


Recombinant Expression of Dextransucrase in Food-grade Lactic Acid Bacteria
-
摘要: 为研究葡聚糖在乳酸菌中的合成情况以及结构分析,本研究将葡聚糖蔗糖酶DsrB与带有SP45分泌信号肽的乳酸乳球菌表达载体pNZ8148质粒载体进行连接,电转入乳酸乳球菌NZ9000中进行异源表达。经由一系列醇沉、三氯乙酸去除蛋白质、二次醇沉、透析以及纯化等步骤得到多糖溶液。采用高效液相色谱法检测单糖组成;利用HPSEC测定葡聚糖的分子量;通过傅里叶红外技术、核磁共振技术以及扫描电镜分析多糖的结构并观察多糖表面特征。结果表明,将葡聚糖蔗糖酶在乳酸菌中进行表达,在蔗糖浓度为10%的条件下,葡聚糖产量为10.54 g/L;单糖组成显示仅含有葡萄糖一种单糖;胞外多糖分子量测定结果为2.4×106 Da;由傅里叶红外和核磁联合表明体外合成的胞外多糖仅含有α-(1,6)糖苷键;扫描电镜表示胞外多糖表面呈现为多孔状结构。在乳酸菌中异源表达,实现了葡聚糖的食品级生产,为葡聚糖于食品中应用提供了理论基础。Abstract: In order to study the synthesis of dextran in Lactobacillus and to analyze its structure, in this study, the dextransucrase DsrB was ligated with the Lactococcus lactis expression vector pNZ8148 plasmid vector with SP45 secretion signal peptide, and electrotransferred into L. lactis NZ9000 for heterologous expression. The polysaccharide solution was obtained through a series of steps including alcohol precipitation, protein removal by trichloroacetic acid, secondary alcohol precipitation, dialysis and purification. The composition of monosaccharides was detected by high performance liquid chromatography (HPLC). The molecular weight of dextran was determined by high-performance size exclusion chromatography (HPSEC). The structure of polysaccharides was analyzed by fourier transform infrared (FT-IR), nuclear magnetic resonance (NMR) and scanning electron microscopy (SEM), and the surface characteristics of polysaccharides were observed. The results showed that dextransucrase was expressed in lactic acid bacteria, and the glucan yield was 10.54 g/L at a sucrose concentration of 10%. The monosaccharide composition showed that it contained only one monosaccharide, glucose. The molecular weight of the extracellular polysaccharide was determined to be 2.4×106 Da. The combination of FT-IR and NMR indicated that the extracellular polysaccharide synthesized in vitro contained only the α-(1,6) glycosidic bond, and SEM indicated that the surface of the extracellular polysaccharide showed a porous structure. The heterologous expression in lactic acid bacteria realized the food-grade production of dextran and would provide a theoretical basis for the application of dextran in food.
-
葡聚糖,又称为右旋糖酐(Dextran),是一类具有支链葡萄糖的高聚物,主链由α-(1→6)糖苷键连接,支链由不同比例的α-(1→2)、α-(1→3)和α-(1→4)糖苷键连接形成[1]。该类多糖因含有较高比例的α-(1→6)糖苷键而具有良好的水溶性,得到了广泛的应用。
葡聚糖蔗糖酶(Dextransucrase)属于GH70家族酶,主要由明串珠菌属和链球菌属等产生的,是一种胞外酶,依附于细胞壁上或者游离于培养基中[2]。在传统意义上,GH70家族的酶仅被定义为葡聚糖蔗糖酶(DS),也被称为葡萄糖基转移酶(Gtfs)[3],它们可以裂解蔗糖并聚合蔗糖衍生的D-葡萄糖部分合成具有不同大小、连接方式和支化程度的α-葡聚糖[4]。还有一些GH70亚家族酶,例如4,6-α-葡聚糖转移酶(4,6-α-GTase)和4,3-α-葡聚糖转移酶(4,3-α-GTase)[5]。
目前葡聚糖的生产利用的是在大肠杆菌体系中葡聚糖蔗糖酶催化进行体外合成或者是由菌株发酵生成[6],而未有利用乳酸菌体系来生产葡聚糖的。由葡聚糖蔗糖酶催化合成或由菌株生成的葡聚糖,可以在不同方面有所应用。比如高分子量葡聚糖可以应用于食品行业中当做乳化剂和增稠剂使用,还可以应用于化工行业中作为分子筛基质等[7]。本研究的目的是利用乳酸菌体系催化合成高分子量葡聚糖,实现葡聚糖的食品级生产,更好的促进葡聚糖的应用。
本实验将来自柠檬明串珠菌的葡聚糖蔗糖酶基因DsrB在乳酸乳球菌NZ9000中分泌表达,以蔗糖作为底物进行体外催化合成葡聚糖,以乳酸乳球菌作为宿主,以期实现葡聚糖的食品级生产,对乳酸菌中合成的葡聚糖的结构进行表征,为葡聚糖的食品级宿主生产提供理论支持。
1. 材料与方法
1.1 材料与仪器
柠檬明串珠菌中葡聚糖蔗糖酶DsrB基因、乳酸乳球菌NZ9000、大肠杆菌MC1061、带有SP45分泌信号肽的pNZ8148质粒载体(前期构建) 均为本实验室保存(均保存于−80 ℃冰箱);Taq DNA高保真酶、DNA产物纯化试剂盒、琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒 北京天根生化技术有限公司;限制性核酸内切酶:PstI、XbaI 宝生物工程(大连)有限公司;Nisin(乳酸链球菌肽)、葡聚糖系列分子量标准品(T1000(1×106 Da)、T500(5×105 Da)、T300(3×105 Da)、T100(1×105 Da)、T70(7×104 Da)、 T10(1×104 Da))、MRS培养基、LB培养基 分析纯,北京索莱宝生物科技有限公司;M17培养基 分析纯,山东青岛奥博星生物技术有限公司;GM17培养基:以M17培养基为基础,加入2%葡萄糖;SGM17培养基:在GM17肉汤培养基的基础上加入10%的蔗糖。
DH 3600型恒温培养箱 天津市泰斯特仪器有限公司;TGL16M型台式高速低温冷冻离心机 长沙易达仪器;S1000型PCR仪 上海伯乐生命医学产品有限公司;Tanon EPS 300型电泳仪、Tanon-4600SF型紫外凝胶成像仪 上海天能生物科技有限公司;ZR2016-0690型高压蒸汽灭菌器 重庆中容石化机械制造有限公司;QL-861型微型涡旋混合仪 海门市麒麟医用仪器厂;SW-CJ-IF型超净台 苏净集团苏州安泰空气技术有限公司;HC-0120-08型凝胶柱 北京瑞达恒辉科技发展有限公司;HL-2型恒流泵 上海青浦沪西仪器厂;FD5-Series型冷冻干燥机 美国SIM冻干机公司;Agilent Technologies 1200 Series型高效液相色谱仪 中国安捷伦科技有限公司;N5000型紫外可见分光光度计 上海佑科仪器仪表有限公司;HJ-3型数显恒温磁力搅拌器 金坛市杰瑞尔电器有限公司;S-4800扫描电子显微镜 日本Hitachi。
1.2 实验方法
1.2.1 重组菌载体构建
1.2.1.1 引物设计
引物是根据前期确定葡聚糖蔗糖酶DsrB基因信息时进行设计的。
葡聚糖蔗糖酶全长基因的扩增:使用可以在乳酸菌中分泌表达的含有分泌信号肽的pNZ8148质粒作为表达载体,引物名称及序列如下:
正向引物:8148DsrBF1:CTGCAGGATAGCACAAACACAG
反向引物:8148DsrBR1:TCTAGACTTTAGCAAGCCGTTG
1.2.1.2 PCR扩增
体系(50 μL):2×buffer添加量为25 μL,dNTP、pfu、8148DsrBF1、8148DsrBR1和基因组DNA的添加量都是1 μL,ddH2O的添加量是20 μL。扩增条件:95 ℃预变性5 min,95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,72 ℃循环30次,72 ℃保持5 min,4 ℃保存。待PCR反应结束,取5 μL PCR扩增反应的产物进行琼脂糖凝胶电泳,剩余的PCR扩增产物使用TIANGEN DNA胶回收试剂盒进行胶回收,使用TIANGEN的DNA产物纯化试剂盒对其进行产物纯化。
1.2.1.3 质粒的提取
将含有质粒(pNZ8148-SP45加强)的大肠杆菌在含有氯霉素的LB固体培养基上进行三区划线,30 ℃培养12 h,挑取平板上的单克隆菌落,将其接种于含有氯霉素的5 mL LB液体培养基中,30 ℃培养12 h,在培养完成后,8000 r/min离心15 min收集菌体,使用TIANGEN的质粒小提试剂盒提取质粒,将其存放于−20 ℃备用。
1.2.1.4 质粒表达载体的构建
将提取到的质粒和葡聚糖蔗糖酶基因,在37 ℃的条件下进行2 h的双酶切反应,酶切体系:10×buffer添加量为5 μL、pNZ-8148-SP45加强质粒载体/目的片段添加量为30 μL、PstI和XbaI的添加量都是1 μL、ddH2O添加量为13 μL。酶切产物纯化:酶切后产物经琼脂糖凝胶电泳,纯化回收酶切片段。
将酶切回收纯化的产物,在25 ℃下连接2 h。连接体系:5×buffer添加量为2 μL、pNZ-8148-SP45加强质粒载体添加量为1 μL、目的片段添加量为6 μL、T4连接酶添加量为1 μL。
1.2.1.5 E. coli MC1061感受态细胞的转化
取储存于-80 ℃冰箱中的E. coli MC1061感受态细胞放置于冰上,将10 μL的连接产物加入到感受态细胞中(轻轻震动,使其混合均匀),冰浴30 min,42 ℃ 热激90 s,冰浴3 min;加入1 mL的LB液体培养基,37 ℃ 200 r/min振荡培养1 h。
涂板:将转化培养后的菌液,取100 μL均匀的涂在含有氯霉素的平板上,30 ℃培养12 h。单克隆验证:挑取单菌落,进行菌落验证PCR。菌落PCR的反应体系为:2×Mix添加量为10 μL、(AcmA-Term、8148-JCF和单菌落)的添加量均为1 μL、ddH2O的添加量均为7 μL。采用琼脂糖凝胶电泳进行验证。实验中的聚合酶链式反应(PCR(Polymerase Chain Reaction))产物的测序以及所需引物均由苏州金唯智生物科技有限公司完成。
重组质粒的提取:将单克隆菌落挑取至含有氯霉素的LB液体培养基中,30 ℃培养12 h,使用TIANGEN质粒小提试剂盒提取重组质粒。
1.2.2 乳酸乳球菌NZ9000电转化
将构建成功的目标重组质粒采用电转化的方法转入到乳酸菌NZ9000中。提取pNZ8148-SP45加强-DsrB质粒,利用电穿孔仪进行电转化。将电转杯提前放入−20 ℃进行预冷。电转仪工作电压为1200 V,25 μF,200 Ω,将上述电转杯放入电转仪中,将质粒电转化至L. lactis NZ9000中,用冰浴的1 mL的乳酸乳球菌电转化复苏液将电转杯中细胞重悬,重悬液转移至1.5 mL离心管中,30 ℃振荡复苏2 h。经8000 r/min离心1 min收集菌体,留下约100 μL上清液重悬菌体,将菌液涂布于含10 μg/mL氯霉素的GM17固体平板上,同时将pNZ8148空载体电转化至L. lactis NZ9000中做为空白对照,于30 ℃培养箱中至长出单菌落,约12~24 h。挑取阳性克隆子进行菌落PCR鉴定,琼脂糖凝胶电泳检测片段大小正确即表明获得在乳酸菌中重组表达的pNZ8148-SP45加强-DsrB菌株。
1.2.3 重组酶DsrB诱导表达条件确定
将pNZ8148-SP45加强-DsrB菌株在5 mL的GM17培养基中活化后,以2%的接种量转接入新鲜的SGM17(含有10%蔗糖)培养基中,在30 ℃条件下静置培养,培养至OD600为0.6时,分别设置诱导剂nisin的浓度为1、4、6、8、10、12、14和16 ng/mL[8−9],在30 ℃培养12 h,以不添加nisin的菌液作为空白对照。
以上述最佳nisin浓度作为nisin终浓度来进行诱导时间的优化。诱导时间设置为10、12、16、20、24、28和32 h[10−11]。
在进行完nisin浓度和诱导时间之后,以葡聚糖产量作为判定指标,来确定最佳诱导条件。
1.2.4 重组酶DsrB合成葡聚糖的纯化以及纯度鉴定
在培养完成后,以4 ℃、8000 r/min离心15 min去除菌体沉淀,用预冷的95%乙醇进行醇沉,再用10%三氯乙酸进行除蛋白,之后再用预冷的95%乙醇进行二次醇沉,得到粗多糖溶液。用截留分子量为3500 Da的透析袋进行透析(4 ℃,48 h),之后用Sephadex G-100对透析后的溶液进行纯化[12],得到纯化后的多糖溶液。对纯化后的多糖溶液用紫外可见分光光度计进行全波长扫描,在波长为190~800 nm范围内来检测多糖的纯度。将纯化后的多糖样品进行冻干,去除水分得到固体的葡聚糖。
1.2.5 重组酶DsrB合成葡聚糖分子量测定
采用高效体积排阻色谱法(HPSEC)对纯化后的胞外多糖样品进行纯度检测以及分子量测定。
HPSEC检测条件:流动相为去离子水;流速为0.8 mL/min;检测器采用的为蒸发光检测器(1260 Infinity ELSD);进样体积为20 μL;色谱柱采用的是Shodex OH-pak SB-804;柱温为25 ℃。
分子量标准曲线的绘制:将葡聚糖系列分子量标准品(T1000、T500、T300、T100、T70和T10)分别溶于水,配制成浓度为10 mg/mL的标准品溶液,用孔径为0.22 μm的膜进行过滤,装入1.5 mL的液相瓶中,开始进样。记录各个标准品的洗脱峰保留时间,以葡聚糖标准品分子量的对数值为纵坐标,洗脱时间为横坐标绘制标准曲线。回归方程为y=−0.5499x+9.8072,决定系数R²=0.9924。
纯化后的胞外多糖分子量测定:准确称取5 mg纯化后的胞外多糖样品溶于5 mL水中,制成浓度为1 mg/mL的溶液,用0.22 μm的膜过滤,装入液相瓶中进行上样。根据得出的洗脱时间代入上述回归方程计算得出胞外多糖的分子量。
1.2.6 重组酶DsrB合成葡聚糖结构表征
1.2.6.1 傅里叶红外光谱分析
将胞多糖粉末与烘干至恒重的溴化钾粉末按照1:100比例混合均匀,将混合物压制成1 mm的薄片,进行测定。使用BIO-RAD FTS3000红外光谱扫描仪获得光谱,扫描范围为400~4000 cm−1。
1.2.6.2 核磁共振波谱分析
通过核磁共振分析阐明多糖的结构,NMR光谱是由Bruker Avance NEO 400MHE光谱仪进行测定。称取5 mg冻干后的多糖样品,加入5 mL D2O充分溶解,制成浓度为1 mg/mL的样品溶液,真空冷冻干燥后,用D2O溶解,重复三次,转入核磁管中,测得一维1H-NMR和一维13C-NMR以及二维的1H-13C HSQC图谱。
1.2.6.3 单糖组成测定
采用高效液相色谱法测定纯化后多糖的单糖组成,内容包括多糖的前处理和液相色谱条件分析。
多糖前处理:取1 mL浓度为10 mg/mL的多糖样品于耐高温的消解管中,60 ℃用氮气进行吹干,加入5 mL的2 mol/L三氟乙酸溶液,用螺纹塞拧紧密封,在121 ℃下酸解2 h,等待冷却将其转移到旋蒸瓶中,用甲醇进行清洗瓶壁,在60 ℃下进行负压旋蒸,加入适量的甲醇旋转蒸干,重复进行两次,等待冷却后加入2 mL的去离子水进行超声复溶,过0.22 μm孔径的滤膜进行上机。
液相色谱条件分析:采用的是岛津LC-2030 3D Plus高效液相色谱系统。采用Waters Xbridge Amide(250 mm×4.6 mm,3.5 μm)色谱柱分析单糖组成。流动相为氨水溶液(1%)与乙腈,体积比为3:7,进样量为20 μL,柱温为25 ℃,流速为1.0 mL/min,示差检测器的型号为RID 20A,检测器的温度为40 ℃,等度洗脱。通过比较标准化合物与样品的保留时间,得到胞外多糖的成分。
1.2.6.4 扫描电镜观察胞外多糖的微观表面结构
取适量干燥后的多糖样品,将样品固定在扫描电镜样品台上,表面镀上一层导电金膜(约10 nm),采用扫描电子显微镜观察样品,加速电压为15 kV,放大倍数为1000倍。
1.3 数据处理
所有样品均测量三次,以计算平均值和标准偏差。数据使用SPSS 20.0版软件进行分析,采用单因素方差分析(ANOVA)。采用邓肯检验来确定处理间的显著差异(P<0.05)。实验结果用Origin 9.8制图。
2. 结果与分析
2.1 重组菌的构建
图1(a)是右旋糖苷蔗糖酶的载体构建示意图。从图中可以看出SP45分泌信号肽与葡聚糖蔗糖酶DsrB连接后连接至pNZ8148载体质粒上[13]。图1(b)是重组质粒电转入到乳酸乳球菌NZ9000中的单克隆菌落PCR验证图,目的基因的大小为4404 bp,由图可知重组载体构建成功,将带有重组质粒的大肠杆菌保存在−80 ℃冰箱中。
![]() 图 1 重组葡聚糖蔗糖酶载体构建示意图注:(a):载体构建示意图;(b):菌落验证图。Figure 1. Schematic diagram of recombinant dextransucrase vector construction
图 1 重组葡聚糖蔗糖酶载体构建示意图注:(a):载体构建示意图;(b):菌落验证图。Figure 1. Schematic diagram of recombinant dextransucrase vector construction2.2 重组酶DsrB诱导表达条件确定
在食品级表达体系NICE表达系统中,nisin可以诱导蛋白进行表达,但是高浓度的nisin也会抑制菌体的生长[14−16]。如图2(a)所示,在培养基中的nisin浓度低于10 ng/mL时,葡聚糖产量是较低的,随着nisin浓度的增加,葡聚糖产量呈现先上升后下降的趋势,在nisin浓度为10 ng/mL时达到最大。因此,确定最佳诱导剂浓度为10 ng/mL。同样,诱导时间对于目的蛋白的表达量影响巨大[17]。如图2(b)所示,随着诱导时间的增加,葡聚糖蔗糖酶的表达增加,进而葡聚糖的生成量在逐渐增加,在28 h时达到最大,之后趋于平稳。由此确定,葡聚糖蔗糖酶在乳酸菌中诱导表达的最佳条件为nisin浓度10 ng/mL,诱导时间28 h,在蔗糖浓度为10%的条件下,葡聚糖的最大产量为10.54 g/L。Feng等[12]采用柠檬明串珠菌B-2发酵,以75 g/L的蔗糖作为底物,生成的葡聚糖产量为28.3 g/L;Song等[18]采用柠檬明串珠菌SK24.002中的葡聚糖蔗糖酶催化蔗糖(100 mg/mL),生成46.4 mg/mL葡聚糖。虽然上述提取率较高,但使用遗传背景不清楚的野生型菌株发酵生成葡聚糖和含有内毒素大肠杆菌重组酶催化合成葡聚糖可能存在一定的安全性隐患[15],未实现葡聚糖的食品级生产,采用食品级表达宿主乳酸乳球菌(Generally Regarded As Safe,GRAS)来合成葡聚糖,合成的葡聚糖是安全的。
![]() 图 2 诱导表达条件确定注:(a)nisin浓度;(b):诱导时间。Figure 2. Determination of induced expression conditions
图 2 诱导表达条件确定注:(a)nisin浓度;(b):诱导时间。Figure 2. Determination of induced expression conditions2.3 重组酶DsrB合成葡聚糖纯度鉴定
采用凝胶柱层析法对粗多糖进行纯化[19],采用全波长扫描对多糖的纯度进行检测[20−21]。全波长扫描结果图如图3所示,最大吸收峰在200 nm左右,在260~280 nm范围内没有特征吸收峰,该结果表明纯化后的多糖不含有核酸和蛋白质。
2.4 重组酶DsrB合成葡聚糖的分子量
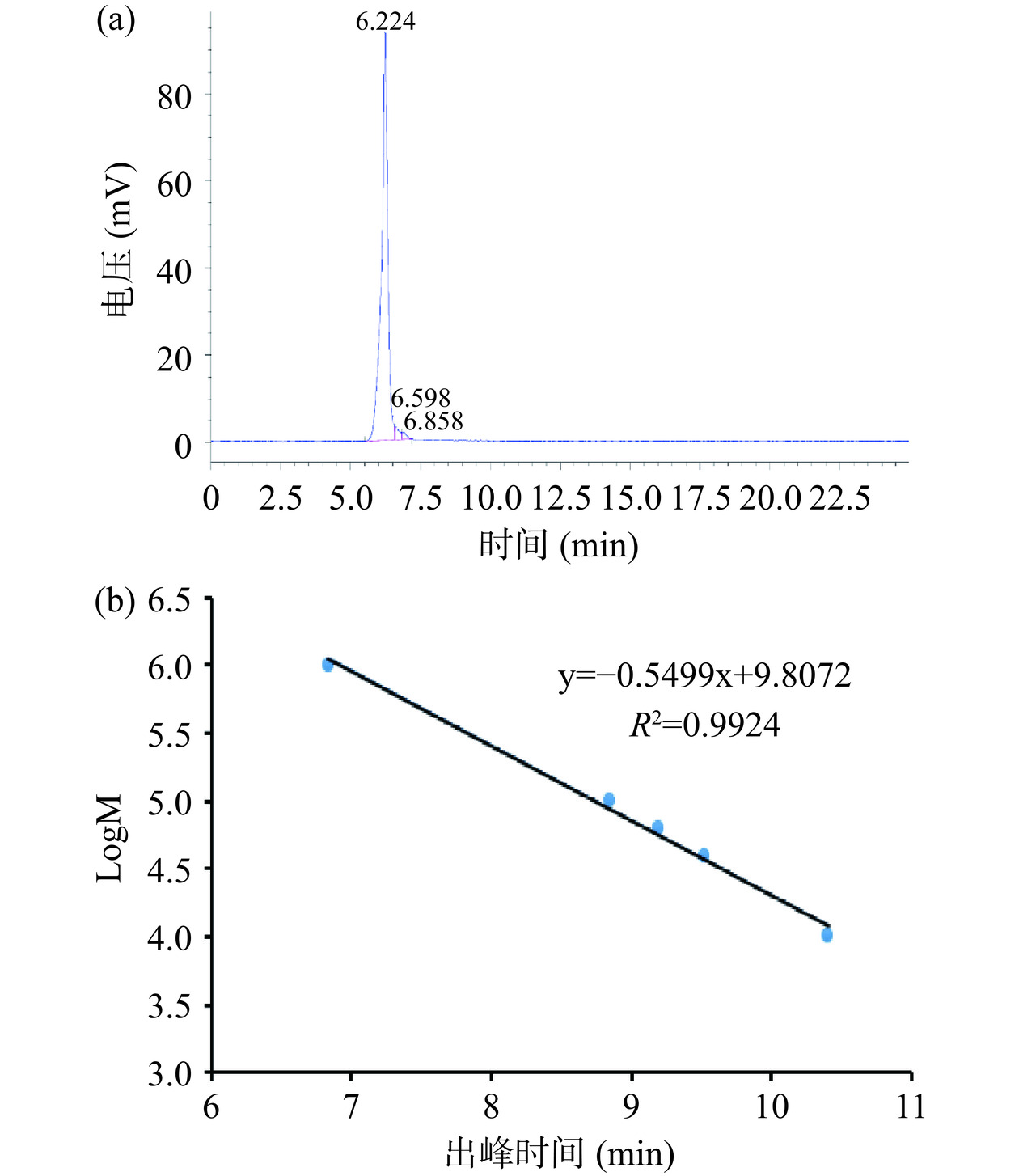
对纯化后的葡聚糖进行分子量测定,采用高效体积排阻色谱法(HPSEC)测定其分子量[12,22]。分子量测定的结果如图4所示,在图4(a)中含有一个独立对称的检出峰,出峰时间为6.224 min。图4(b)是葡聚糖的标准曲线,根据建立的标准曲线,将出峰时间代入其中得出在乳酸乳球菌中合成的葡聚糖的分子量为2.4×106 Da。
![]() 图 4 葡聚糖的分子量图谱注:(a):HPSEC图谱;(b):葡聚糖标准曲线。Figure 4. Molecular weight profile of dextran
图 4 葡聚糖的分子量图谱注:(a):HPSEC图谱;(b):葡聚糖标准曲线。Figure 4. Molecular weight profile of dextran2.5 重组酶DsrB合成葡聚糖的结构表征
在乳酸菌中生成的葡聚糖经纯化后的傅里叶红外光谱图如图5所示。在3418 cm−1的特征吸收峰是由O-H键拉伸振动引起的[23]。2927 cm−1处特征吸收峰是由C-H键拉伸振动引起的[24]。1645 cm−1处特征吸收峰是由C=O拉伸振动引起的。1017 cm−1位置的特征吸收峰表明该EPS含有α-(1,6)糖苷键。
![]() 图 5 乳酸菌中合成葡聚糖的傅里叶红外光谱图Figure 5. FT-IR map of dextran production in lactic acid bacteria
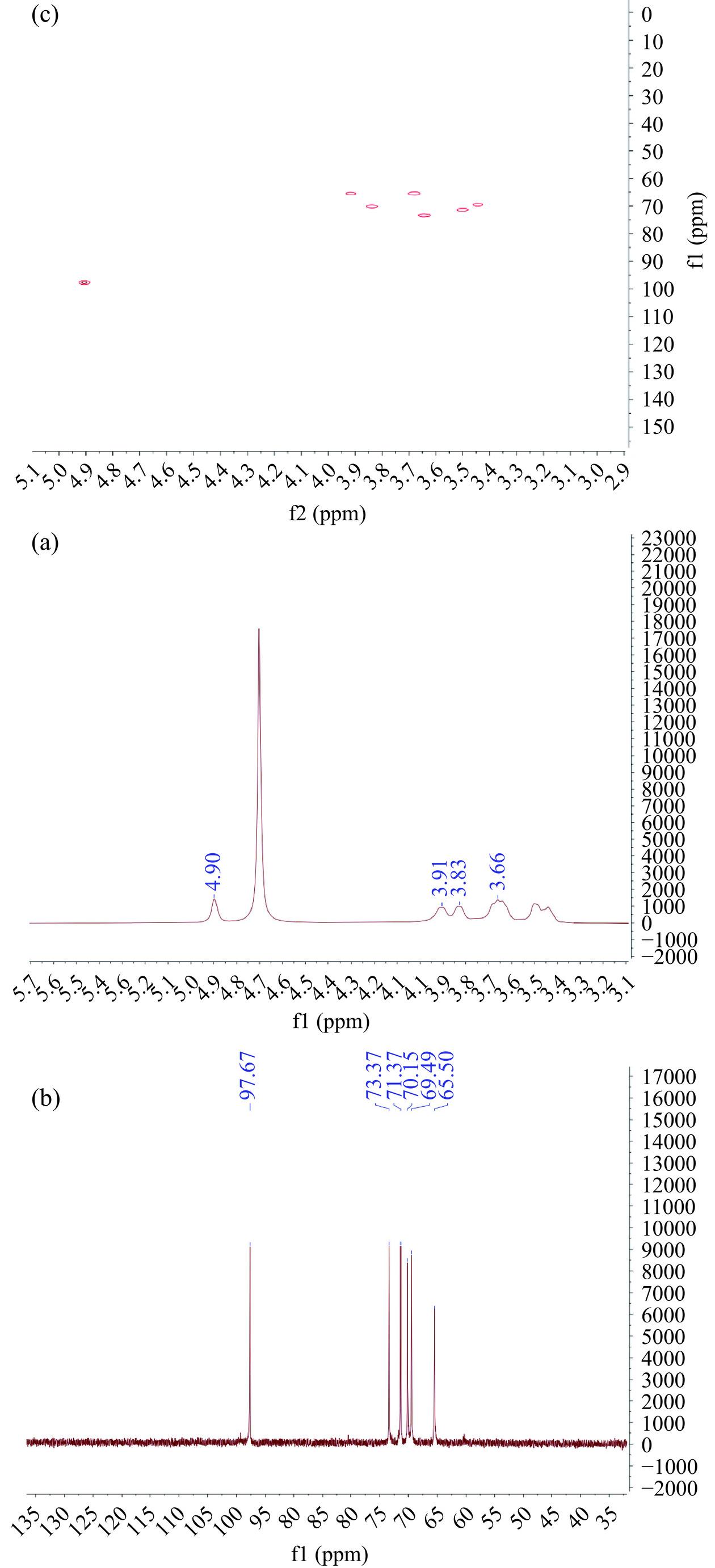
图 5 乳酸菌中合成葡聚糖的傅里叶红外光谱图Figure 5. FT-IR map of dextran production in lactic acid bacteria图6(a)中的异头质子区域仅有一个高强度信号峰,位于4.90 ppm处[25],此结果与傅立叶红外光谱中916 cm−1处的特征吸收峰相对应。在图6(b)中得出,异头碳区只有一个高强度信号峰,位于97.67 ppm处[23],代表α-(1,6)糖苷键,该位置的峰与1H图谱中4.90 ppm处的特征吸收峰是互相对应[26]。核磁1H图谱和13C图谱的结果联合证明了该多糖样品只含有一个α-(1,6)糖苷键[27]。图6(c)是纯化后葡聚糖的HSQC图谱,该多糖样品显示了六个信号,证实了多糖是以α-(1,6)糖苷键组成的直链葡聚糖[28]。在异头碳区域仅有一个信号峰,即13C图谱中97.67 ppm处的C1信号与1H图谱中4.90 ppm处的质子信号的交叉峰[29−31],证明该葡聚糖的结构单元中只存在一种多糖。
![]() 图 6 乳酸菌中合成葡聚糖的核磁共振图注:(a):1H图谱;(b):13C图谱;(c):HSQC图谱。Figure 6. NMR image of dextran production in lactic acid bacteria
图 6 乳酸菌中合成葡聚糖的核磁共振图注:(a):1H图谱;(b):13C图谱;(c):HSQC图谱。Figure 6. NMR image of dextran production in lactic acid bacteria采用高效液相色谱法对生成的葡聚糖进行单糖组成的测定[18,32]。单糖组成的结果如图7所示,由图7(a)可以看出,在7.050 min处有一个检出峰,与图7(b)的标准品液相图谱对比得出,此位置的检出峰代表的是葡萄糖,表明该多糖样品中只含有葡萄糖一种单糖。
![]() 图 7 乳酸菌中合成葡聚糖的单糖组成图注:(a):葡聚糖高效液相图谱;(b):标准品高效液相图谱。Figure 7. Dextran-producing monosaccharide composition in lactic acid bacteria
图 7 乳酸菌中合成葡聚糖的单糖组成图注:(a):葡聚糖高效液相图谱;(b):标准品高效液相图谱。Figure 7. Dextran-producing monosaccharide composition in lactic acid bacteria在乳酸菌中生成经纯化后的葡聚糖的扫描电镜图谱如图8所示。图8中扫描电镜的放大倍数为1000倍。从图中可以看出,葡聚糖表面光滑、有少量的孔洞存在,这种孔洞结构可使其具有高度的亲水性,可作为胶凝剂或增稠剂应用于食品和化妆品行业等[33−34]。
![]() 图 8 乳酸菌中合成葡聚糖的扫描电镜结果(1000×)Figure 8. Scanning electron microscopy of dextran production in lactic acid bacteria (1000×)
图 8 乳酸菌中合成葡聚糖的扫描电镜结果(1000×)Figure 8. Scanning electron microscopy of dextran production in lactic acid bacteria (1000×)3. 结论
本研究将柠檬明串珠菌DsrB基因在乳酸乳球菌NZ9000中进行重组表达,并且对生成的葡聚糖的结构进行测定。结果表明,在蔗糖浓度为10%时,在乳酸菌中葡聚糖产量为10.54 g/L;在乳酸菌中合成的葡聚糖仅含有葡萄糖一种单糖;在乳酸菌中合成的葡聚糖分子量为2.4×106 Da;该胞外多糖仅含有α-(1,6)糖苷键,不含有支链;其表面特征为孔状结构,此结构在化工领域有很大的应用前景。本研究在食品级表达体系乳酸乳球菌中合成葡聚糖,为葡聚糖的食品级生产提供了理论支持以及实验数据支持,同时也为葡聚糖的合成体系提供了一种新的思路。
-
![]()
图 1 重组葡聚糖蔗糖酶载体构建示意图
注:(a):载体构建示意图;(b):菌落验证图。
Figure 1. Schematic diagram of recombinant dextransucrase vector construction
![]()
图 2 诱导表达条件确定
注:(a)nisin浓度;(b):诱导时间。
Figure 2. Determination of induced expression conditions
![]()
图 4 葡聚糖的分子量图谱
注:(a):HPSEC图谱;(b):葡聚糖标准曲线。
Figure 4. Molecular weight profile of dextran
![]()
图 5 乳酸菌中合成葡聚糖的傅里叶红外光谱图
Figure 5. FT-IR map of dextran production in lactic acid bacteria
![]()
图 6 乳酸菌中合成葡聚糖的核磁共振图
注:(a):1H图谱;(b):13C图谱;(c):HSQC图谱。
Figure 6. NMR image of dextran production in lactic acid bacteria
![]()
图 7 乳酸菌中合成葡聚糖的单糖组成图
注:(a):葡聚糖高效液相图谱;(b):标准品高效液相图谱。
Figure 7. Dextran-producing monosaccharide composition in lactic acid bacteria
-
[1] MOLINA M, CIOCI G, MOULIS C, et al. Bacterial α-glucan and branching sucrases from GH70 family:Discovery, structure–function relationship studies and engineering[J]. Microorganisms,2021,9(8):1607. doi: 10.3390/microorganisms9081607
[2] HU Y, WINTER V, CHEN X Y, et al. Effect of acceptor carbohydrates on oligosaccharide and polysaccharide synthesis by dextransucrase DsrM from Weissella cibaria[J]. Food Research International,2017,99:603−611. doi: 10.1016/j.foodres.2017.06.026
[3] CHEN Z, NI D, ZHANG W, et al. Lactic acid bacteria-derived α-glucans:From enzymatic synthesis to miscellaneous applications[J]. Biotechnology Advances,2021,47:107708. doi: 10.1016/j.biotechadv.2021.107708
[4] LI X, WANG X, MENG X, et al. Structures, physico-chemical properties, production and (potential) applications of sucrose-derived α-d-glucans synthesized by glucansucrases[J]. Carbohydrate Polymers,2020,249:116818. doi: 10.1016/j.carbpol.2020.116818
[5] WANG C, ZHANG H B, LI M Q, et al. Functional analysis of truncated and site-directed mutagenesis dextransucrases to produce different type dextrans[J]. Enzyme and Microbial Technology,2017,102:26−34. doi: 10.1016/j.enzmictec.2017.03.011
[6] YILMAZ M T, İSPIRLI H, TAYLAN O, et al. Structural and bioactive characteristics of a dextran produced by Lactobacillus kunkeei AK1[J]. International Journal of Biological Macromolecules,2022,200:293−302. doi: 10.1016/j.ijbiomac.2022.01.012
[7] DU R, QIAO X, WANG Y, et al. Determination of glucansucrase encoding gene in Leuconostoc mesenteroides[J]. International Journal of Biological Macromolecules,2019,137:761−766. doi: 10.1016/j.ijbiomac.2019.06.200
[8] 陈琪, 张亚敏, 赵颖, 等. 表达外源谷氨酸脱羧酶基因对重组乳酸乳球菌胁迫抗性的影响[J]. 食品科学, 2018, 39(20):132−139. [CHEN Q, ZHANG Y M, ZHAO Y, et al. Effect of expressing exogenous glutamic acid decarboxylase gene on stress resistance of recombinant Lactococcus lactis[J] Food Science, 2018, 39(20):132−139.] CHEN Q, ZHANG Y M, ZHAO Y, et al. Effect of expressing exogenous glutamic acid decarboxylase gene on stress resistance of recombinant Lactococcus lactis[J] Food Science, 2018, 39(20): 132−139.
[9] 郝凤奇, 李景梅, 杨桂连. 乳酸菌Nisin诱导表达载体的构建和鉴定[J]. 中国预防兽医学报, 2012, 34(8):624−628. [HAO F Q, LI J M, YANG G L, Construction and characterization of Nisin-inducible expression vectors of Lactobacillus casei[J]. Chinese Journal of Preventive Veterinary Medicine, 2012, 34(8):624−628.] HAO F Q, LI J M, YANG G L, Construction and characterization of Nisin-inducible expression vectors of Lactobacillus casei[J]. Chinese Journal of Preventive Veterinary Medicine, 2012, 34(8): 624−628.
[10] 张艳丽, 聂春明, 周利伟, 等. 牛凝乳酶原基因在食品级乳酸乳球菌中的重组表达[J]. 食品科学, 2014, 35(7):123−127. [ZHANG Y L, NIE C M, ZHOU L W, et al. Recombinant expression of bovine rennetogen gene in food-grade Lactococcus lactis[J] Food Science, 2014, 35(7):123−127.] ZHANG Y L, NIE C M, ZHOU L W, et al. Recombinant expression of bovine rennetogen gene in food-grade Lactococcus lactis[J] Food Science, 2014, 35(7): 123−127.
[11] WU M, MA Y, DOU X, et al. A review of potential antibacterial activities of nisin against Listeria monocytogenes:The combined use of nisin shows more advantages than single use[J]. Food Research International,2023,164:112363. doi: 10.1016/j.foodres.2022.112363
[12] FENG F, ZHOU Q, YANG Y, et al. Characterization of highly branched dextran produced by Leuconostoc citreum B-2 from pineapple fermented product[J]. International Journal of Biological Macromolecules,2018,113:45−50. doi: 10.1016/j.ijbiomac.2018.02.119
[13] ZHAO F, SONG Q, WANG B, et al. Purification and immobilization of the soluble and insoluble portions of recombinant lipase by gram-positive enhancer matrix (GEM) particles[J]. International Journal of Biological Macromolecules,2020,145:1099−1105. doi: 10.1016/j.ijbiomac.2019.09.203
[14] 卓倩, 韩煜东, 徐文雨, 等. pNZ8149-IL21基因乳酸菌表达载体的构建与表达特性[J]. 药物生物技术,2019,26(2):105−109. [ZHUO Q, HAN Y D, XU W Y, et al. Construction and expression characterization of pNZ8149 - IL21 gene lactic acid bacteria expression vector[J]. Pharmaceutical Biotechnology,2019,26(2):105−109.] ZHUO Q, HAN Y D, XU W Y, et al. Construction and expression characterization of pNZ8149 - IL21 gene lactic acid bacteria expression vector[J]. Pharmaceutical Biotechnology, 2019, 26(2): 105−109.
[15] 袁晓宇, 许文涛, 黄昆仑, 等. Nisin-rbLF-N融合基因的构建及其在大肠杆菌中的表达[J]. 食品科学,2010,31(15):194−197. [YUAN X Y, XU W T, HUANG K L, et al. Construction of Nisin-rbLF-N fusion gene and its expression in Escherichia coli[J]. Food Science,2010,31(15):194−197.] YUAN X Y, XU W T, HUANG K L, et al. Construction of Nisin-rbLF-N fusion gene and its expression in Escherichia coli[J]. Food Science, 2010, 31(15): 194−197.
[16] RUKYING N, AJINGI Y U S, NOKYOD S, et al. Design, expression and characterization of lactiscin—A novel broad-spectrum peptidic bacteriocin[J]. Biocatalysis and Agricultural Biotechnology, 2023, 52.
[17] FU Y, ZHOU L, KUIPERS O P. Discovery, biosynthesis, and characterization of a lanthipeptide from Bacillus subtilis EH11 with a unique lanthionine ring pattern[J]. Cell Reports Physical Science, 2023, 4(8):101524.
[18] SONG L, MIAO M, JIANG B, et al. Leuconostoc citreum SK24.002 glucansucrase:Biochemical characterisation and de novo synthesis of α-glucan[J]. International Journal of Biological Macromolecules,2016,91:123−131. doi: 10.1016/j.ijbiomac.2016.05.019
[19] WU C, ZHANG J, DU G, et al. Heterologous expression of Lactobacillus casei RecO improved the multiple-stress tolerance and lactic acid production in Lactococcus lactis NZ9000 during salt stress[J]. Bioresource Technology,2013,143:238−241. doi: 10.1016/j.biortech.2013.05.050
[20] WANG B, SONG Q, ZHAO F, et al. Isolation and characterization of dextran produced by Lactobacillus sakei L3 from Hubei sausage[J]. Carbohydrate Polymers, 2019, 223:115111.
[21] 徐波. Lactococcus lactis食品级诱导表达系统构建及铜绿假单胞菌融合外膜蛋白的表达[D]. 南昌:南昌大学, 2006. [XU B. Food-grade expression host construction in Lactococcus lactis and expression of Pseudomonas aeruginosa fusion outer membrane proteins[D], Nanchang:Nanchang University, 2006.] XU B. Food-grade expression host construction in Lactococcus lactis and expression of Pseudomonas aeruginosa fusion outer membrane proteins[D], Nanchang: Nanchang University, 2006.
[22] GUO L, WANG C, BROOS J, et al. Lipidated variants of the antimicrobial peptide nisin produced via incorporation of methionine analogs for click chemistry show improved bioactivity[J]. Journal of Biological Chemistry,2023,299(7):104845. doi: 10.1016/j.jbc.2023.104845
[23] VETTORI M H P B, FRANCHETTI S M M, CONTIERO J. Structural characterization of a new dextran with a low degree of branching produced by Leuconostoc mesenteroides FT045B dextransucrase[J]. Carbohydrate Polymers,2012,88(4):1440−1444. doi: 10.1016/j.carbpol.2012.02.048
[24] YANG Y, PENG Q, GUO Y, et al. Isolation and characterization of dextran produced by Leuconostoc citreum NM105 from manchurian sauerkraut[J]. Carbohydrate Polymers,2015,133:365−372. doi: 10.1016/j.carbpol.2015.07.061
[25] MAINA N H, TENKANEN M, MAAHEIMO H, et al. NMR spectroscopic analysis of exopolysaccharides produced by Leuconostoc citreum and Weissella confusa[J]. Carbohydrate Research,2008,343(9):1446−1455. doi: 10.1016/j.carres.2008.04.012
[26] WANG J, ZHAO X, TIAN Z, et al. Characterization of an exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibet Kefir[J]. Carbohydrate Polymers,2015,125:16−25. doi: 10.1016/j.carbpol.2015.03.003
[27] ZHANG Y, LIU J, HU G, et al. Fusion enzyme design based on the ''channelization'' cascade theory and homogenous dextran product improvement[J]. International Journal of Biological Macromolecules,2022,222:652−660. doi: 10.1016/j.ijbiomac.2022.09.222
[28] DU R, XING H, YANG Y, et al. Optimization, purification and structural characterization of a dextran produced by L. mesenteroides isolated from Chinese sauerkraut[J]. Carbohydrate Polymers,2017,174:409−416. doi: 10.1016/j.carbpol.2017.06.084
[29] GANGOITI J, PIJNING T, DIJKHUIZEN L. Biotechnological potential of novel glycoside hydrolase family 70 enzymes synthesizing α-glucans from starch and sucrose[J]. Biotechnology Advances,2018,36(1):196−207. doi: 10.1016/j.biotechadv.2017.11.001
[30] SUN Y X, LIU J C, YANG X D, et al. Purification, structural analysis and hydroxyl radical-scavenging capacity of a polysaccharide from the fruiting bodies of Russula virescens[J]. Process Biochemistry,2010,45(6):874−879. doi: 10.1016/j.procbio.2010.02.007
[31] PURAMA R K, GOSWAMI P, KHAN A T, et al. Structural analysis and properties of dextran produced by Leuconostoc mesenteroides NRRL B-640[J]. Carbohydrate Polymers,2009,76(1):30−35. doi: 10.1016/j.carbpol.2008.09.018
[32] WANG B, SONG Q, ZHAO F, et al. Purification and characterization of dextran produced by Leuconostoc pseudomesenteroides PC as a potential exopolysaccharide suitable for food applications[J]. Process Biochemistry,2019,87:187−195. doi: 10.1016/j.procbio.2019.08.020
[33] FAUCARD P, GRIMAUD F, LOURDIN D, et al. Macromolecular structure and film properties of enzymatically-engineered high molar mass dextrans[J]. Carbohydrate Polymers,2018,181:337−344. doi: 10.1016/j.carbpol.2017.10.065
[34] İSPIRLI H, DERTLI E. Production of lactose derivative hetero-oligosaccharides from whey by glucansucrase E81 and determination of prebiotic functions[J]. LWT, 2021, 137:110471.
 下载:
下载:








 下载:
下载:






计量
- 文章访问数: 82
- HTML全文浏览量: 12
- PDF下载量: 15



