小提取试剂盒、琼脂糖 生工生物工程(上海);5× All-In-One RT MasterMix、EvaGreen Express 2×qPCR MasterMix-Low Rox ABM国际科技集团有限公司;M5 HiClear DL2000 DNA marker 北京聚合美生物科技有限公司。
ME204型电子天平、FE20型pH计 梅特勒-托利多仪器有限公司;TGL-16G型高速冷冻离心机 上海安亭科学仪器厂;TU-1810PC型紫外-可见分光光度计 北京普析通用公司;Nefuqe 15R高速分散器 宁波新芝生物科技股份有限公司;Nefuqe 15R高速冷冻离心机 上海力申科学仪器有限公司;GL-88B漩涡混合器 海门市其林贝尔仪器制造有限公司;K20型干式恒温器 杭州美盛仪器有限公司;Nano-100微量分光光度计 杭州奥盛仪器有限公司;DYY-6C型电泳仪电源、DYCZ-21电泳槽 北京市六一仪器厂;2500凝胶成像系统 上海天能科技有限公司;MyCycler Thermal Cycler PCR仪 美国Bio-Rad公司;7500 FastReal Time PCR instrument 美国ABI公司;RM2016型病理切片机 上海徕卡仪器有限公司;KD-P型组织摊片机 浙江省金华市科迪仪器设备有限公司;NIKON ECLIPSE E100型正置光学显微镜、NIKON DS-U3型成像系统 日本尼康株式会社。
1.2 实验方法
1.2.1 甜瓜预处理
清水冲洗甜瓜表面后使用2% H2O2水溶液浸泡30 s,清水冲洗3次。晾干贮于室内阴凉干燥处。
1.2.2 链格孢菌的侵染
损伤接种:甜瓜果实沿赤道穿刺打孔3次,每孔直径3.5 mm、深度5 mm,每孔等距(相距7 cm)。实验组:制备浓度1.0×106 spores/mL链格孢菌孢子悬浮液,每孔接20 μL;对照组:无菌水,每孔接20 μL。室温贮藏(25±5 ℃,空气相对湿度35%~40%)
1.2.3 取样方法
链格孢菌接种后,贮藏室温条件下,于0、2、3、6、8、10、12、14、16 d两种甜瓜各选择6个,实验组于接种处分离出含有果皮、果肉完整病斑组织,伽师瓜0~2 d未出现明显病斑,取样部位为损伤接种处;对照组于打孔接入无菌水部位取样,使用液氮速冻后,存放于-80 ℃超低温冰箱。
1.2.4 甜瓜果实病斑直径的测定
将甜瓜果实沿赤道部位接种点横向剖开,使用游标卡尺测量各病斑直径(包括过敏反应组织)[15],计算平均值,单位标识为cm。
1.2.5 组织切片显微结构观察
选择0 d(未侵染样品)与4、8、12、16 d同时含有果皮果肉的病斑组织,采用基底膜六胺银染色法(periodic acid-silver methenamine,PASM):甜瓜病斑组织石蜡切片分别使用入二甲苯Ⅰ、二甲苯Ⅱ浸泡20 min,石蜡溶解后,分别是使用无水乙醇、75%乙醇浸泡5 min并用蒸馏水清洗;使用PASM染液A,将切片浸泡过夜;使用PASM染液B,将切片浸泡15 min;使用烘箱预热PASM染液C,预热至60 ℃后,滴加在甜瓜病斑组织切片上,56~59 ℃加盖孵育40 min;镜下观察切片染色效果,并使用蒸馏水冲洗切片;PASM染液F浸泡10 min并使用蒸馏水清洗;将甜瓜病斑组织切片先后使用无水乙醇、二甲苯浸泡5 min,直到切片透明清晰时,使用中性树胶对切片封片;显微镜观察,Pannoramic全自动数字玻片扫描系统采集图像;Caseviewe(rC.V 2.4)软件于150倍的放大倍数下分析图像。
1.2.6 采后甜瓜细胞壁降解酶活性的测定方法
1.2.6.1 粗酶液的制备
称取0.20 g果肉组织,加入2 mL预冷的醋酸钠缓冲液,冰浴研磨,转入离心管,于4 ℃,1200 r/min条件下离心30 min,上清液即为粗酶液,于4 ℃冰箱中保存备用。
1.2.6.2 果胶裂解酶测定方法
参照张蕊等[16]的方法。取2.0 mL 1%果胶溶液(pH6.8)于试管中,40 ℃水浴平衡5 min,加入待测酶液1 mL,40 ℃保温10 min。取上述反应混合物1 mL于9 mL的0.01 mol HCl中充分混合终止反应,在235 nm处测定吸光值。
酶活单位定义(U):在40 ℃,pH6.8条件下,每分钟水解1%果胶,使235 nm处吸光值增加0.01的酶量定义为1个果胶裂解酶(PL)酶活单位。
1.2.6.3 纤维素酶测定方法
纤维素酶测定方法参照曹建康[17]的方法,纤维素酶活性以每小时每克果蔬组织样品(鲜重)在37 ℃催化羧甲基纤维素水解形成还原糖的质量表示,即μg/(h·g)。
1.2.6.4 β-葡萄糖苷酶测定方法
β-葡萄糖苷酶测定方法参照曹建康[17]的方法,β-葡萄糖苷酶活性以每小时每克果蔬组织样品(鲜重)在37 ℃催化水杨苷水解生成葡萄糖的质量表示,即μg/(h·g)。
1.2.6.5 多聚半乳糖醛酸酶测定方法
多聚半乳糖醛酸酶测定方法参照曹建康[17]的方法,多聚半乳糖醛酸酶活性以每小时每克果蔬组织样品(鲜重)在37 ℃催化多聚半乳糖醛酸水解生成半乳糖醛酸的质量表示,即μg/(h·g)。
1.2.7 链格孢菌细胞壁降解酶基因表达量的测定方法
1.2.7.1 链格孢菌总RNA提取和cDNA合成
RNA提取步骤总结淡明等[18]与王学仁等[19]的方法后稍作修改:取1 mL Buffer Rlysis-F加入1.5 mL RNase-free的离心管中备用,取100 mg病斑组织果肉部分用液氮研磨成粉末,加到上述1.5 mL离心管中,立即震荡混匀,向裂解样品中加入200 μL氯仿,离心5 min(转速12000 r/min,4 ℃),提取上清液,加入1/3体积无水乙醇混匀,常温下(25 ℃)放置3 min,去除上清液,将离心管于室温倒置10 min,加入50 μL RNase free Water溶解沉淀。核酸蛋白定量仪检测RNA浓度,琼脂糖电泳检测RNA完整性。
确定提取的RNA质量合格后进行反转录(反转录体系:RNA 1 μg、5×All-In-One RT MasterMix 4 μL最后使用RNase-free Water定量至20 μL),7 ℃反应15 min,60 ℃反应10 min取出,95 ℃反应3 min,得到的cDNA并置于−80 ℃超低温冰箱保存进行后续实验。
1.2.7.2 质粒提取
使用LB培养基对菌液进行扩培;离心12 min(转速3000 r/min,室温)并去除上清液;向离心管内加入250 μL含RNase A的细胞悬浮液,充分震荡分散悬浮;加入350 μL细胞裂解液,上下颠倒离心管5次,静置5 min;加入350 μL中和缓冲液,上下颠倒离心管5次后,离心10 min(转速12000 r/min,室温);去除上清液后,转移至离心吸附柱内离心1 min(转速12000 r/min,室温),去除管中废液;向吸附柱内加入500 μL漂洗液后离心1 min(转速12000 r/min,室温);取出吸附柱,套入1.5 mL无菌离心管中,加入35 μL洗脱缓冲液,静置3 min后,离心2 min(转速12000 r/min,室温),收集质粒DNA并进行质检。
1.2.7.3 质粒标准品的标准曲线制作
根据标准品的浓度按照下述公式计算质粒的初始拷贝数:
copynumber(copies/μL)=6.02×1023(copies/mol)MW(g/mol)×concentration(g/μL) (1) MW(g/mol)=DNA片段长度(bp)×660daltons (2) DNA片段长度(bp)=克隆片段长度+载体长度(bp) (3) 挑选一例单克隆质粒DNA将其作为起始质粒模板,即为质粒标准品1号梯度,按1:10比例依次递减稀释至7号梯度,每个梯度3个重复,进行荧光定量PCR检测,根据标准品每个梯度的拷贝数的log值与测得CT值的线性关系,绘制标准曲线。
1.2.7.4 荧光定量PCR引物设计
链格孢菌四种细胞壁降解酶基因AaPL、AaCx、AaPG、AaCB的引物序列见表1,引物设计参照本课题组前期对链格孢菌PL(GenBank:OP004937)、Cx(GenBank:OP004934)、PG(GenBank:OP004936)、CB(GenBank:OP004933)的同源克隆测序结果。
表 1 荧光定量mRNA检测引物信息Table 1. Information of Fluorescent Quantitative mRNA Detection Primers引物名称 序列(5'→3') 引物大小 PL-F TCCCATGAACTTCTTCAGCA 120 bp PL-R GTCTTAAGG CCACCGTCAAA Cx-F CTCCACCAACGTTTCCAACT 138 bp Cx-R TTGGATGGTAGACCCTCAGC PG-F AGGGTCCCCTCATCTCTGTT 117 bp PG-R TCTTGCCACCGTTAGTACCC CB-F ACCATTACGCCAGTGACTCC 100 bp CB-R CCTCTGAAGCTGGAGGAATG 1.2.7.5 链格孢菌细胞壁降解酶基因荧光定量PCR
荧光定量PCR反应体系:cDNA 2 μL、BlasTaq 2×qPCR Master Mix 10 μL、上下游引物各0.5 μL使用RNase-free water定容至20 μL。荧光定量PCR反应条件:预变性95 ℃,3 min;变性95 ℃,15 s(40个循环);退火/延伸60 ℃,1 min(40个循环)。根据PCR反应的Ct值代入相应的标准曲线计算出目的基因的表达量。
1.3 数据处理
采用IBM SPSS Statistics 23.0进行正态分布检验;对基因表达量的显著性分析采用独立样本T检验;对酶活性的显著性分析采用ANOVA单因素方差分析和Duncan’s多重比较检验;对基因表达量及酶活性进行双变量相关性分析;采用Origin 2021软件进行绘图。结果以平均值±标准差表示,P<0.05时差异显著。
2. 结果与分析
2.1 链格孢菌侵染采后甜瓜果实病斑发生规律
如图1所示,接种链格孢菌后,随着侵染天数的增加,两种甜瓜病害逐渐恶化,发病部位的果皮与果肉组织变黑,果皮组织出现塌陷,果肉病斑组织边缘出现褐色水渍状组织。前人研究发现,在自然生长环境下被链格孢菌侵染发生黑斑病的甜瓜果皮表面有黑色、微凹陷的圆斑,果肉病理组织颜色呈褐色至黑色,易于健康组织分离[20],本研究所发现的甜瓜黑斑病发病规律及特点与其描述相符。
![]() 图 1 链格孢菌侵染甜瓜果实发病情况Figure 1. Disease status of muskmelon fruit infected by Alternaria alternata
图 1 链格孢菌侵染甜瓜果实发病情况Figure 1. Disease status of muskmelon fruit infected by Alternaria alternata86-1甜瓜于2 d出现病斑,伽师瓜于4 d出现病斑,86-1甜瓜病斑出现时间早于伽师瓜。从图2可以看出,接种链格孢菌后在贮藏期间两类甜瓜病斑直径呈不断扩大的趋势。相同天数下,除4 d以外86-1甜瓜果肉病斑直径均显著大于伽师瓜果肉病斑(P<0.05);果皮病斑直径在个别天数下(4、8、16 d)伽师瓜组大于86-1甜瓜组,且两种甜瓜的果肉病斑增长速度均大于果皮病斑增长速度。
![]() 图 2 链格孢菌侵染甜瓜果实病斑直径变化趋势注:不同字母表示在相同侵染天数下,该细胞壁降解酶在不同品种甜瓜的不同组织上存在显著性差异(P<0.05),图4同。Figure 2. Variation of the diameter of melon lesions infected by Alternaria alternate
图 2 链格孢菌侵染甜瓜果实病斑直径变化趋势注:不同字母表示在相同侵染天数下,该细胞壁降解酶在不同品种甜瓜的不同组织上存在显著性差异(P<0.05),图4同。Figure 2. Variation of the diameter of melon lesions infected by Alternaria alternate2.2 链格孢菌侵染采后甜瓜组织切片染色观察
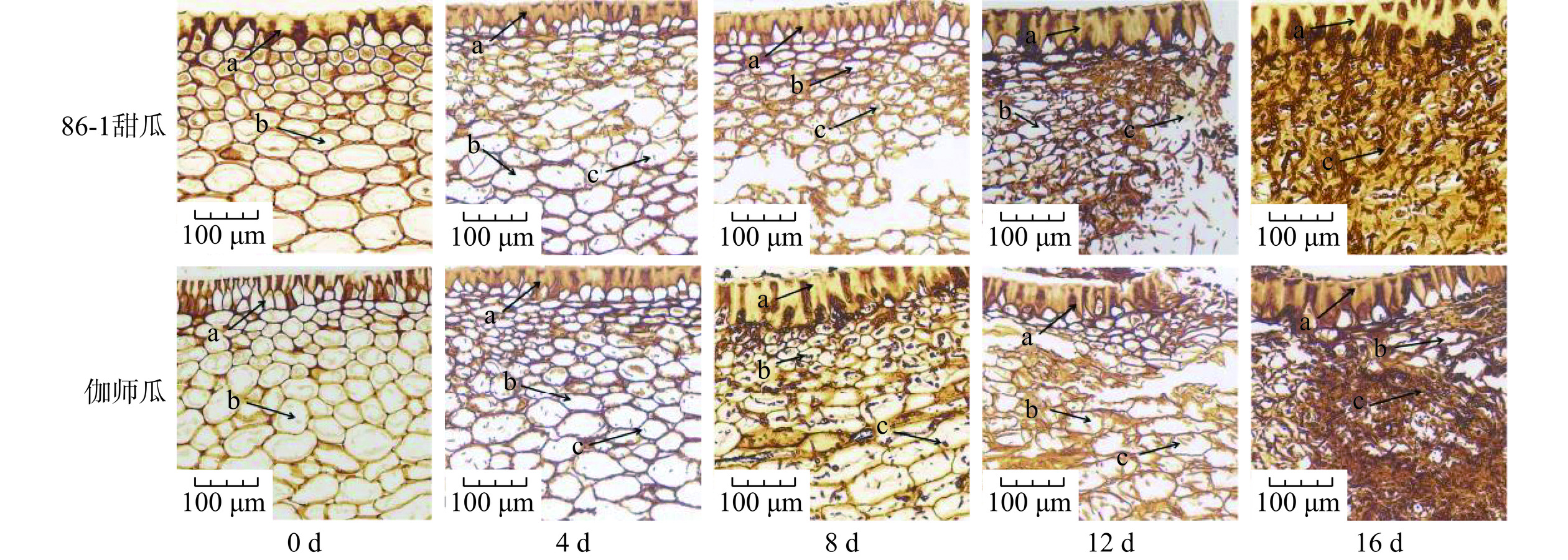
链格孢菌侵染采后甜瓜组织切片染色观察结果如图3所示。0 d时两种甜瓜细胞饱满、组织液充盈、结构完整、排列紧实;4 d时可以观察到两种甜瓜细胞内均有链格孢菌侵入,86-1甜瓜细胞内链格孢菌数目较多,能够发现部分菌丝体的存在,并有少部分果肉细胞破裂,而伽师瓜没有发现细胞破裂;链格孢菌侵染8 d,两种甜瓜细胞内链格孢菌数量较4 d时有明显增加,并且能够观察到有孢子产生,在86-1甜瓜中发现更多细胞破裂,而伽师瓜内仍然没有观察到细胞破裂的现象;12 d时,链格孢菌在两种甜瓜果肉组织内大量繁殖,其中86-1甜瓜的果皮细胞内可以发现链格孢菌,果肉细胞大量破碎,伽师瓜果肉细胞开始破裂;16 d时86-1甜瓜的果皮、果肉细胞完全解体,无法分辨细胞结构,果皮角质层下链格孢菌密集繁殖,伽师瓜仅有极少部分角质层下的果皮细胞形态完整,果肉细胞完全破碎。
![]() 图 3 链格孢菌侵染甜瓜果实组织切片PASM染色观察(150×)注:a:甜瓜果皮组织;b:甜瓜果肉细胞;c:链格孢菌(菌丝体或孢子)。Figure 3. PASM staining observation of Muskmelon fruit slices infected by Alternaria alternata (150×)
图 3 链格孢菌侵染甜瓜果实组织切片PASM染色观察(150×)注:a:甜瓜果皮组织;b:甜瓜果肉细胞;c:链格孢菌(菌丝体或孢子)。Figure 3. PASM staining observation of Muskmelon fruit slices infected by Alternaria alternata (150×)植物病原真菌侵染植物的之后,植物组织中与之最先接触的是植物细胞壁[21]。病原真菌可突破细胞壁,产生细胞壁降解酶降解植物细胞初生细胞壁和次生细胞壁中的多糖类物质,成功侵入并从中摄取营养[22−23]。从4 d的显微结构观察可以看出,两种甜瓜的细胞内部均有链格孢菌的存在,说明链格孢菌已突破细胞壁;16 d时两种甜瓜的果肉细胞已无法辨别细胞结构,链格孢菌密集繁殖,说明链格孢菌在后期已将甜瓜细胞内的营养物质消耗殆尽,开始通过细胞壁降解酶将甜瓜细胞壁中纤维素等物质水解至还原糖为自身提供能量,这与Tavares等[24]研究发现病原真菌可以直接通过水解植物细胞壁汲取营养的观点一致。
2.3 链格孢菌侵染采后甜瓜过程中细胞壁降解酶活性变化规律
如图4所示,随着贮藏天数的增加,四种细胞壁降解酶在两种甜瓜的健康组织中活性均呈现逐渐上升的趋势,其活性在病斑组织中显著高于健康组织(P<0.05)(2 d时Cx在伽师瓜病斑组织中活性高于健康组织但不显著)。两个品种甜瓜的实验组中,Cx和PL的活性变化都呈现上升-下降-上升-下降的趋势,CB和PG的活性变化分别在6~8 d、8~10 d较前两个时间点的斜率有明显下降。
![]() 图 4 链格孢菌侵染采后甜瓜过程中果胶裂解酶(A)、纤维素酶(B)、β-葡萄糖苷酶(C)多聚半乳糖醛酸酶(D)的活性变化规律Figure 4. Activity changes of pectin lyase (A), cellulase (B) and β-glucosidase (C) polygalacturonase (D) in postharvest muskmelon infected by Alternaria alternata
图 4 链格孢菌侵染采后甜瓜过程中果胶裂解酶(A)、纤维素酶(B)、β-葡萄糖苷酶(C)多聚半乳糖醛酸酶(D)的活性变化规律Figure 4. Activity changes of pectin lyase (A), cellulase (B) and β-glucosidase (C) polygalacturonase (D) in postharvest muskmelon infected by Alternaria alternata如图4A所示,伽师瓜病斑组织中,PL活性在6、14 d时达到峰值,分别为0 d时的5.46倍和8.02倍;86-1甜瓜病斑组织中,PL活性在4、10 d时达到峰值,分别为2 d时的4.68倍和7.55倍。2、4、8、10 d时,PL在86-1甜瓜中的活性均显著高于伽师瓜接种组(P<0.05);6、12、14、16 d时PL在伽师瓜接种组的活性显著高于86-1甜瓜接种组(P<0.05);其中86-1接种组PL活性最高峰值(10 d)是伽师瓜接种组最高峰值(14 d)的1.09倍。PL在伽师瓜接种组的活性第一次达到峰值的天数比86-1甜瓜接种组推后2 d,第二次达到峰值的天数比86-1甜瓜接种组推后4 d。
如图4B所示,伽师瓜病斑组织中,Cx在6、12 d时达到峰值,分别为0 d时的17.90倍和33.99倍;86-1甜瓜病斑组织中,Cx活性在4、12 d时达到峰值,分别为0 d时的22.98倍和50.56倍。除第6、14 d外,在相同天数下Cx在86-1甜瓜中的活性均高于伽师瓜接种组,且差异显著(P<0.05);其中86-1接种组Cx活性最高峰值(12 d)是伽师瓜接种组最高峰值(12 d)的1.27倍。Cx在伽师瓜接种组的活性两次达到峰值的天数均比86-1甜瓜接种组推后2 d。
如图4C所示,伽师瓜病斑组织中,CB活性在16 d时达到最大值,为0 d时的48.33倍;86-1甜瓜病斑组织中,CB活性在12 d达到峰值,为0 d时的39.31倍。除第14、16 d外,在相同天数下CB在86-1甜瓜中的活性均显著高于伽师瓜接种组(P<0.05);其中86-1接种组CB活性最高峰值(10 d)是伽师瓜接种组最大值(16 d)的1.10倍。
如图4D所示,伽师瓜病斑组织中,PG活性在12 d时达到峰值,是0 d时的20.38倍;86-1甜瓜病斑组织中,PG活性在12 d达到峰值,是0 d时的20.07倍,相同天数下PG在86-1甜瓜中的活性均显著高于伽师瓜接种组(P<0.05),12 d时同样高于伽师瓜接种组但差异不显著;14~16 d时PG在86-1甜瓜中的活性均高于伽师瓜接种组,且在16 d存在显著性差异(P<0.05)。其中86-1接种组PG活性峰值(12 d)是伽师瓜接种组峰值(12 d)的1.03倍。
2.4 链格孢菌侵染采后甜瓜过程中细胞壁降解酶基因的变化
如图5所示,链格孢菌侵染两个不同品种的甜瓜,除AaPG在伽师瓜接种组中、AaPL在86-1甜瓜接种组中、AaCB在86-1甜瓜接种组中表达量的变化趋势为升高-降低的趋势外,其余5组基因表达量的变化均呈现升高-降低-升高-降低的趋势。正常情况下,甜瓜果实内部病程相关蛋白及苯丙烷代谢相关酶的基因表达都处于较低的水平,但植物体在受到病原菌侵染后可以诱导病程相关蛋白[25]及苯丙烷代谢相关酶[14]的基因表达,通过病程相关蛋白裂解菌丝体,激活苯丙烷代谢途径释放黄酮等抗病物质抵御病原真菌的侵染[26−27]表现出抗病性。本研究中,细胞壁降解酶活性达到第一个峰值后的下降、酶活性增长速度减慢,以及基因表达量达到峰值后降低的现象,推测是由于甜瓜受到链格孢菌的侵染后激活相应的防御机制,通过上调自身病程相关蛋白、苯丙烷代谢酶的基因表达,加快病程相关蛋白、苯丙烷代谢酶的合成,从而抵御病原真菌的侵染。四种细胞壁降解酶的活性及基因表达量在后期的下降,则是由于甜瓜组织的营养成分被大量消耗,链格孢菌种内竞争导致了自身的消亡。
![]() 图 5 链格孢菌侵染过程中AaPL(A)、AaCx(B)、AaCB(C)、AaPG(D)的表达量变化注:*差异显著(P<0.05);**差异极显著(P<0.01)。Figure 5. Changes of AaPL (A), AaCx (B), AaCB (C), AaPG (D) expression during Alternaria alternata infection
图 5 链格孢菌侵染过程中AaPL(A)、AaCx(B)、AaCB(C)、AaPG(D)的表达量变化注:*差异显著(P<0.05);**差异极显著(P<0.01)。Figure 5. Changes of AaPL (A), AaCx (B), AaCB (C), AaPG (D) expression during Alternaria alternata infection如图5A所示,伽师瓜病斑组织中,AaPL表达量在6 d和12 d两次达到峰值;86-1病斑组织中,AaPL基因表达量在12 d达到峰值;其中86-1接种组AaPL基因表达量的峰值(12 d)是伽师瓜接种组峰值(12 d)的1.23倍。除14 d外,在相同侵染天数下,链格孢菌AaPL在86-1甜瓜接种组的表达量,均高于伽师瓜接种组;其中除6 d外,其他天数两个接种组的对比均存在显著或极显著的差异(P<0.05,P<0.01)。
如图5B所示,伽师瓜病斑组织中,AaCx表达量在6 d和12 d两次达到峰值;86-1病斑组织中AaCx表达量在4 d和12 d两次达到峰值;其中86-1接种组AaCx基因表达量的峰值(12 d)是伽师瓜接种组峰值(12 d)的1.27倍。除6~8 d外,在相同侵染天数下,链格孢菌AaCx在86-1甜瓜接种组的表达量均高于伽师瓜接种组,且相同天数下不同接种组AaCx表达量的对比均存在显著或极显著的差异(P<0.05,P<0.01)。AaCx在伽师瓜接种组中的表达量达到第一个高峰的时间比86-1接种组量推迟了2 d。
如图5C所示,伽师瓜病斑组织中,AaCB表达量在14 d达到峰值;86-1病斑组织中,AaCB表达量在10 d时AaCB的表达量达到峰值;其中86-1接种组AaCB基因表达量的峰值(10 d)是伽师瓜接种组峰值(14 d)的1.40倍。相同的侵染天数下,仅在侵染末期时(14、16 d)AaCB的在伽师瓜上的表达水平高于在86-1上的表达,其余天数AaCB在86-1甜瓜上的表达水平均高于在伽师瓜上的表达,且相同天数下不同接种组AaCB表达量的对比均存在显著或极显著的差异(P<0.05,P<0.01)。AaCB在伽师瓜接种组中的表达量最大峰值的出现时间较86-1甜瓜接种组推迟4 d。
如图5D所示,伽师瓜病斑组织中,AaPG表达量在12 d时达到峰值;86-1病斑组织中,在12 d达到峰值;其中86-1接种组AaPG基因表达量的峰值(12 d)是伽师瓜接种组峰值(12 d)的1.22倍。相同的侵染天数下,仅在侵染末期时(14、16 d)AaPG的在伽师瓜上的表达水平高于在86-1上的表达,其余天数AaPG在86-1甜瓜上的表达水平均高于在伽师瓜上的表达,且相同天数下不同接种组AaPG表达量的对比除6 d之外,均存在极显著的差异(P<0.01)。
玛尔哈巴等[28]将链格孢菌通过损伤接种的方式侵染86-1甜瓜和伽师瓜,通过比较病斑直径、病程相关蛋白活性,得出伽师瓜抗病能力高于86-1甜瓜的结论;而通常病原真菌的致病能力与寄主植物的抗病能力呈负相关的关系[29]。四种细胞壁降解酶的基因表达量在相同侵染天数下,多数天数在86-1甜瓜接种组中的表达量显著高于在伽师瓜接种组(P<0.05);同时,相同侵染天数下,多数天数86-1甜瓜接种组中的四种细胞壁降解酶的活性都显著高于伽师瓜接种组(P<0.05)。对比四种细胞壁降解酶在不同甜瓜接种组上基因表达量及酶活性出现峰值的时间,伽师瓜接种组上出现酶活性峰值及基因表达量峰值推迟的现象。结合前人研究,推测这与伽师瓜较高的抗病能力相关,同时也体现了链格孢菌在86-1甜瓜组中的侵染能力更强。
2.5 链格孢菌细胞壁降解酶活性与其基因表达量的相关性分析
通过SPSS软件对伽师瓜接种组及86-1甜瓜接种组的链格孢菌四种细胞壁降解酶活性与其基因表达进行相关性分析。由表2可知,链格孢菌整个侵染过程中,伽师瓜接种组的四种细胞壁降解酶活性与其基因表达量均呈正相关,其中PL酶活性与AaPL表达量没有显著相关性,其他三种酶的活性与其对应基因表达量相关性均为极显著(P<0.01)。由表3可知,在侵染过程中,86-1甜瓜接种组的四种细胞壁降解酶活性与其对应基因表达量均呈显著性正相关(P<0.05),其中Cx、CB、PG的酶活性和对应基因表达量相关性极显著(P<0.01)。
表 2 伽师瓜接种组链格孢菌细胞壁降解酶活性与其基因表达量的相关性分析Table 2. Correlation analysis of cell wall degrading enzyme activities and gene expression of Alternaria alternata in Jiashi melon inoculation group指标 Cx活性 PL活性 CB活性 PG活性 AaCx表达量 0.778** − − − AaPL表达量 − 0.617 − − AaCB表达量 − − 0.985** − AaPG表达量 − − − 0.945** 注:**表示差异极显著(P<0.01),*表示差异显著(P<0.05),表3同。 表 3 86-1甜瓜接种组链格孢菌细胞壁降解酶活性与其基因表达量的相关性分析Table 3. Correlation analysis of cell wall degrading enzyme activities and gene expression of Alternaria alternata in 86-1 melon inoculation group指标 Cx活性 PL活性 CB活性 PG活性 AaCx表达量 0.872** − AaPL表达量 − 0.715* AaCB表达量 0.930** AaPG表达量 0.678* 3. 结论
在链格孢菌侵染采后甜瓜过程中,细胞壁降解酶的活性及基因表达量变化趋势在6~10 d时有所波动,是受甜瓜自身抗病机制影响;总体来说四种细胞壁降解酶在86-1甜瓜中酶活性、基因表达量更高,结合对病斑组织的显微结构的观察,体现出链格孢菌对86-1甜瓜的侵染能力更强,同时也验证了伽师瓜较高的抗病性。本研究主要对链格孢菌侵染采后甜瓜过程中几种细胞壁降解酶的酶活性及基因表达量变化进行分析,还需要进一步发掘、研究链格孢菌其他致病关键酶,更深层次揭示链格孢菌的致病机理。
-
![]()
图 1 链格孢菌侵染甜瓜果实发病情况
Figure 1. Disease status of muskmelon fruit infected by Alternaria alternata
![]()
图 2 链格孢菌侵染甜瓜果实病斑直径变化趋势
注:不同字母表示在相同侵染天数下,该细胞壁降解酶在不同品种甜瓜的不同组织上存在显著性差异(P<0.05),图4同。
Figure 2. Variation of the diameter of melon lesions infected by Alternaria alternate
![]()
图 3 链格孢菌侵染甜瓜果实组织切片PASM染色观察(150×)
注:a:甜瓜果皮组织;b:甜瓜果肉细胞;c:链格孢菌(菌丝体或孢子)。
Figure 3. PASM staining observation of Muskmelon fruit slices infected by Alternaria alternata (150×)
![]()
图 4 链格孢菌侵染采后甜瓜过程中果胶裂解酶(A)、纤维素酶(B)、β-葡萄糖苷酶(C)多聚半乳糖醛酸酶(D)的活性变化规律
Figure 4. Activity changes of pectin lyase (A), cellulase (B) and β-glucosidase (C) polygalacturonase (D) in postharvest muskmelon infected by Alternaria alternata
![]()
图 5 链格孢菌侵染过程中AaPL(A)、AaCx(B)、AaCB(C)、AaPG(D)的表达量变化
注:*差异显著(P<0.05);**差异极显著(P<0.01)。
Figure 5. Changes of AaPL (A), AaCx (B), AaCB (C), AaPG (D) expression during Alternaria alternata infection
表 1 荧光定量mRNA检测引物信息
Table 1 Information of Fluorescent Quantitative mRNA Detection Primers
引物名称 序列(5'→3') 引物大小 PL-F TCCCATGAACTTCTTCAGCA 120 bp PL-R GTCTTAAGG CCACCGTCAAA Cx-F CTCCACCAACGTTTCCAACT 138 bp Cx-R TTGGATGGTAGACCCTCAGC PG-F AGGGTCCCCTCATCTCTGTT 117 bp PG-R TCTTGCCACCGTTAGTACCC CB-F ACCATTACGCCAGTGACTCC 100 bp CB-R CCTCTGAAGCTGGAGGAATG  下载: 导出CSV
下载: 导出CSV
表 2 伽师瓜接种组链格孢菌细胞壁降解酶活性与其基因表达量的相关性分析
Table 2 Correlation analysis of cell wall degrading enzyme activities and gene expression of Alternaria alternata in Jiashi melon inoculation group
指标 Cx活性 PL活性 CB活性 PG活性 AaCx表达量 0.778** − − − AaPL表达量 − 0.617 − − AaCB表达量 − − 0.985** − AaPG表达量 − − − 0.945** 注:**表示差异极显著(P<0.01),*表示差异显著(P<0.05),表3同。
下载: 导出CSV
表 3 86-1甜瓜接种组链格孢菌细胞壁降解酶活性与其基因表达量的相关性分析
Table 3 Correlation analysis of cell wall degrading enzyme activities and gene expression of Alternaria alternata in 86-1 melon inoculation group
指标 Cx活性 PL活性 CB活性 PG活性 AaCx表达量 0.872** − AaPL表达量 − 0.715* AaCB表达量 0.930** AaPG表达量 0.678*
下载: 导出CSV
-
[1] NUANGMEK W , AIDUANG W , SUWANNARACH N , et al. First report of fruit rot on cantaloupe caused by Fusarium equiseti in Thailand[J]. Journal of General Plant Pathology, 2019, 85(4):24-29.
[2] 郑贺云, 朱丽霞, 姚军, 等. 高通量测序法分析研究贮藏期甜瓜果实中真菌群落结构组成及动态变化[J]. 食品科学,2023,44(21):267−273. [ZHENG H Y, ZHU L X, YAO J, et al. Dynamic changes in fungal community structure of melon during storage[J]. Food Science,2023,44(21):267−273.] doi: 10.7506/spkx1002-6630-20230202-021 ZHENG H Y, ZHU L X, YAO J, et al. Dynamic changes in fungal community structure of melon during storage[J]. Food Science, 2023, 44(21): 267−273. doi: 10.7506/spkx1002-6630-20230202-021
[3] HUANG K, ZOU Y, LUO J, et al. Combining UV-C treatment with biocontrol yeast to control postharvest decay of melon[J]. Environmental Science and Pollution Research International, 2015, 22(18):7-13.
[4] PRENDES L P, MERIN M G, ZACHETTI V G L, et al. Impact of antagonistic yeasts from wine grapes on growth and mycotoxin production by Alternaria alternata[J]. Journal of Applied Microbiology,2021,131(2):72−79.
[5] 郑鄢燕, 赵力卉, 王宇滨, 等. 链格孢粉红单端孢通过降解果皮细胞壁导致哈密瓜病害[J]. 食品与发酵工业,2020,46(10):124−131. [ZHENG Y Y, ZHAO L H, WANG Y B, et al. Alternaria alternata and Trichothecium roseum cause disease of Hami melon by degrading the cell wall in the pericarp[J]. Food and Fermentation Industries,2020,46(10):124−131.] ZHENG Y Y, ZHAO L H, WANG Y B, et al. Alternaria alternata and Trichothecium roseum cause disease of Hami melon by degrading the cell wall in the pericarp[J]. Food and Fermentation Industries, 2020, 46(10): 124−131.
[6] 王砀砀, 赵聪聪, 蔡传江, 等. 厌氧真菌及其植物细胞壁降解酶应用研究进展[J]. 动物营养学报,2018,30(3):888−895. [WANG D D, ZHAO C C, CAI C J, et al. Research advances on application of anaerobic fungi and its plant cell wall degrading enzymes[J]. Chinese Journal of Animal Nutrition,2018,30(3):888−895.] doi: 10.3969/j.issn.1006-267x.2018.03.011 WANG D D, ZHAO C C, CAI C J, et al. Research advances on application of anaerobic fungi and its plant cell wall degrading enzymes[J]. Chinese Journal of Animal Nutrition, 2018, 30(3): 888−895. doi: 10.3969/j.issn.1006-267x.2018.03.011
[7] 安钢力. 实时荧光定量PCR技术的原理及其应用[J]. 中国现代教育装备,2018(21):19−21. [AN G L. The principle and application of real-time fluorescent quantitative PCR[J]. China Modern Educational Equipment,2018(21):19−21.] AN G L. The principle and application of real-time fluorescent quantitative PCR[J]. China Modern Educational Equipment, 2018(21): 19−21.
[8] 纪冬, 辛绍杰. 实时荧光定量PCR的发展和数据分析[J]. 生物技术通讯,2009,20(4):598−600. [JI D, XIN S J. Development and data analysis of real-time fluorescent quantitative PCR[J]. Letters in Biotechnology,2009,20(4):598−600.] doi: 10.3969/j.issn.1009-0002.2009.04.041 JI D, XIN S J. Development and data analysis of real-time fluorescent quantitative PCR[J]. Letters in Biotechnology, 2009, 20(4): 598−600. doi: 10.3969/j.issn.1009-0002.2009.04.041
[9] 郭杨, 陈世界, 郭万柱, 等. 荧光定量PCR技术及其应用研究进展[J]. 动物医学进展,2009,30(2):78−82. [GUO Y, CHEN S J, GUO W Z, et al. Advance in fluorescent quantitative PCR and its applications[J]. Progress in Veterinary Medicine,2009,30(2):78−82.] doi: 10.3969/j.issn.1007-5038.2009.02.021 GUO Y, CHEN S J, GUO W Z, et al. Advance in fluorescent quantitative PCR and its applications[J]. Progress in Veterinary Medicine, 2009, 30(2): 78−82. doi: 10.3969/j.issn.1007-5038.2009.02.021
[10] 曾禹睿慷, 白羽嘉, 王乾乾, 等. 链格孢菌侵染采后伽师瓜抗病关键酶活性变化规律[J]. 食品研究与开发,2022,43(16):8−17. [ZENG Y R K, BAI Y J, WANG Q Q, et al. Activities of key enzymes related to disease resistance in postharvest jiashi melon infected by Alternaria alternata[J]. Food Research and Development,2022,43(16):8−17.] doi: 10.12161/j.issn.1005-6521.2022.16.002 ZENG Y R K, BAI Y J, WANG Q Q, et al. Activities of key enzymes related to disease resistance in postharvest jiashi melon infected by Alternaria alternata[J]. Food Research and Development, 2022, 43(16): 8−17. doi: 10.12161/j.issn.1005-6521.2022.16.002
[11] 刘阳, 白羽嘉, 张郑啸, 等. 甜瓜抗链格孢侵染病程相关蛋白及苯丙烷代谢酶的变化[J]. 新疆农业科学,2023,60(7):1780−1789. [LIU Y, BAI Y J, ZHANG Z X, et al. Changes of related proteins and phenylpropane metabolic enzeymes in melon infected by Alternaria alternata[J]. Xinjiang Agricultural Sciences,2023,60(7):1780−1789.] doi: 10.6048/j.issn.1001-4330.2023.07.026 LIU Y, BAI Y J, ZHANG Z X, et al. Changes of related proteins and phenylpropane metabolic enzeymes in melon infected by Alternaria alternata[J]. Xinjiang Agricultural Sciences, 2023, 60(7): 1780−1789. doi: 10.6048/j.issn.1001-4330.2023.07.026
[12] 李梦, 白羽嘉, 王玉红, 等. 链格孢侵染对厚皮甜瓜抗菌物质及活性氧代谢相关酶活性的影响[J]. 食品工业科技,2017,38(23):245−249. [LI M, BAI Y J, WANG Y H, et al. Effects of Alternaria alternata infection on antimicrobial substances and activities of enzymes related to reactive oxygen metabolism in Muskmelon[J]. Science and Technology of Food Industry,2017,38(23):245−249.] LI M, BAI Y J, WANG Y H, et al. Effects of Alternaria alternata infection on antimicrobial substances and activities of enzymes related to reactive oxygen metabolism in Muskmelon[J]. Science and Technology of Food Industry, 2017, 38(23): 245−249.
[13] 王瑾, 白羽嘉, 冯作山, 等. 链格孢侵染采后甜瓜病程相关蛋白基因表达及酶活性变化规律[J]. 食品科学,2021,42(14):94−102. [WANG J, BAI Y J, FENG Z S, et al. Gene expression and activities of pathogenesis-related enzymes in postharvest muskmelons infected with Alternaria alternata[J]. Food Science,2021,42(14):94−102.] doi: 10.7506/spkx1002-6630-20200228-313 WANG J, BAI Y J, FENG Z S, et al. Gene expression and activities of pathogenesis-related enzymes in postharvest muskmelons infected with Alternaria alternata[J]. Food Science, 2021, 42(14): 94−102. doi: 10.7506/spkx1002-6630-20200228-313
[14] BAI Y, FENG Z, PAERHATI M, et al. Phenylpropanoid metabolism enzyme activities and gene expression in postharvest melons inoculated with Alternaria alternata[J]. Applied Biological Chemistry, 2021, 64(1):1−10.
[15] 白羽嘉, 张培岭, 黄伟, 等. 链格孢菌侵染采后甜瓜果实组织几丁质酶和β-1,3-葡聚糖酶基因表达分析[J]. 食品科学,2018,39(2):185−191. [BAI Y J, ZHANG P L, HUANG W, et al. Analysis of expression profiles of Chitinase and β-1,3-glucanase genes in muskmelon fruit tissue inoculated with Alternaria alternata[J]. Food Science,2018,39(2):185−191.] doi: 10.7506/spkx1002-6630-201802029 BAI Y J, ZHANG P L, HUANG W, et al. Analysis of expression profiles of Chitinase and β-1,3-glucanase genes in muskmelon fruit tissue inoculated with Alternaria alternata[J]. Food Science, 2018, 39(2): 185−191. doi: 10.7506/spkx1002-6630-201802029
[16] 张蕊, 陈慧洁, 章银军, 等. 黑曲霉果胶裂解酶和聚半乳糖醛酸酶合成的发酵调控[J]. 食品与发酵工业,2008,34(5):43−47. [ZHANG R, CHEN H J, ZHANG Y J, et al. Fermentation conditons of Aspergillus niger for production of pectin lyase and polygalacturonase[J]. Food and Fermentation Industries,2008,34(5):43−47.] ZHANG R, CHEN H J, ZHANG Y J, et al. Fermentation conditons of Aspergillus niger for production of pectin lyase and polygalacturonase[J]. Food and Fermentation Industries, 2008, 34(5): 43−47.
[17] 曹建康, 姜微波, 赵玉梅. 果蔬采后生理生化实指导验[M]. 北京:中国轻工业出版社, 2007:126−127. [CAO J K, JIANG W B, ZHAO Y M. Experiment guidance of postharvest physiology and biochemistry of fruits and vegetables[M]. Beijing:China Light Industry Press, 2007:126−127.] CAO J K, JIANG W B, ZHAO Y M. Experiment guidance of postharvest physiology and biochemistry of fruits and vegetables[M]. Beijing: China Light Industry Press, 2007: 126−127.
[18] 淡明, 黄海波, 郭安平, 等. 一种简单高效的真菌总RNA提取方法[J]. 福建热作科技,2006,31(4):19−20. [DAN M, HUANG H B, GUO A P, et al. A simple and efficient method for total RNA extraction from fungi[J]. Fujian Science & Technology of Tropical Crops,2006,31(4):19−20.] doi: 10.3969/j.issn.1006-2327.2006.04.009 DAN M, HUANG H B, GUO A P, et al. A simple and efficient method for total RNA extraction from fungi[J]. Fujian Science & Technology of Tropical Crops, 2006, 31(4): 19−20. doi: 10.3969/j.issn.1006-2327.2006.04.009
[19] 王学仁, 张改娜, 何涛, 等. 一种快速有效提取植物和真菌DNA和RNA的简易方法[J]. 基因组学与应用生物学,2009(6):1183−1188. [WANG X R, ZHANG G N, HE T, et al. A simple, rapid and effective method for extraction of DNA and RNA from fungi and plants[J]. Genomics and Applied Biology,2009(6):1183−1188.] WANG X R, ZHANG G N, HE T, et al. A simple, rapid and effective method for extraction of DNA and RNA from fungi and plants[J]. Genomics and Applied Biology, 2009(6): 1183−1188.
[20] 陈存坤. 新疆主栽厚皮甜瓜采后贮藏保鲜试验研究[D]. 天津:天津科技大学, 2009. [CHEN C K. Studies on post-harvest storage sinkiang Thick-skinned melon[D]. Tianjin:Tianjin University of Science and Technology, 2009.] CHEN C K. Studies on post-harvest storage sinkiang Thick-skinned melon[D]. Tianjin: Tianjin University of Science and Technology, 2009.
[21] 李沅泽, 李保华, 王彩霞. 苹果轮纹病菌产生细胞壁降解酶种类及其活性分析[J]. 青岛农业大学学报:自然科学版,2021,38(2):85−90,105. [LI Y Z, LI B H, WANG C X. The varieties and activities of cell wall-degrading enzymes produced by Botr yos phaeria dothidea from apple fruit[J]. Journal of Qingdao Agricultural University(Natural Science),2021,38(2):85−90,105.] LI Y Z, LI B H, WANG C X. The varieties and activities of cell wall-degrading enzymes produced by Botr yos phaeria dothidea from apple fruit[J]. Journal of Qingdao Agricultural University(Natural Science), 2021, 38(2): 85−90,105.
[22] LANGNER T G, HRE V. Fungal chitinases:function, regulation, and potential roles in plant/pathogen interactions[J]. Current Genetics,2015,62(2):243−254.
[23] SMIRNOVA O G, KOCHETOV A V. Plant cell wall and the mechanisms of resistance to pathogens[J]. Vavilov Journal of Genetics and Breeding,2016,19(6):715−723. doi: 10.18699/VJ15.109
[24] TAVARES M P, TÚLIO M, GOMES R F, et al. Secretomic insight into the biomass hydrolysis potential of the phytopathogenic fungus Chrysoporthe cubensis[J]. Journal of Proteomics,2021,236(62):104−121.
[25] 张培岭, 黄伟, 马玲, 等. 链格孢侵染对甜瓜病程相关蛋白活性及基因表达的影响[J]. 食品工业科技,2017,38(18):290−294,299. [ZHANG P L, HUANG W, MA L, et al. Effects of activities and gene expression of pathogenesis-related proteins in muskmelon fruit inoculated with Alternaria alternata[J]. Science and Technology of Food Industry,2017,38(18):290−294,299.] ZHANG P L, HUANG W, MA L, et al. Effects of activities and gene expression of pathogenesis-related proteins in muskmelon fruit inoculated with Alternaria alternata[J]. Science and Technology of Food Industry, 2017, 38(18): 290−294,299.
[26] RAMAROSON M L, HELESBEUX J J, HAMAMA L, et al. Deciphering the role of three specific flavonoids produced by carrot as potential breeding markers for resistance against Alternaria dauci[J]. Acta Horticulturae, 2023, 1(18):72-79.
[27] RADADIYA N, ANTALA V, DESAI H, et al. Expression of the pathogenesis related proteins during sesame-Macrophomina phaseolina interaction[J]. International Journal of Chemical Studies, 2020, 8:2698−2703.
[28] 玛尔哈巴·帕尔哈提, 白羽嘉, 王瑾, 等. 采后伽师瓜和86-1甜瓜果实抗链格孢菌侵染研究[J]. 食品与机械,2018,34(12):135−140. [MAERHABA P, BAI Y J, WANG J, et al. Infection of Alternaria alternata in postharvest Jiashi melon and 86-1 melon[J]. Food & Machinery,2018,34(12):135−140.] MAERHABA P, BAI Y J, WANG J, et al. Infection of Alternaria alternata in postharvest Jiashi melon and 86-1 melon[J]. Food & Machinery, 2018, 34(12): 135−140.
[29] CAGA B, MACHÁ R. Different pathogenicity of ergot isolates (Claviceps purpurea [Fr.] Tul.) on kentucky bluegrass (Poa pratensis L.)[J]. Plant Protection Science,2018,38(No.1):18−22.
-
期刊类型引用(0)
其他类型引用(3)
 下载:
下载:









计量
- 文章访问数: 64
- HTML全文浏览量: 14
- PDF下载量: 11
- 被引次数: 3



