


Screening of Selenium-enriched Yeasts with Enhanced Aroma and High Alcohol Production by ARTP Mutagenesis
-
摘要: 为了得到产酒精和产香能力较强的枸杞内生富硒酵母菌,进而促进富硒发酵产品的开发和风味的提升,本研究以实验室保藏的枸杞内生酵母菌为实验材料,通过耐硒法和红硒法初筛得到富硒能力较强的酵母菌,采用常压室温等离子体(Atmospheric room temperature plasma,ARTP)诱变技术,经TTC培养基、杜氏小管发酵法、产酒精、产酯产酸、耐受性等实验筛选出增香和高产酒精的目标菌株,并用氢化物发生原子荧光光谱法测定酵母胞内和发酵液中硒含量。结果表明,富硒初筛得到了M1酿酒酵母(Saccharomyces cerevisiae)和M16葡萄牙棒孢酵母(Clavispora lusitaniae),经ARTP诱变分别筛选出产酒精能力较高的S. cerevisiae诱变菌株M1-5和产香能力较高的C. lusitaniae诱变菌株M16-28,M1-5的产酒精能力比M1提高了38.07%,M16-28的产酯能力比M16提升了107.62%,遗传稳定性均良好。经测定M1-5菌株胞内和发酵液中的硒含量分别为204.8和1158 μg/L,M16-28胞内和发酵液中的硒含量分别为666.45和1830 μg/L,因此,菌株M16-28比M1-5的富硒能力强,且发酵液硒含量极显著高于酵母细胞硒含量(P<0.001)。本研究筛选出诱变菌株M1-5产酒精和M16-28产香能力强的富硒酵母具有优良的发酵特性,在发酵食品和富硒功能性产品的挖掘和开发方面具有潜在的应用前景。Abstract: In order to obtain endophytic selenium-rich yeast from Lycium barbarum with strong ability producing alcohol and aroma, and then promote the selenium-rich fermentation products and improve the flavor, this study took the endophytic yeast of L. barbarum stored in the laboratory as experimental materials, and the yeast with strong selenium-rich ability was obtained through the initial screening of selenium-resistant method and red selenium methods. Atmospheric room temperature plasma (ARTP) technology was used to obtain mutant strains and through TTC medium, Duchenne tubule fermentation, alcohol production, ester and acid production, tolerance and other experiments to screen out the target strains of aroma enhancement and high alcohol production. The content of selenium in yeast cell and fermentation liquid was determined by hydride generation atomic fluorescence spectrometry. The results showed that M1 Saccharomyces cerevisiae and M16 Clavispora lusitaniae were obtained by primary selenium screening, the mutagenic strain M1-5 of S. cerevisiae with higher alcohol production capacity and the mutagenic strain M16-28 of C. lusitaniae with higher aroma production capacity were obtained through ARTP mutagenesis. The alcohol production capacity of M1-5 increased by 38.07% compared with M1, and the ester production capacity of M16-28 increased by 107.62% compared with M16, and the genetic stability was good. The selenium contents in the intracellular and fermentation broth of the M1-5 strains were 204.8 and 1158 μg/L, respectively, and in M16-28 were 666.45μg/L and 1830 μg/L, respectively. Therefore, the selenium enrichment capacity of M16-28 was higher than that of M1-5, and the selenium content of fermentation liquid was extremely significantly higher than that of yeast cells (P<0.001). In conclusion, the selenium-rich yeast with mutagenic strain M1-5 producing alcohol and M16-28 producing aroma were screened out, which had excellent fermentation characteristics and potential application prospects in the development and mining of fermented food and selenium-rich functional products.
-
硒(Se)是人体必需微量元素,具有抗氧化、抗衰老、抗癌症、提高机体的细胞免疫及体液免疫水平,降低细胞炎症反应和预防心血管疾病的发生等生物学功能,对于人体的新陈代谢及心脏、肝脏等重要器官起着至关重要的作用[1−2]。但是硒元素在人体中无法自身合成,需要从食物中摄取,包括一些天然富硒产品,比如富硒大蒜、富硒水稻、富硒茶叶、富硒食用菌等,以及一些人工转化富硒食品,比如富硒猪肉等[3]。传统的补硒方法是使用亚硒酸钠,但其毒性较大、生物利用度较低,对人体有着潜在的危害[4]。因此,从食物中获取有机硒是人们追求健康的趋势所需。目前,富硒产品在发酵工业中被广泛用作理想的富硒补充剂,其中,酵母菌起着至关重要的作用,富硒酵母能利用富硒培养基将无机硒转化为有机硒,其生物活性和安全性更高,以促进人体健康[5]。同时,酵母菌是一类具有产酒精、产酯能力的微生物,是传统发酵食品中产酒精、产酯类物质主要产生菌株。目前用于产酒精的酵母菌主要是酿酒酵母,产酯的非酿酒酵母有克鲁维毕赤酵母、葡萄汁有孢汉逊酵母[6]、异常汉逊酵母[7]、戴尔有孢圆酵母[8]、葡萄牙棒孢酵母[9]等。这些产酯酵母在酒的风味中起重要作用,所产生的酯类物质能增强并丰富酒的香气,而己酸乙酯是其中的特色风味物质,但其在天然的产酯酵母中研究较少,且产量低[9]。
选育优良的酵母菌种对于改善发酵特性和品质意义重大。国内外对于酵母菌种的选育方式有自然选育、杂交育种、诱变育种和基因工程等,均具有一定的优缺点。近些年,常压室温等离子体(ARTP)诱变作为诱变育种的一种新型育种方式之一,基于氦射频大气压放电等离子体的新开发的工具,ARTP产生的活性颗粒可以穿透细胞壁和细胞膜,诱导DNA损伤,导致突变率增加,但不产生有毒有害物质,与传统的紫外线辐射或化学诱变剂相比,显示出低成本和环境友好性,且具有操作简便、突变率高等优点,适用于各类微生物菌种选育[10−11]。汪锡武等[12]以啤酒酵母WX-01为出发菌,通过高浓度亚硒酸钠初筛,再经过ARTP诱变处理以及亚硒酸钠抗性平板筛选出一株富硒优势啤酒酵母。林仙菊等[13]用ARTP对雪白白僵菌FIM-1809菌株进行诱变,得到一株遗传稳定性较高的突变高产菌FIM-1809-7,其产环孢菌素A能力较初始菌株提高约39%。Hong等[14]通过三轮ARTP诱变和高盐筛选获得了耐盐酿酒酵母菌株HF-130,且该突变体HF-130的乙醇产量提高了98.8%。此外,ATF1基因在菌株HF-130中过表达,生成产酯菌株HF-130-ATF1。目前,研究者对于枸杞内生酵母的选育大多集中于自然选育,基于ARTP诱变选育的研究少之甚少,并且对于富硒酵母的筛选,有从桑葚[15]、茯砖茶[16]等中进行筛选的,但从枸杞内生酵母菌进行富硒增香酵母菌株的筛选鲜有报道,利用ARTP诱变筛选突出枸杞风味和地域典型性的富硒增香酵母菌株对于开发枸杞相关富硒发酵食品的具有重要意义。
因此,本研究以枸杞内生酵母菌株为出发菌,通过耐硒法和红硒法进行初筛,得到富硒能力较强的菌株M1酿酒酵母(Saccharomyces cerevisiae)和M16葡萄牙棒孢酵母(Clavispora lusitaniae),进而通过ARTP诱变技术对这两株酵母菌进行诱变,以期选育出产酒精和产香能力强的富硒酵母菌株,从而为后续研究富硒酵母的性能优化以及食品发酵提供实验数据支撑,并在枸杞酒生产中,有望突出枸杞品种的典型性和产区的地域性,对枸杞酒风味多样化以及混菌发酵等方面提供理论参考。
1. 材料与方法
1.1 材料与仪器
14株枸杞内生酵母菌包括酿酒酵母(Saccharomyces cerevisiae)M1、葡萄牙棒孢酵母(Clavispora lusitaniae)M16菌株 课题组前期筛选鉴定,−20 ℃保藏菌种(各保藏3株),均是从新鲜枸杞内部经表面消毒后富集培养分离出的内生酵母;酵母浸粉、亚硒酸钠(Na2SeO3) 化学纯,纯度99%,山东高泰化工科技有限公司;2,3,5-三苯基氯化四氮唑(TTC) 纯度98%,Amresco公司;浓盐酸、葡萄糖 均为分析纯,天津致远化学试剂有限公司;酚酞 分析纯,天津津北精细化工有限公司;氢氧化钠 分析纯,徐州天鸿化工有限公司。
ARTP-C1-18ARTP诱变仪 天木生物;SW-CJ-1FD超净工作台 上海析牛科技公司;N4紫外分光光度计、HPS-250恒温恒湿培养箱、KG-SX-500全自动高压灭菌锅、TGL16高速离心机 上海仪电分析仪器有限公司;CP224C电子天平 常州市幸运电子设备有限公司;BPG-9240A烘干箱 上海一恒科技有限公司;PHS-3C pH计 雷磁分析仪器厂;VDRTEX Point-2振荡器 海门市其林贝尔仪器制造有限公司。
1.2 实验方法
1.2.1 溶液及培养基配制
1000 μg/mL的硒标准溶液的配制[17]:准确称取2.1902 g亚硒酸钠(Na2SeO3)溶解于1000 mL无菌水中(只计算硒元素的量),并在无菌操作台内用0.22 μm孔径的水系滤膜过滤;富硒酵母筛选培养基:10 g/L酵母浸粉、20 g/L葡萄糖、20 g/L蛋白胨,121 ℃灭菌20 min,将在无菌操作下配制的1000 μg/mL的硒标准液,分别吸取不同体积按比例配制成不同浓度的富硒酵母培养基。
1.2.2 富硒酵母菌的初筛
将14株内生酵母接种到含有20.00 μg/mL硒的YPD培养基上,分别观察14株内生酵母的生长状况以及颜色变化情况,得到生长良好的酵母菌。分别按5%的接种量接种于含有硒质量浓度为0.00、5.00、10.00、20.00、30.00、40.00、50.00 μg/mL的YPD液体培养基的试管中,28 ℃培养48 h,观察并记录试管颜色的变化和菌体在试管底部的沉积量,通过耐硒法和红硒法对比筛选出富硒能力较强的酵母菌[15]。
1.2.3 富硒酵母硒含量的测定
将所测菌株按5%接种于100 mL/250 mL锥形瓶中进行摇瓶培养至6 h时,在无菌操作下加入2 mL的20 μg/mL硒标准溶液,培养至48 h。将培养好的待测菌液取4 mL在4 ℃离心机8000 r/min下离心10 min,无菌水洗涤离心2~3次,加入4 mL无菌水悬浮,测得酵母菌胞内富硒含量,将离心得到的上清液用0.22 μm除菌过滤器过滤,测得酵母菌胞外硒含量。采用GB 5009.93-2017《食品安全国家标准 食品中硒的测定》中的第一法:氢化物原子荧光光谱法,测定酵母菌液硒含量,重复三次取均值[18]。
1.2.4 菌株M1和M16生长曲线的绘制
按5%接种量于100 mL/250 mL锥形瓶中,在28 ℃的条件下连续培养40 h,每4 h取菌液于比色皿中,并用YPD液体培养基作为对照,通过用分光光度计测定OD600 nm值,每个菌株做3个重复,绘制酵母菌生长曲线。
1.2.5 ARTP诱变试验
试验参数:诱变仪紫外杀菌30 min,气体为高纯度氦气(99.999%),诱变功率120 W,温度20 ℃,载片距发射源2 mm,诱变时间设置为0、10、20、30、40、50、60、70、80、90、100、110、120 s。将灭好菌的载片在火焰上方烤5 s,晾至室温,备用。
诱变试验:将原始菌株M1和M16在28 ℃摇瓶培养,分别取对数时期(16 h)原始菌液,测量OD600 nm下菌液浓度,稀释菌液浓度于0.6~0.8,吸取10 μL菌液于载片上,置于诱变室凹槽中,按上述试验参数设置后即可开始试验,将处理好的载片分别装在有1 mL 0.9%无菌生理盐水的EP管中。诱变处理结束,将上述EP管中的诱变菌株用涡旋振荡器振荡1 min进行洗脱,并将菌液稀释10−1~10−6,分别吸取100 μL涂布在YPD固体培养基上(三个平行),28 ℃培养48 h,记录菌落数,按以下公式计算致死率:
1.2.6 TTC培养基筛选
将诱变出的单菌落接种于YPD固体培养基上,28 ℃培养48 h,将TTC上层培养基用一次性接种环取下倒扣在TTC下层培养基上,28 ℃培养3 h,观察显色情况,对诱变菌株进行筛选,将产酒精能力强的菌株进行下一步筛选。
1.2.7 菌株发酵性能
杜氏小管发酵法:将筛选得到的TTC显色明显变红的突变菌株,按5%的接种于5 mL YPD液体培养基中,28 ℃培养,定时观察菌株产气情况,筛选出发酵性能优良的菌株。
1.2.8 菌株耐受性筛选
将筛选得到的发酵性能优良的突变菌株进行耐受性实验,分别按5%接种量接种于不同浓度的乙醇(4%vol、8%vol、12%vol、16%vol、20%vol)、氯化钠盐溶液(2.5%、5%、10%、15%、20%)、葡萄糖质量浓度(5%、10%、15%、20%、25%)、pH(1.5、2、2.5、3.0、3.5、4.0)的YPD液体培养基内,28 ℃培养48 h,测量OD600 nm的吸光度,以不接种菌株的YPD液体培养基作对照,分别做三个平行[19]。
1.2.9 总酯、总酸测定
总酯:M16菌株及上述筛选的诱变菌株进行摇瓶培养3 d后,用移液枪吸取4 mL菌液,加入40 mL/250 mL的去离子水,滴2滴酚酞指示剂,用0.1 mol/L的NaOH溶液滴定,滴至红色且0.5 min不变色后再加入25 mL 0.1 mol/L的NaOH溶液。沸水浴中45 min后,冷却1 h至室温,随即用0.1 mol/L的HCl溶液滴定至无色,0.5 min不变色,记录HCl溶液的消耗量[20]。总酯含量按以下公式计算:
式中,C1为0.1 mol/L的NaOH溶液浓度,mol/L;C2为0.1 mol /L的HCl溶液浓度,mol/L;V1为滴定0.1 mol/L HCl的消耗量,mL。
总酸:采用标准滴定法,用移液枪吸取2 mL待测样品,并加入150 mL的去离子水于250 mL的锥形瓶中,滴加3滴酚酞指示剂,随即用0.1 mol/L的NaOH溶液滴定至微红色,0.5 min不变色,另做空白组滴定,总酸按以下公式计算。
式中,V1为滴定0.1 mol/L的NaOH溶液消耗量,mL;V2为空白组滴定NaOH消耗的体积,mL。
1.2.10 遗传稳定性试验
将最终得到的诱变菌株M1-5和M16-28分别连续传代培养5次,分别对诱变菌株的遗传稳定性进行验证。M1-5菌株每一代菌株以5%接种量接种于YPD液体培养基培养8 d,测量其酒精度的含量,M16-28以5%的接种量接种于YPD液体培养基培养3 d,测定产酯产酸能力。
1.2.11 发酵能力验证
酒精度的测定[21]:参照GB 5009.225-2016《食品安全国家标准 酒中乙醇浓度的测定》方法测定。蒸馏后用密度瓶法测出馏出液20 ℃时的密度,查询酒精水溶液密度与酒精度(乙醇含量)对照表,得到乙醇体积分数,即为酒精度。
1.3 数据处理
每组实验重复测定三次,结果均用平均值±标准差表示。采用SPSS 23.0软件对试验数据进行显著性检验,P>0.05为差异不显著,P<0.05为差异显著,P<0.001为差异极显著,其余数据采用Excel 2016 软件进行统计分析及绘图。
2. 结果与分析
2.1 富硒酵母菌的初筛
经富硒酵母筛选固体培养基培养,筛选出9株长势较好的酵母菌,为JY-1、M、G2、M16、M19、G1、M1、M17、B7,进行液体培养筛选。耐硒法是基于微生物对无机硒的抗性与其对无机硒的生物转化能力成正相关,即当菌体沉淀量变化较小,菌株对无机硒亚硒酸钠的耐受性较强;红硒法是基于微生物对硒的生物转化能力与红硒的形成呈负相关,即菌体沉淀颜色变化越浅,则酵母菌对亚硒酸钠的转化能力强[15]。由表1可知,当加入的硒质量浓度增加时,各株菌的菌体的沉淀量逐渐减少,在20.00 μg/mL的硒浓度下,菌株M、G2、M16和M1的菌体沉淀量与自身初始未加硒母液沉淀量变化较小,则说明菌株M、G2、M16和M1对无机硒亚硒酸钠的耐受性较强。由表2可知,随着加入的硒质量浓度增加,菌体沉淀量的颜色也发生了明显逐渐加深的现象,在20.00 μg/mL的硒浓度下,菌株JY-1、M16、G1和M1显示为粉红色,菌株B7无沉淀,其他菌体显示为红色,表明菌株JY-1、M16、G1和M1对亚硒酸钠的转化能力强。结合耐硒法和红硒法,最终选择M16和M1作为后续试验菌株。
表 1 不同酵母菌在富硒培养液中的菌体沉淀量Table 1. Precipitation amount of different yeasts in selenium-rich medium菌株编号 硒质量浓度(μg/mL) 0.00 5.00 10.00 20.00 30.00 40.00 50.00 JY-1 **** **** *** ** * * * M **** **** **** *** *** *** *** G2 **** **** **** *** *** *** *** M16 **** **** *** *** ** ** * M19 **** *** ** ** ** ** * G1 **** **** *** * * * * M1 **** **** **** **** *** *** ** M17 **** *** *** ** * * * B7 **** *** *** − − − − 注: “****”表示6/6(每株菌在加硒后菌体沉淀量与初始未加硒菌体沉淀量的比值,以下以此类推);“***”表示5/6,“**”表示4/6;“*”表示3/6,“−”表示无沉淀。 表 2 不同酵母菌在富硒培养液中的颜色比较Table 2. Color comparison of different yeasts in selenium-rich medium菌株编号 硒质量浓度(μg/mL) 0.00 5.00 10.00 20.00 30.00 40.00 50.00 JY-1 − * ** ** *** *** *** M − * ** *** *** **** **** G2 − * ** *** *** *** *** M16 − * ** ** *** *** *** M19 − * ** *** *** **** **** G1 − * ** ** *** **** **** M1 − − * ** *** *** **** M17 − * ** *** **** **** **** B7 − * ** − − − − 注:“−”代表正常;“*”微红色;“**”粉红色;“***”红色;“****”深红色。 2.2 菌株M1和M16生长曲线的绘制
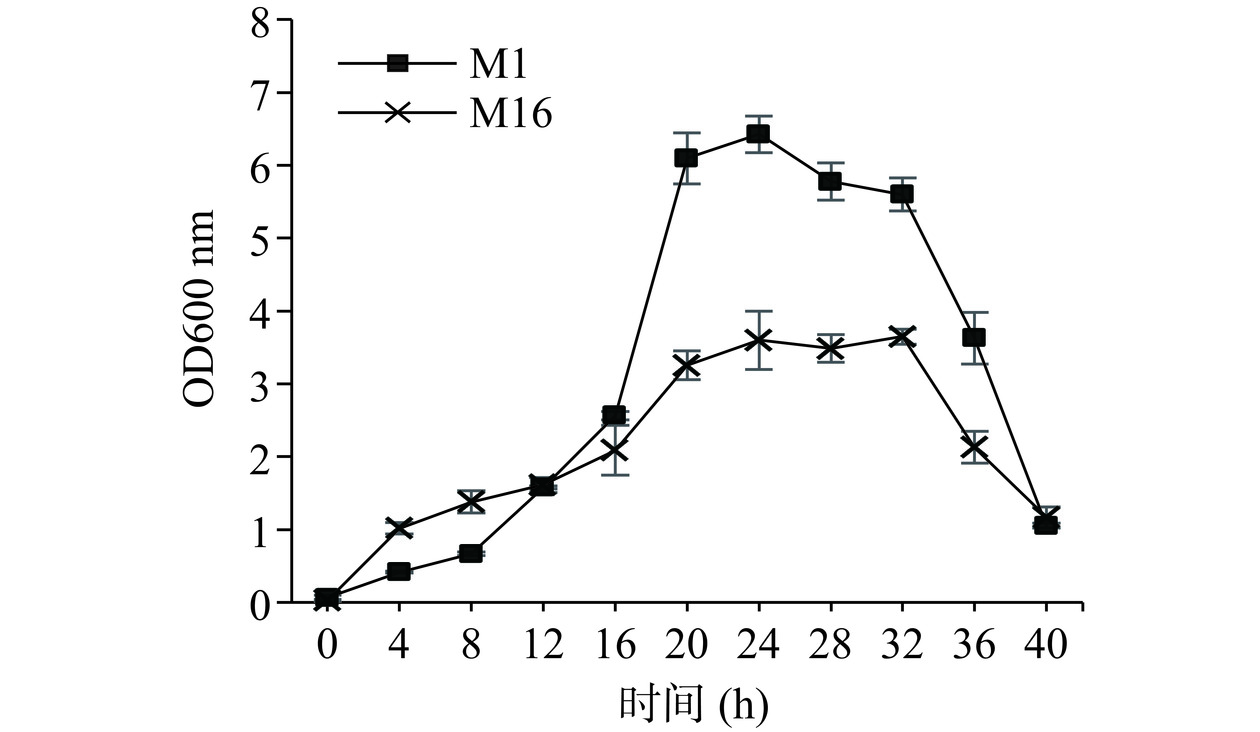
对原始菌株进行生长曲线绘制,以便更好确定其对数生长时期,利于后续诱变筛选。2株酵母生长曲线如图1所示。在0~8 h,菌株M1处于延滞期,而菌株M16的延滞期大致处于4 h前,之后各进入对数期,菌株快速生长,此时期菌株对环境敏感,易受培养基组分及物理化学条件影响,20~32 h两株菌都进入了稳定期。32 h后均缓慢下降,进入衰亡期。
2.3 ARTP诱变时间的确定
在诱变时,突变率会随着诱变处理时间的延长而逐渐增加,但时间过长,会导致死亡率过高、诱变效果变差。ARTP诱变的时间致死率在80%~95%,更有利于后续的筛选工作。如果致死率高于95%时,会导致菌株整体的生长性能受损,对成品酒的风味会产生不利的影响;如果突变率过于低的话,会导致菌株突变效率不高[22]。原始菌株M1和M16分别在对数时期16 h时经ARTP诱变后致死率曲线如图2所示。当诱变时间延长时,致死率增大。当诱变时间达到40 s时,M1的致死率为90.31%,在50 s时,M16的致死率为90.26%,110 s时,两株菌的致死率均达到100%,未发现有活菌存活。当致死率80%~95%时,菌株突变概率较高,突变幅度较大。因此,M1和M16各选40 s和50 s作为ARTP诱变最佳时间。
![]() 图 2 不同诱变时间下菌株M1和M16的致死率Figure 2. Mortality of strains M1 and M16 under different mutagenesis times
图 2 不同诱变时间下菌株M1和M16的致死率Figure 2. Mortality of strains M1 and M16 under different mutagenesis times2.4 诱变酵母菌株初筛
2.4.1 TTC培养基筛选
TTC显色剂用于培养菌株时观察颜色的深浅,颜色越深时,说明菌株呼吸酶的活力越旺盛,产酒精能力越强。经ARTP诱变后,将原始菌株M1及其诱变获得的44株诱变菌株分别重新接种划线于TTC下层培养基上,经筛选得到的9株产酒精能力较强的诱变菌株,显色情况如表3所示。原始菌株M1呈浅粉色,菌株M1-5、M1-11、M1-13、M1-19、M1-24、M1-30、M1-34、M1-36和M1-40显示为红色。
表 3 M1菌株产酒精能力测定结果Table 3. Determination results of alcohol production capacity of M1 strains菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色情况 菌株编号 显色情况 M1 + M1-9 + M1-18 + M1-27 + M1-36 +++ M1-1 ++ M1-10 + M1-19 +++ M1-28 + M1-37 + M1-2 + M1-11 +++ M1-20 ++ M1-29 + M1-38 ++ M1-3 ++ M1-12 ++ M1-21 ++ M1-30 +++ M1-39 ++ M1-4 ++ M1-13 +++ M1-22 ++ M1-31 ++ M1-40 +++ M1-5 +++ M1-14 ++ M1-23 ++ M1-32 ++ M1-41 + M1-6 ++ M1-15 ++ M1-24 +++ M1-33 ++ M1-42 ++ M1-7 + M1-16 ++ M1-25 ++ M1-34 +++ M1-43 ++ M1-8 ++ M1-17 + M1-26 ++ M1-35 + M1-44 ++ 注:“+++”呈红色;“++”呈粉红色;“+”呈浅粉色。 经ARTP诱变后,将原始菌株M16及其诱变获得的58株诱变菌株分别重新接种划线于TTC下层培养基上,经筛选得到的9株产酒精能力较强的诱变菌株,显色情况如表4所示。原始菌株M16显红色,菌株M16-14、M16-17、M16-28、M16-40、M16-41、M16-42、M16-44、M16-47和M16-48呈深红色,将筛选得到的9株诱变菌株进行发酵能力筛选试验。
表 4 M16菌株产酒精能力测定结果Table 4. Determination results of alcohol production capacity of M16 strains菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色情况 菌株编号 显色情况 M16 +++ M16-12 +++ M16-24 +++ M16-36 +++ M16-48 ++++ M16-1 +++ M16-13 +++ M16-25 +++ M16-37 +++ M16-49 +++ M16-2 +++ M16-14 ++++ M16-26 +++ M16-38 +++ M16-50 +++ M16-3 +++ M16-15 +++ M16-27 +++ M16-39 +++ M16-51 +++ M16-4 +++ M16-16 +++ M16-28 ++++ M16-40 ++++ M16-52 +++ M16-5 +++ M16-17 ++++ M16-29 +++ M16-41 ++++ M16-53 +++ M16-6 +++ M16-18 +++ M16-30 +++ M16-42 ++++ M16-54 +++ M16-7 +++ M16-19 +++ M16-31 +++ M16-43 +++ M16-55 +++ M16-8 +++ M16-20 +++ M16-32 +++ M16-44 ++++ M16-56 +++ M16-9 +++ M16-21 +++ M16-33 +++ M16-45 +++ M16-57 ++ M16-10 +++ M16-22 +++ M16-34 +++ M16-46 +++ M16-58 +++ M16-11 +++ M16-23 +++ M16-35 +++ M16-47 ++++ 注:“++++”呈深红色;“+++”呈红色。 2.4.2 杜氏小管发酵能力筛选
对M1的诱变得到的9株诱变菌株进行杜氏小管发酵产气实验,结果如表5所示。2 h时观察到诱变菌株M1-5、M1-13、M1-40有气体产生,说明其起酵速度快,且M1-13和M1-40比M1-5起酵能力好;在3 h时观察到诱变菌株除M1-36、M1-19外,其余菌株均有气体产生且M1-13和M1-40发酵能力强,杜氏小管中气体生成量为2/5;在4 h时,诱变菌株M1-5、和M1-11杜氏小管中气体生成量为2/5,M1-13、M1-40气体生成量为3/5;当培养至5 h后诱变菌株M1-5、M1-13、M1-30、M1-40的气体充满了杜氏小管;培养6 h原始菌株M1产生的气体也充满小管,M1-19、M1-24、M1-34、M1-36已产生4/5气体;7 h后所有菌株均能将杜氏小管中充满气体;菌株M1-11的起酵速度和产气情况与原始菌株M1一致。综合考虑,选择起酵能力快,发酵能力强的诱变菌株M1-5、M1-13、M1-30、M1-40作为乙醇耐受性、氯化钠盐溶液耐受性、高糖耐受性、pH耐受性试验的出发菌株。
表 5 M1菌株杜氏小管产气结果Table 5. Du's tubule gas production result of M1 strain菌株编号 培养时间(h) 1 2 3 4 5 6 7 M1 − − + ++ ++++ +++++ +++++ M1-5 − + + ++ +++++ +++++ +++++ M1-11 − − + ++ ++++ +++++ +++++ M1-13 −+ + ++ +++ +++++ +++++ +++++ M1-19 − − − + +++ ++++ +++++ M1-24 − − −+ + +++ ++++ +++++ M1-30 − − −+ + +++++ +++++ +++++ M1-34 − − + + +++ ++++ +++++ M1-36 − − − + +++ ++++ +++++ M1-40 −+ + ++ +++ +++++ +++++ +++++ 注:杜氏小管中气体充斥量:“+++++”表示5/5;“++++”表示4/5;“+++”表示3/5;“++”表示2/5;“+”表示1/5;“−+”表示小于1/5;“−”表示无气体产生。 对M16的诱变得到的9株诱变菌株进行杜氏小管发酵产气实验,结果如表6所示。诱变菌株M16-41、M16-42、M16-44培养2 h就观察到有气体产生,说明诱变菌株起酵能力迅速;培养4 h后诱变菌株除M16-17、M16-47外,其余菌株均有气体产生;培养6 h后诱变菌株M16-42、M16-44、M16-48菌株产生较其他菌株较多,培养8 h后诱变菌株M16-28、M16-40、M16-42、M16-47已在杜氏小管中产生3/5气体,M16-44、M16-48已在杜氏小管中产生4/5气体;培养10 h后所有菌株均能将杜氏小管中充满气体;菌株M16-14的起酵速度和产气情况与原始菌株M16一致。综合考虑,选择起酵能力快,发酵能力强的诱变菌株M16-28、M16-40、M16-42、M16-44、M16-48作为乙醇耐受性、氯化钠盐溶液耐受性、高糖耐受性、pH耐受性试验的出发菌株。
表 6 M16菌株杜氏小管产气结果Table 6. Du's tubule gas production result of M16 strain菌株编号 培养时间(h) 2 4 6 8 10 12 M16 − −+ + ++ +++++ +++++ M16-14 − −+ + ++ +++++ +++++ M16-17 − − + + ++++ +++++ M16-28 − −+ + +++ +++++ +++++ M16-40 − −+ + +++ +++++ +++++ M16-41 −+ −+ + ++ +++++ +++++ M16-42 −+ −+ ++ +++ +++++ +++++ M16-44 −+ −+ +++ ++++ +++++ +++++ M16-47 − − + +++ +++++ +++++ M16-48 − −+ ++ ++++ +++++ +++++ 注:杜氏小管中气体充斥量:“+++++”表示5/5;“++++”表示4/5;“+++”表示3/5;“++”表示2/5;“+”表示1/5;“−+”表示小于1/5;“−”表示无气体产生。 2.5 诱变菌株耐受性筛选
2.5.1 乙醇耐受性
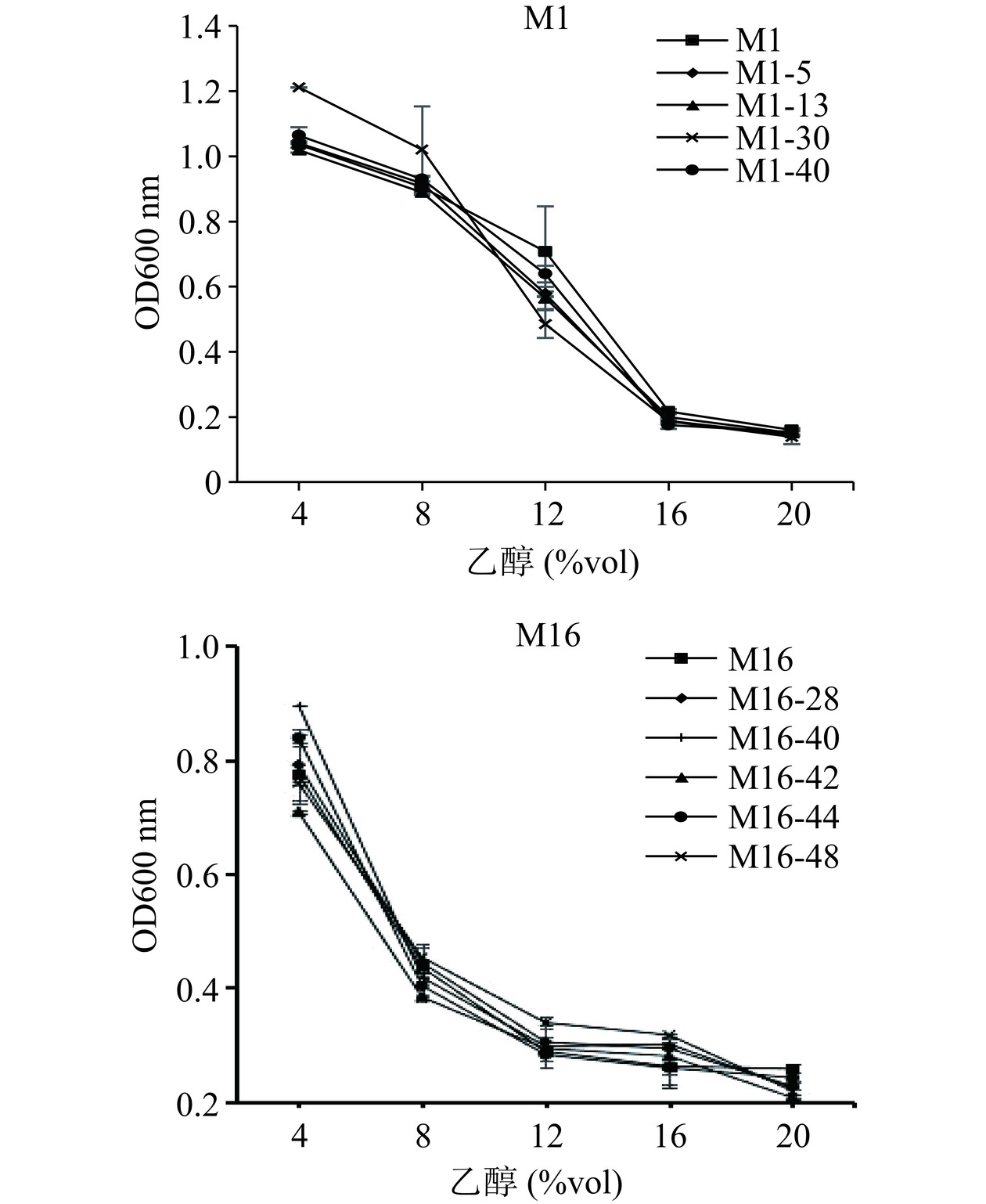
传统的非酿酒酵母对乙醇的耐受性较低,发酵周期较短。因此,筛选高酒精耐受性非酿酒酵母,能进一步提高其存活率,使后续发酵产品风味物质增加。不同酒精度对菌株活性影响如图3所示。随着乙醇添加量增多,菌株的生长代谢抑制变强。对于M1菌株,酒精度在8%vol以内,诱变菌株M1-30的生长状况要优于其他菌株,酒精度在8%vol~16%vol,诱变菌株M1-30表现出较低的乙醇耐受性,当酒精度高于16%vol时,所有菌株几乎不生长。M16菌株酒精度在8%vol以内,原始菌株M16以及诱变菌株M16-28、M16-40、M16-44、M16-48对乙醇的耐受性整体优于诱变菌株M16-42;酒精度在8%vol~16%vol,诱变菌株M16-48表现出较高的乙醇耐受性,当酒精度高于20%vol时,所有菌株几乎不生长。
![]() 图 3 不同酒精度对菌株生长的影响Figure 3. Effects of different alcohol content on the growth of strains
图 3 不同酒精度对菌株生长的影响Figure 3. Effects of different alcohol content on the growth of strains2.5.2 高盐耐受性
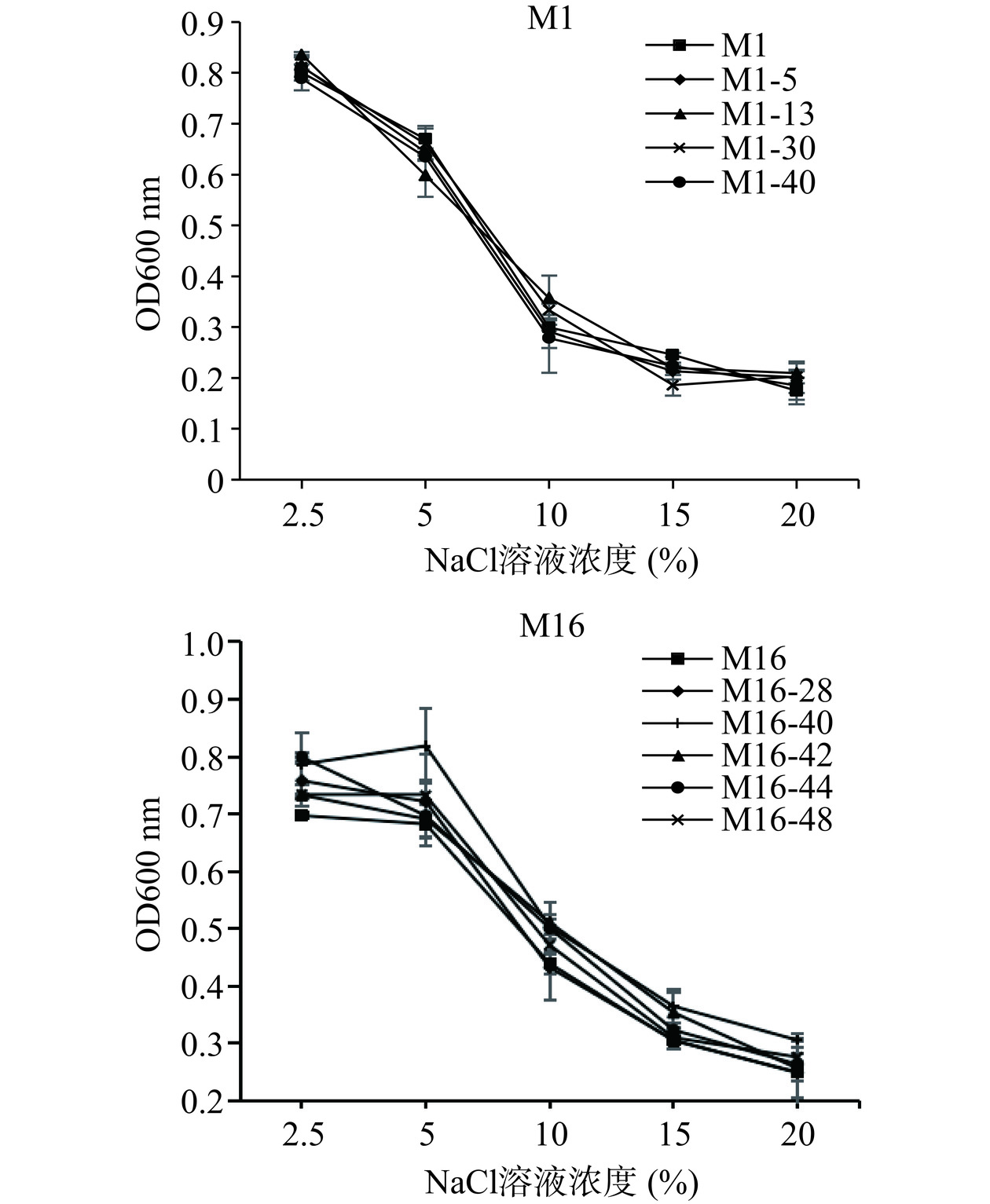
高渗透压胁迫主要发生在刚开始发酵时,使酵母细胞形态改变,发生失水现象,使其生长缓慢;也会使有害溶质进入细胞内,进而改变酵母存活率[23]。由图4可知,高渗透压对酵母生长具有明显的抑制作用。对于M1菌株而言,氯化钠浓度为2.5%时,各菌株均呈现出良好的生长状况;在氯化钠浓度在5%~15%范围内,高渗透压对原始菌株M1以及M1的诱变菌株的抑制作用逐级增加,M1-30的抑制作用尤为明显,当氯化钠浓度为15%以上时,所有菌株几乎不生长,对此渗透压条件耐受性差异不大;对于M16菌株而言,所有诱变菌株均比原始菌株M16耐受性好,且诱变菌株M16-40的耐受性优于其他菌株;氯化钠浓度为2.5%~5%时,各菌株均呈现出良好的生长状况,且诱变菌株M16-40和M16-44优于其他菌株;在氯化钠浓度在5%~20%范围内,高渗透压对原始菌株M16以及M16的诱变菌株的抑制作用逐级增加;当氯化钠浓度为20%以上时,所有菌株几乎不生长。
![]() 图 4 不同NaCl溶液浓度对菌株生长的影响Figure 4. Effect of different NaCl solution concentration on the growth of strain
图 4 不同NaCl溶液浓度对菌株生长的影响Figure 4. Effect of different NaCl solution concentration on the growth of strain2.5.3 糖度耐受性
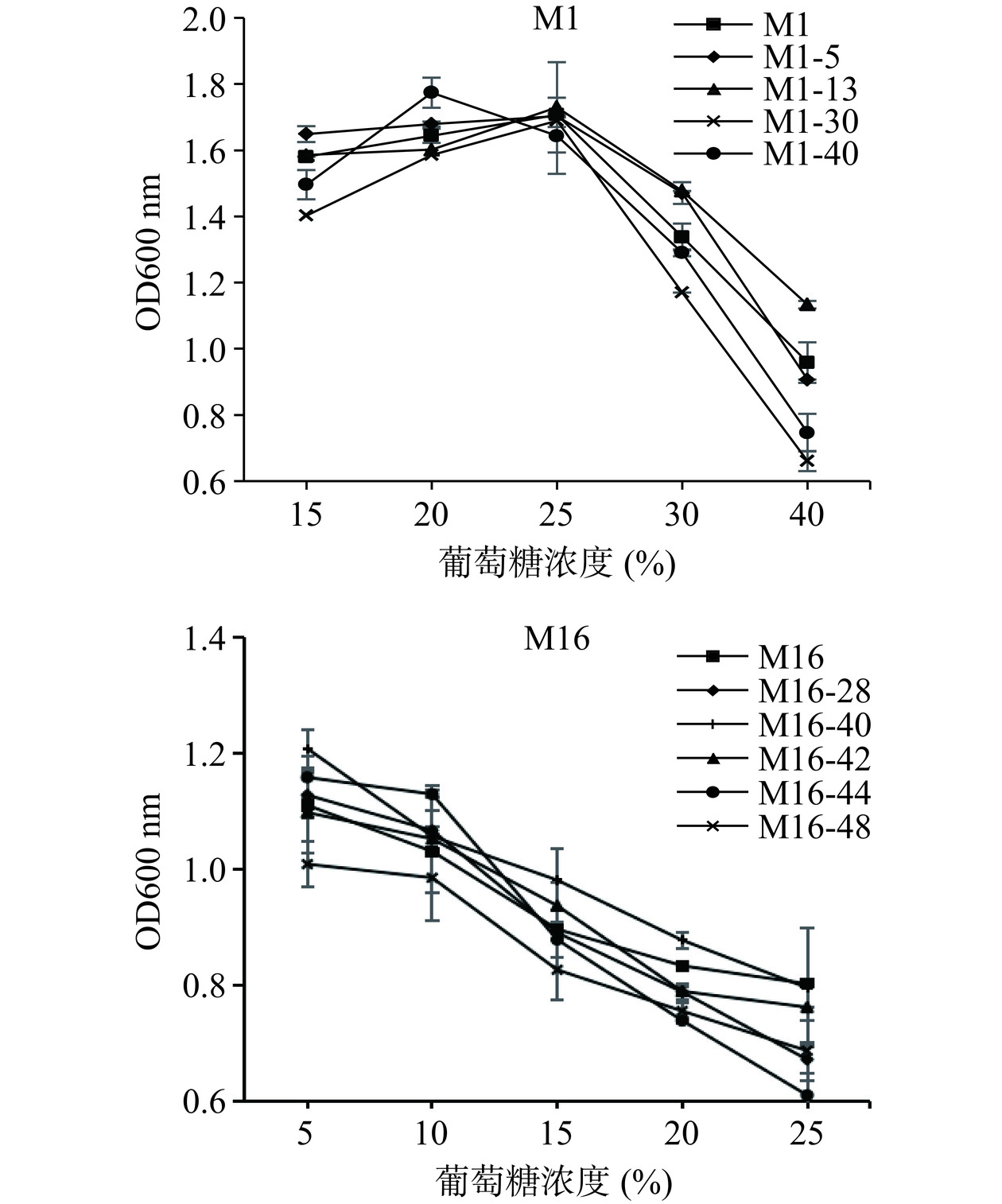
发酵时,糖浓度太高会胁迫酵母细胞的生长和繁殖。菌株对葡萄糖的耐受性如图5所示。对于M1菌株,随着糖浓度的增加,酵母菌体浓度先增加后减少,在培养基糖浓度为20%时,诱变菌株M1-40的OD600 nm值达到最大为1.774,表现出最好的生长状况,当浓度为25%时,原始菌株M1、M1-5、M1-13和M1-30的OD600 nm值达到最大。整体来看,M1-30对高糖的耐受能力较差。对于原始M16菌株以及M16的诱变菌株均随着糖浓度的增加,菌体浓度逐渐降低,细胞生长受糖浓度的抑制程度逐渐增加,在糖浓度为5%时,生长状况最好,在糖浓度为25%时,生长繁殖明显减弱,对高糖的耐受性较差,且M16-48菌株的整体耐受性较差。
![]() 图 5 不同葡萄糖质量浓度对菌株生长的影响Figure 5. Effect of different glucose mass concentration on the growth of strain
图 5 不同葡萄糖质量浓度对菌株生长的影响Figure 5. Effect of different glucose mass concentration on the growth of strain2.5.4 低pH耐受性试验
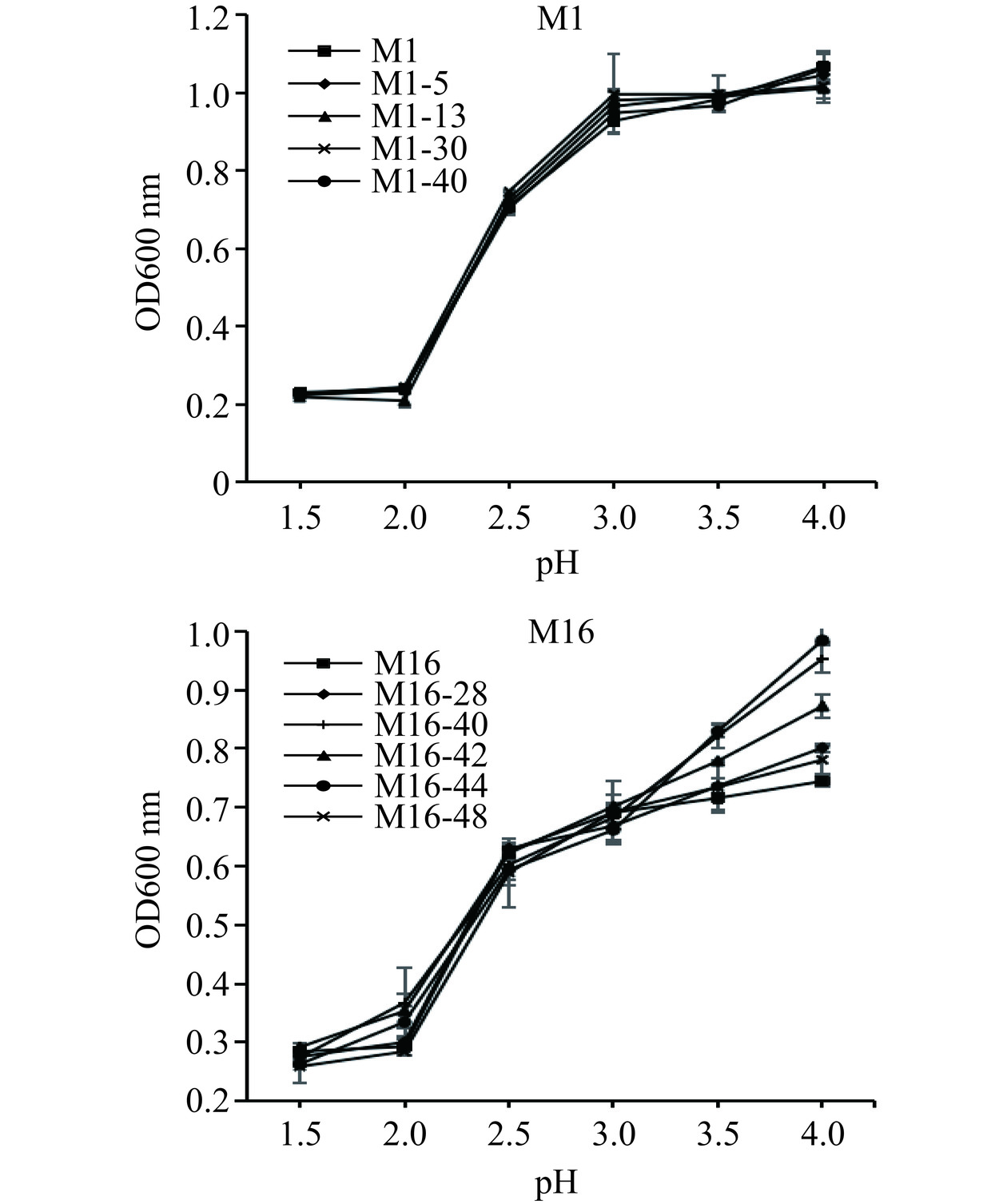
发酵时,酸度过高会抑制酵母菌代谢生长甚至使得酵母菌死亡。因此,筛选出有一定的耐酸能力的酵母是有必要的。不同pH下菌株的生长状况如图6所示。原始菌株M1和M16以及所有诱变菌株均在pH为2下抑制严重;当pH增大,所有菌株生长代谢均有了增强的趋势;pH在3~3.5此范围内发酵时会抑制杂菌生长,利于酒的品质。因此,原始菌株M1和M16及其所有诱变菌株在此酸度范围内能较好生长。随着pH的增大,OD值增大。所有M1菌株在pH为2以下基本不生长,所有M16菌株在pH为2以下生长缓慢,在pH为2以上,所有菌株均开始快速的生长。在pH为3~4之间,原始菌株M1、M16以及所有的诱变菌株均生长良好。
综上所述,结合菌株耐受性分析,选择诱变菌株M1-5、M1-13、M1-40进行高产酒精的复筛实验,选择诱变菌株M16-28、M16-40、M16-44进行下一步产香复筛。
2.6 诱变酵母菌株复筛
2.6.1 M1酿酒酵母的复筛
将M1及其耐受性强的3株诱变菌株在发酵8 d后,测定产酒精含量,如表7所示。发酵结束后,诱变菌株M1-5中总酒精含量显著增强(P<0.05),为0.907%vol,产酒精能力相比于原始菌株M1提升了38.071%;诱变菌株M1-40中总酒精含量虽然也显著增强(P<0.05),但产酒精能力仅比于M1提升了7.107%;M1-13与原始菌株M1无显著差异(P>0.05)。根据产酒精能力测试,综合分析筛选出产酒精能力最优的诱变菌株M1-5。
表 7 酿酒酵母菌株的酒精含量Table 7. Alcohol content of the Saccharomyces cerevisiae strains菌株 酒精(%vol) 提升百分比(%) M1 0.657±0.0153c − M1-5 0.907±0.0252a 38.071 M1-13 0.673±0.0306bc 2.838 M1-40 0.703±0.0208b 7.107 注:不同字母表示差异显著(P<0.05)。 2.6.2 M16产香非酿酒酵母复筛
酯类化合物在很大程度上影响发酵酒浓郁的水果香和花香,生成不同风味和特点的酒香。对M16及其耐受性强的3株诱变菌株的总酯、总酸含量测定,如表8所示。培养3 d后,诱变菌株M16-28发酵液中总酯和总酸均显著高于原始菌株M16(P<0.05),分别为2.0347 g/100mL、1.575 g/L。诱变菌株M16-40、M16-44与原始菌株M16产酯能力差异不显著(P>0.05),但菌株M16-40的产酸要显著高于原始菌株M16(P<0.05)。综合根据产酯、产酸能力测定,分析筛选出产香能力最优的诱变菌株M16-28。
表 8 产香非酿酒酵母菌株总酯、总酸含量Table 8. Total ester and total acid content of nonSaccharomyces cerevisiae strains菌株 总酯(g/100 mL) 总酸(g/L) M16 0.98±0.007b 1.125±0.031b M16-28 2.0347±0.041a 1.575±0.045a M16-40 1.05±0.073b 1.575±0.009a M16-44 1.06±0.058b 1.125±0.021b 注:同列数据不同字母表示差异显著(P<0.05),相同字母表示差异不显著(P>0.05)。 2.7 诱变菌株遗传稳定性测试
2.7.1 诱变菌株M1-5产酒精能力稳定性
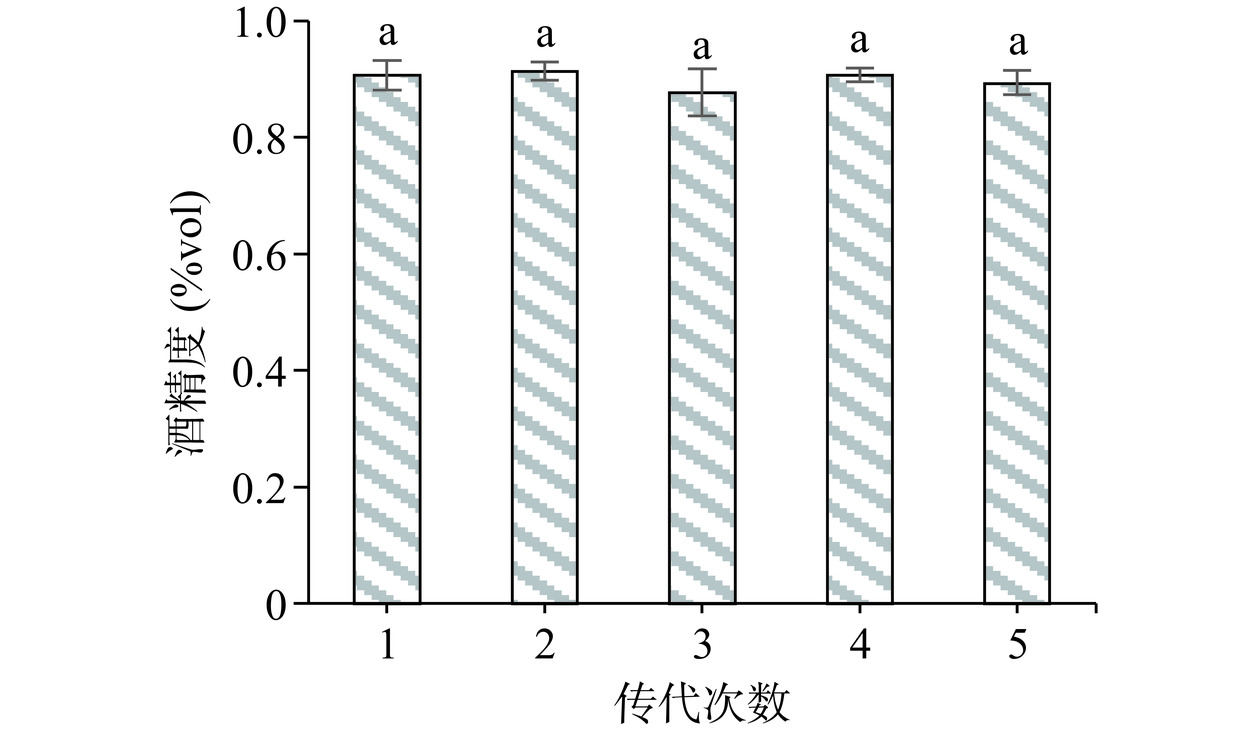
将诱变菌株M1-5连续传代5次,对产酒精能力进行测定,如图7所示。诱变菌株M1-5产酒精能力稳定在0.877%vol~0.913%vol之间,各代菌株总酒精含量无显著差异(P>0.05)。因此,可以证明诱变菌株M1-5产酒精能力可以稳定遗传。
![]() 图 7 M1-5菌株遗传稳定性测试结果注:相同字母表示差异不显著(P>0.05)。Figure 7. Genetic stableility test results of the M1-5 strain
图 7 M1-5菌株遗传稳定性测试结果注:相同字母表示差异不显著(P>0.05)。Figure 7. Genetic stableility test results of the M1-5 strain2.7.2 诱变菌株M16-28产香能力稳定性
将诱变菌株M16-28连续传代5次,对产酯产酸能力进行测定,如图8所示,各代菌株总酯、总酸含量变化范围小,诱变菌株M16-28产酯能力之间无显著差异(P>0.05);产酸能力之间也无显著差异(P>0.05)。因此,可以证明诱变菌株M16-28产酯、产酸能力可以稳定遗传。
![]() 图 8 M16-28菌株遗传稳定性测试结果注:相同字母表示差异不显著(P>0.05),不同大小写的字母用以区分不同组间差异。Figure 8. Genetic stableility test results of the M16-28 strain
图 8 M16-28菌株遗传稳定性测试结果注:相同字母表示差异不显著(P>0.05),不同大小写的字母用以区分不同组间差异。Figure 8. Genetic stableility test results of the M16-28 strain2.8 酵母菌硒含量测定
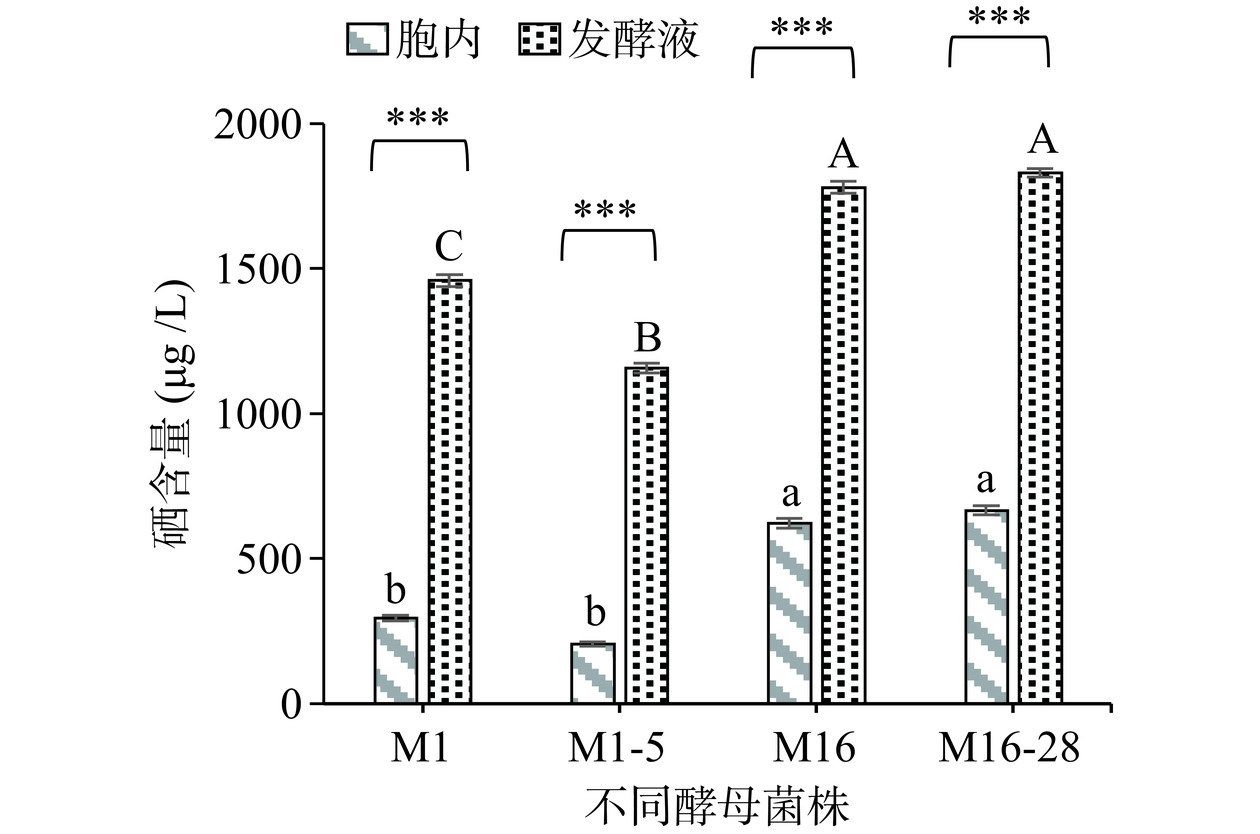
对上述诱变产酒精和产香能力高得菌株及其原始菌株进行了硒含量的测定,如图9所示。在硒质量浓度为20.00 μg/mL时,酿酒酵母M1及其诱变菌株M1-5,葡萄牙棒孢酵母M16及其诱变菌株M16-28的酵母细胞内的硒含量分别为294.9、204.8、622.45、666.45 μg/L,发酵液中的硒含量分别为:1459.5、1158、1780、1830 μg/L。由此可见,发酵液中的硒含量要极显著高于酵母细胞内的硒含量(P<0.001),并且菌株M16和M16-28酵母菌无论是在细胞内的硒含量,还是在发酵液中所测得的硒含量与M1、M1-5均存在极显著差异(P<0.001)。因此,菌株M16和M16-28比M1和M1-5的富硒能力强,其中诱变菌株M16-28比原始菌株M16的硒含量高,但诱变菌株M1-5比原始菌株M1的硒含量低,可能是ARTP诱变时等离子束照射改变了富硒酵母菌对硒的富集能力。以上酵母菌有望用于后续富硒产品发酵应用中。
![]() 图 9 原始菌及诱变菌株胞内和发酵液的硒含量注:不同字母表示差异极显著(P<0.001),不同大小写的字母用以区分不同组间差异,***表示各自组内之间差异极显著(P<0.001)。Figure 9. Selenium content of intracellular and fermentation broth of primitive bacteria and mutagenic strains
图 9 原始菌及诱变菌株胞内和发酵液的硒含量注:不同字母表示差异极显著(P<0.001),不同大小写的字母用以区分不同组间差异,***表示各自组内之间差异极显著(P<0.001)。Figure 9. Selenium content of intracellular and fermentation broth of primitive bacteria and mutagenic strains3. 讨论
本研究以实验室保藏本土枸杞内生酵母M1酿酒酵母(Saccharomyces cerevisiae)和M16葡萄牙棒孢酵母(Clavispora lusitaniae)为原始菌株,经ARTP诱变筛选M1-5的产酒精能力比M1提高了38.07%,M16-28的产酯产酸能力均有了提高,产酯能力比M16提升了107.62%。ARTP技术为酵母菌株的改良和筛选做出了重大贡献。它已被证明可有效生产具有特定性状的各种酵母突变体,例如高产酵母,耐盐酵母和耐酸酵母等[11,24−25]。Liu等[26]通过ARTP突变和驯化,突变菌株LAL-8a的乙醛比M88少14%。此外,LAL-8a的酯与高级醇的比率(0.28)高于M14(0.16)。Wei等[27]经ARTP诱变酿酒酵母LF1,诱变菌株LF1-6表现出显著增强的耐受性,LF6-6通过ARTP诱变迭代突变,6M-15的发酵时间最短,乙醇产率最高,分别为28 h和0.525 g/g/h。王伟雄[20]对新疆本土非酿酒酵母马克斯克鲁维酵母(Kluyveromyces marxianus)YK诱变,诱变菌株YK-29发酵液中总酯、总酸含量(2.15 g/100mL、0.135%)增加,产香能力得到了提升。王犁烨等[28]从葡萄表面筛选的Y6菌株进行ARTP诱变,诱变菌株Y6-8的产酒精、产酸能力分别提高了28.13%和214.93%,对新疆葡萄酒的品质有了很大提升。本研究中,诱变菌株M1-5和M16-28的产酒精和产香能力分别有了显著提升(P<0.05),且两株菌均是枸杞内生酵母菌,目前有关枸杞内生菌的相关研究较少,后续可以用于枸杞发酵的相关产品中,以期提高宁夏枸杞相关发酵产品品质,进一步提升枸杞地域典型性和产品风味多样化。
本研究对实验室现有的枸杞内生酵母通过耐硒法和红硒法进行初筛得到富硒能力较强的酵母菌,并在选育高产酒精和产香的基础上定量测定M1、M1-5、M16、M16-28的胞内和发酵液中硒含量,发现M16及诱变菌株M16-28的硒含量均极显著高于M1及M1-5(P<0.001),且发酵液中的硒含量要极显著高于酵母胞内的硒含量(P<0.001),但本研究没有通过ARTP诱变提升酵母硒含量,由于ARTP诱变具有突变不确定性等特点,所以得到的诱变菌株M16-28硒含量并没有得到很大提升,而且M1-5菌株的硒含量减少。汪锡武等[12]人以啤酒酵母WX-01为出发菌,通过高浓度亚硒酸钠初筛,再经过ARTP诱变处理以及亚硒酸钠抗性平板筛选,选育出一株富硒优势啤酒酵母,硒含量达到1475 μg/g。蔡飞等[29]以分离鉴定得到菌株Z5库德毕赤酵母(Pichia kudriavzevii)通过单因素实验和正交试验,探究各影响因素对菌株Z5富硒能力的影响,在最佳优化条件下,总硒含量可达到862 μg/g。凡敏等[30]将泡菜汁样品筛选出一株耐硒菌株光滑假丝酵母FXY-4,当Na2SeO3的浓度为0.5 g/L时,菌株富硒能力达0.54 mg/g,添加在鱼饲料中,可以增强鱼免疫和促进鱼生长的活性,营养价值提高。目前,许多研究者对硒含量的测定是不区分胞内和发酵液的硒含量的,本研究中通过将两者分开测定,也明确了富硒酵母菌的硒含量主要存在于发酵液。虽经诱变得到的酵母M1-5和M16-28菌株富硒能力较强,但没有进一步的进行优化,在后续的研究中,可对其硒含量进行优化,有望对枸杞富硒产品进行开发和挖掘,提升产品中有机硒含量,强化其营养价值。
4. 结论
本研究以实验室保藏枸杞内生酵母菌株为实验菌,利用耐硒法和红硒法进行初筛得到富硒能力较强的M1酿酒酵母(Saccharomyces cerevisiae)和M16葡萄牙棒孢酵母(Clavispora lusitaniae),经ARTP诱变后对菌株进行耐受性筛选、M1的产酒精能力和M16的产香能力复筛,分别获得产酒精能力提升了38.07%的M1诱变菌株M1-5和产酯能力提升了107.62%的M16诱变菌株M16-28。并测定M1、M1-5、M16和M16-28酵母菌的胞内和发酵液中的硒含量,胞内的硒含量分别为294.9、204.8、622.45、666.45 μg/L,发酵液中的硒含量分别为:1459.5、1158、1780、1830 μg/L。发现M16及M16-28菌株的硒含量均极显著高于M1及M1-5菌株(P<0.001),且发酵液中的硒含量要极显著高于酵母胞内的硒含量(P<0.001)。筛选出的诱变菌株M1-5产酒精和M16-28产香能力强的富硒酵母,不仅丰富了菌株资源,也可在后续实验研究中可作为混合发酵菌种用在枸杞酒生产中,对酒发酵缩短发酵时间、提高原料利用率、提高风味物质等方面提供理论参考,对富硒功能性产品的开发和挖掘具有潜在的应用前景。
-
![]()
图 2 不同诱变时间下菌株M1和M16的致死率
Figure 2. Mortality of strains M1 and M16 under different mutagenesis times
![]()
图 3 不同酒精度对菌株生长的影响
Figure 3. Effects of different alcohol content on the growth of strains
![]()
图 4 不同NaCl溶液浓度对菌株生长的影响
Figure 4. Effect of different NaCl solution concentration on the growth of strain
![]()
图 5 不同葡萄糖质量浓度对菌株生长的影响
Figure 5. Effect of different glucose mass concentration on the growth of strain
![]()
图 7 M1-5菌株遗传稳定性测试结果
注:相同字母表示差异不显著(P>0.05)。
Figure 7. Genetic stableility test results of the M1-5 strain
![]()
图 8 M16-28菌株遗传稳定性测试结果
注:相同字母表示差异不显著(P>0.05),不同大小写的字母用以区分不同组间差异。
Figure 8. Genetic stableility test results of the M16-28 strain
![]()
图 9 原始菌及诱变菌株胞内和发酵液的硒含量
注:不同字母表示差异极显著(P<0.001),不同大小写的字母用以区分不同组间差异,***表示各自组内之间差异极显著(P<0.001)。
Figure 9. Selenium content of intracellular and fermentation broth of primitive bacteria and mutagenic strains
表 1 不同酵母菌在富硒培养液中的菌体沉淀量
Table 1 Precipitation amount of different yeasts in selenium-rich medium
菌株编号 硒质量浓度(μg/mL) 0.00 5.00 10.00 20.00 30.00 40.00 50.00 JY-1 **** **** *** ** * * * M **** **** **** *** *** *** *** G2 **** **** **** *** *** *** *** M16 **** **** *** *** ** ** * M19 **** *** ** ** ** ** * G1 **** **** *** * * * * M1 **** **** **** **** *** *** ** M17 **** *** *** ** * * * B7 **** *** *** − − − − 注: “****”表示6/6(每株菌在加硒后菌体沉淀量与初始未加硒菌体沉淀量的比值,以下以此类推);“***”表示5/6,“**”表示4/6;“*”表示3/6,“−”表示无沉淀。  下载: 导出CSV
下载: 导出CSV
表 2 不同酵母菌在富硒培养液中的颜色比较
Table 2 Color comparison of different yeasts in selenium-rich medium
菌株编号 硒质量浓度(μg/mL) 0.00 5.00 10.00 20.00 30.00 40.00 50.00 JY-1 − * ** ** *** *** *** M − * ** *** *** **** **** G2 − * ** *** *** *** *** M16 − * ** ** *** *** *** M19 − * ** *** *** **** **** G1 − * ** ** *** **** **** M1 − − * ** *** *** **** M17 − * ** *** **** **** **** B7 − * ** − − − − 注:“−”代表正常;“*”微红色;“**”粉红色;“***”红色;“****”深红色。
下载: 导出CSV
表 3 M1菌株产酒精能力测定结果
Table 3 Determination results of alcohol production capacity of M1 strains
菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色情况 菌株编号 显色情况 M1 + M1-9 + M1-18 + M1-27 + M1-36 +++ M1-1 ++ M1-10 + M1-19 +++ M1-28 + M1-37 + M1-2 + M1-11 +++ M1-20 ++ M1-29 + M1-38 ++ M1-3 ++ M1-12 ++ M1-21 ++ M1-30 +++ M1-39 ++ M1-4 ++ M1-13 +++ M1-22 ++ M1-31 ++ M1-40 +++ M1-5 +++ M1-14 ++ M1-23 ++ M1-32 ++ M1-41 + M1-6 ++ M1-15 ++ M1-24 +++ M1-33 ++ M1-42 ++ M1-7 + M1-16 ++ M1-25 ++ M1-34 +++ M1-43 ++ M1-8 ++ M1-17 + M1-26 ++ M1-35 + M1-44 ++ 注:“+++”呈红色;“++”呈粉红色;“+”呈浅粉色。
下载: 导出CSV
表 4 M16菌株产酒精能力测定结果
Table 4 Determination results of alcohol production capacity of M16 strains
菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色结果 菌株编号 显色情况 菌株编号 显色情况 M16 +++ M16-12 +++ M16-24 +++ M16-36 +++ M16-48 ++++ M16-1 +++ M16-13 +++ M16-25 +++ M16-37 +++ M16-49 +++ M16-2 +++ M16-14 ++++ M16-26 +++ M16-38 +++ M16-50 +++ M16-3 +++ M16-15 +++ M16-27 +++ M16-39 +++ M16-51 +++ M16-4 +++ M16-16 +++ M16-28 ++++ M16-40 ++++ M16-52 +++ M16-5 +++ M16-17 ++++ M16-29 +++ M16-41 ++++ M16-53 +++ M16-6 +++ M16-18 +++ M16-30 +++ M16-42 ++++ M16-54 +++ M16-7 +++ M16-19 +++ M16-31 +++ M16-43 +++ M16-55 +++ M16-8 +++ M16-20 +++ M16-32 +++ M16-44 ++++ M16-56 +++ M16-9 +++ M16-21 +++ M16-33 +++ M16-45 +++ M16-57 ++ M16-10 +++ M16-22 +++ M16-34 +++ M16-46 +++ M16-58 +++ M16-11 +++ M16-23 +++ M16-35 +++ M16-47 ++++ 注:“++++”呈深红色;“+++”呈红色。
下载: 导出CSV
表 5 M1菌株杜氏小管产气结果
Table 5 Du's tubule gas production result of M1 strain
菌株编号 培养时间(h) 1 2 3 4 5 6 7 M1 − − + ++ ++++ +++++ +++++ M1-5 − + + ++ +++++ +++++ +++++ M1-11 − − + ++ ++++ +++++ +++++ M1-13 −+ + ++ +++ +++++ +++++ +++++ M1-19 − − − + +++ ++++ +++++ M1-24 − − −+ + +++ ++++ +++++ M1-30 − − −+ + +++++ +++++ +++++ M1-34 − − + + +++ ++++ +++++ M1-36 − − − + +++ ++++ +++++ M1-40 −+ + ++ +++ +++++ +++++ +++++ 注:杜氏小管中气体充斥量:“+++++”表示5/5;“++++”表示4/5;“+++”表示3/5;“++”表示2/5;“+”表示1/5;“−+”表示小于1/5;“−”表示无气体产生。
下载: 导出CSV
表 6 M16菌株杜氏小管产气结果
Table 6 Du's tubule gas production result of M16 strain
菌株编号 培养时间(h) 2 4 6 8 10 12 M16 − −+ + ++ +++++ +++++ M16-14 − −+ + ++ +++++ +++++ M16-17 − − + + ++++ +++++ M16-28 − −+ + +++ +++++ +++++ M16-40 − −+ + +++ +++++ +++++ M16-41 −+ −+ + ++ +++++ +++++ M16-42 −+ −+ ++ +++ +++++ +++++ M16-44 −+ −+ +++ ++++ +++++ +++++ M16-47 − − + +++ +++++ +++++ M16-48 − −+ ++ ++++ +++++ +++++ 注:杜氏小管中气体充斥量:“+++++”表示5/5;“++++”表示4/5;“+++”表示3/5;“++”表示2/5;“+”表示1/5;“−+”表示小于1/5;“−”表示无气体产生。
下载: 导出CSV
表 7 酿酒酵母菌株的酒精含量
Table 7 Alcohol content of the Saccharomyces cerevisiae strains
菌株 酒精(%vol) 提升百分比(%) M1 0.657±0.0153c − M1-5 0.907±0.0252a 38.071 M1-13 0.673±0.0306bc 2.838 M1-40 0.703±0.0208b 7.107 注:不同字母表示差异显著(P<0.05)。
下载: 导出CSV
表 8 产香非酿酒酵母菌株总酯、总酸含量
Table 8 Total ester and total acid content of nonSaccharomyces cerevisiae strains
菌株 总酯(g/100 mL) 总酸(g/L) M16 0.98±0.007b 1.125±0.031b M16-28 2.0347±0.041a 1.575±0.045a M16-40 1.05±0.073b 1.575±0.009a M16-44 1.06±0.058b 1.125±0.021b 注:同列数据不同字母表示差异显著(P<0.05),相同字母表示差异不显著(P>0.05)。
下载: 导出CSV
-
[1] ZHU H Q, CHEN S Q, LI R B, et al. Selenium-rich yeast counteracts the inhibitory effect of nanoaluminum on the formation of porcine neutrophil extracellular traps[J]. Research in Veterinary Science,2023,161:138−144. doi: 10.1016/j.rvsc.2023.06.018
[2] CAO C Y, LI X W, QIN L, et al. High Selenium Yeast mitigates aluminum-induced cerebral inflammation by increasing oxidative stress and blocking NO production[J]. Biometals,2018,31(5):835−843. doi: 10.1007/s10534-018-0128-0
[3] 涂青, 杨双全, 章之柱, 等. 富硒酵母发酵工艺的优化[J]. 中国酿造,2022,41(10):140−145. [TU Q, YANG S Q, ZHANG Z Z, et al. Optimization of fermentation process of selenium-rich yeast[J]. China Brewing,2022,41(10):140−145.] TU Q, YANG S Q, ZHANG Z Z, et al. Optimization of fermentation process of selenium-rich yeast[J]. China Brewing, 2022, 41(10): 140−145.
[4] SUN N, DANG H, ZHANG Y Y, et al. Inorganic Selenium Transformation into Organic Selenium by Monascus purpureus[J]. Foods,2023,12(18):3375. doi: 10.3390/foods12183375
[5] LIAO X L, RAO S, YU T, et al. Selenium yeast promoted the Se accumulation, nutrient quality and antioxidant system of cabbage (Brassica oleracea var. capitata L.)[J]. Plant Signal Behav,2021,16(6):1907042. doi: 10.1080/15592324.2021.1907042
[6] 王凤梅, 张邦建, 岳泰新. 内蒙古西部地区野生葡萄酒相关酵母发酵及产酯能力分析[J]. 南方农业学报,2019,50(4):825−830. [WANG F M, ZHANG B J, YUE T X. The ability of fermentation and esters production of wild winemaking yeasts isolated from the western district of Inner Mongolia[J]. Acta Agriculturae Sinica of Southern China,2019,50(4):825−830.] doi: 10.3969/j.issn.2095-1191.2019.04.20 WANG F M, ZHANG B J, YUE T X. The ability of fermentation and esters production of wild winemaking yeasts isolated from the western district of Inner Mongolia[J]. Acta Agriculturae Sinica of Southern China, 2019, 50(4): 825−830. doi: 10.3969/j.issn.2095-1191.2019.04.20
[7] 郑超群. 基于酿酒酵母和异常汉逊酵母混合发酵技术的黄酒生产工艺及风味成分研究[D]. 杭州:浙江工商大学, 2023. [ZHENG C Q. Research on the production process and flavor components of rice wine based on the mixed fermentation technology of Saccharomyces cerevisiae and Hansenula anomalies[D].Hangzhou: Zhejiang Gongshang University, 2023.] ZHENG C Q. Research on the production process and flavor components of rice wine based on the mixed fermentation technology of Saccharomyces cerevisiae and Hansenula anomalies[D].Hangzhou: Zhejiang Gongshang University, 2023.
[8] 王静尧. 高产酯非酿酒酵母筛选及在混菌发酵中的发酵特性研究[D]. 阿拉尔:塔里木大学, 2023. [WANG J Y. Screening of non-Saccharomyces cerevisiae with high ester yield and its fermentation characteristics in mixed fermentation[D]. Alaer:Tarim University, 2023.] WANG J Y. Screening of non-Saccharomyces cerevisiae with high ester yield and its fermentation characteristics in mixed fermentation[D]. Alaer: Tarim University, 2023.
[9] 李秀婷, 范光森, 孙宝国, 等. 产己酸乙酯的葡萄牙棒孢酵母菌株及其培养方法与应用:中国,111909861A[P]. 2020-11-10. [LI X T, FAN G S, SUN B G, et al. Strain of Clavispora lusitaniae producing ethyl caproate and its culture method and application:China, 111909861A[P]. 2020-11-10.] LI X T, FAN G S, SUN B G, et al. Strain of Clavispora lusitaniae producing ethyl caproate and its culture method and application: China, 111909861A[P]. 2020-11-10.
[10] LIU Y, CHEN X, WEI D, et al. Breeding a novel chlorophyll-deficient mutant of Auxenochlorella pyrenoidosa for high-quality protein production by atmospheric room temperature plasma mutagenesis[J]. Bioresour Technol,2023,390:129907. doi: 10.1016/j.biortech.2023.129907
[11] ZHANG Q, MIAO R Y, FENG R C, et al. Application of Atmospheric and room-temperature plasma (ARTP) to microbial breeding[J]. Current Issues in Molecular Biology,2023,45(8):6466−6484. doi: 10.3390/cimb45080408
[12] 汪锡武, 冯婉婷, 方旭娟, 等. 富硒啤酒酵母的选育及有机硒含量的测定[J]. 食品与发酵工业,2024,50(10):48−54. [WANG X W, FENG W T, FANG X J, et al. Breeding of selenium-rich beer yeast and determination of organic selenium content[J]. Food and Fermentation Industry,2024,50(10):48−54.] WANG X W, FENG W T, FANG X J, et al. Breeding of selenium-rich beer yeast and determination of organic selenium content[J]. Food and Fermentation Industry, 2024, 50(10): 48−54.
[13] 林仙菊, 张祝兰, 严凌斌, 等. ARTP诱变选育环孢菌素A高产菌株及发酵工艺优化[J]. 工业微生物,2023,53(4):158−163. [LIN X J, ZHANG Z L, YAN L B, et al. Study on screening of high-yield cyclosporin A producing strain by atmospheric and room temperature plasma mutagenesis and optimization of fermentation conditions[J]. Industrial Microbiology,2023,53(4):158−163.] doi: 10.3969/j.issn.1001-6678.2023.04.044 LIN X J, ZHANG Z L, YAN L B, et al. Study on screening of high-yield cyclosporin A producing strain by atmospheric and room temperature plasma mutagenesis and optimization of fermentation conditions[J]. Industrial Microbiology, 2023, 53(4): 158−163. doi: 10.3969/j.issn.1001-6678.2023.04.044
[14] HONG K Q, FU X M, LEI F F, et al. Selection of salt-tolerance and ester-producing mutant Saccharomyces cerevisiae to improve flavour formation of soy sauce during co-fermentation with torulopsis globosa[J]. Foods,2023,12(18):3449. doi: 10.3390/foods12183449
[15] 杨新, 陈莉, 杨双全, 等. 开阳地区桑葚果园富硒酵母菌的分离、筛选及鉴定[J]. 食品与发酵工业,2020,46(24):66−73. [YANG X, CHEN L, YANG S Q, et al. Isolation, screening and identification of selenium-enrichedyeast from mulberry orchard of Kaiyang[J]. Food and Fermentation Industry,2020,46(24):66−73.] YANG X, CHEN L, YANG S Q, et al. Isolation, screening and identification of selenium-enrichedyeast from mulberry orchard of Kaiyang[J]. Food and Fermentation Industry, 2020, 46(24): 66−73.
[16] 刘程, 梁静一美, 王周利, 等. 茯砖茶中冠突散囊菌的分离鉴定及富硒菌株筛选[J]. 西北农业学报,2022,31(10):1344−1356. [LIU C, LIANG JING Y M, WANG Z L, et al. Isolation and identification of eurotium cristatum in Fucha tea and screening of selenium-rich strains[J]. Journal of Northwest Agriculture,2022,31(10):1344−1356.] LIU C, LIANG JING Y M, WANG Z L, et al. Isolation and identification of eurotium cristatum in Fucha tea and screening of selenium-rich strains[J]. Journal of Northwest Agriculture, 2022, 31(10): 1344−1356.
[17] 胡惠影. 乳杆菌(Lactobacillus sp.)JZ07富集转化硒的特性研究[D]. 郑州:河南工业大学, 2018. [HU H Y. Characteristics of Selenium accumulation and transformation by Lactobacillus sp. JZ07[D]. Zhengzhou:Henan University of Technology, 2018.] HU H Y. Characteristics of Selenium accumulation and transformation by Lactobacillus sp. JZ07[D]. Zhengzhou: Henan University of Technology, 2018.
[18] 中华人民共和国国家监督检验检疫总局. GB/T 5009.93-2017食品安全国家标准 食品中硒的测定[S]. 北京:中国标准出版社,2017. [General Administration of Supervision, Inspection and Quarantine of the People's Republic of China. Determination of selenium in GB/T 5009.93-2017 National standard for food safety. Determination of selenium in food[S]. Beijing:China Standards Press,2017.] General Administration of Supervision, Inspection and Quarantine of the People's Republic of China. Determination of selenium in GB/T 5009.93-2017 National standard for food safety. Determination of selenium in food[S]. Beijing: China Standards Press, 2017.
[19] 金海炎. 猕猴桃酒优良酿酒菌株的选育及混菌发酵工艺优化研究[D].南阳: 南阳师范学院, 2023. [JIN H Y. Study on yeast screening of kiwifruit wine and optimization of its fermentation conditions for compound barm[D]. Nanyang:Nanyang Normal University, 2023.] JIN H Y. Study on yeast screening of kiwifruit wine and optimization of its fermentation conditions for compound barm[D]. Nanyang: Nanyang Normal University, 2023.
[20] 王伟雄. 基于Kluyveromyces marxianus与酿酒酵母混合发酵葡萄酒动力学分析及质量提升研究[D]. 乌鲁木齐:新疆农业大学,2022. [WANG W X. DynamicAnalysis and Quality Improvement of Mixed Fermentation Wine Based on Kluyveromyces Marxianus and Saccharomyces cerevisiae[D]. Urumqi:Xinjiang Agricultural University,2022.] WANG W X. DynamicAnalysis and Quality Improvement of Mixed Fermentation Wine Based on Kluyveromyces Marxianus and Saccharomyces cerevisiae[D]. Urumqi: Xinjiang Agricultural University, 2022.
[21] 中华人民共和国国家监督检验检疫总局. GB/T 5009.225-2016食品安全国家标准 酒中乙醇浓度的测定[S]. 北京:中国标准出版社,2016. [General Administration of Supervision, Inspection and Quarantine of the People's Republic of China. GB/T 5009.225-2016 National standard for food safety. Determination of ethanol concentration in alcohol[S]. Beijing:China Standard Press,2016.] General Administration of Supervision, Inspection and Quarantine of the People's Republic of China. GB/T 5009.225-2016 National standard for food safety. Determination of ethanol concentration in alcohol[S]. Beijing: China Standard Press, 2016.
[22] 凌思雨, 王洲, 张会敏, 等. 常压室温等离子体诱变与微生物微液滴培养选育谷胱甘肽高产菌株[J]. 食品科学,2023,44(4):200−208. [LING S Y, WANG Z, ZHANG H M, et al. Breeding of a high-yield glutathione-producing strain by atmospheric and room temperature plasma mutagenesis and microbial microdroplet culture system[J]. Food Science,2023,44(4):200−208.] LING S Y, WANG Z, ZHANG H M, et al. Breeding of a high-yield glutathione-producing strain by atmospheric and room temperature plasma mutagenesis and microbial microdroplet culture system[J]. Food Science, 2023, 44(4): 200−208.
[23] 杨昳津, 俞剑燊, 夏永军, 等. 优良黄酒酵母的筛选及其抗逆性能分析[J]. 现代食品科技,2015,31(8):261−267. [YANG Y J, YU J S, XIA Y J, et al. Screening for Quality Strains of Yeast from Yellow Rice Wine and Stress Resistance Analysis[J]. Modern Food Science and Technology,2015,31(8):261−267.] YANG Y J, YU J S, XIA Y J, et al. Screening for Quality Strains of Yeast from Yellow Rice Wine and Stress Resistance Analysis[J]. Modern Food Science and Technology, 2015, 31(8): 261−267.
[24] LI Y C, RAO J W, MENG F B, et al. Combination of mutagenesis and adaptive evolution to engineer salt-tolerant and aroma-producing yeast for soy sauce fermentation[J]. Journal of the Science of Food and Agriculture,2021,101(10):4288−4297. doi: 10.1002/jsfa.11068
[25] TIAN T T, WU D H, CHAN-TAT NG, et al. A multiple-step strategy for screening Saccharomyces cerevisiae strains with improved acid tolerance and aroma profiles[J]. Applied Microbiology and Biotechnology,2020,104(7):3097−3107. doi: 10.1007/s00253-020-10451-z
[26] LIU C F, LI Q, NIU C D, et al. The use of atmospheric and room temperature plasma mutagenesis to create a brewing yeast with reduced acetaldehyde production[J]. Journal of the Institute of Brewing,2018,124:236−243. doi: 10.1002/jib.498
[27] WEI F Q, LI M L, WANG M, et al. A C6/C5 co-fermenting Saccharomyces cerevisiae strain with the alleviation of antagonism between xylose utilization and robustness[J]. GCB Bioenergy,2020,13:83−97.
[28] 王犁烨, 王浩臣, 马珊, 等. 常压室温等离子体选育高产酒精及酸的酿酒酵母[J]. 食品与机械,2019,35(5):26−31. [WANG L Y, WANG H C, MA S, et al. Breeding of acid-tolerant Saccharomyces cerevisiae strains by ARTP mutation[J]. Food and Machinery,2019,35(5):26−31.] WANG L Y, WANG H C, MA S, et al. Breeding of acid-tolerant Saccharomyces cerevisiae strains by ARTP mutation[J]. Food and Machinery, 2019, 35(5): 26−31.
[29] 蔡飞, 高飞飞, 王斌, 等. 富硒酵母的分离鉴定及富硒条件的优化[J]. 中国酿造,2018,37(6):103−108. [CAI F, GAO F F, WANG B, et al. Isolation and identification of selenium-enriched yeast and optimization of selenium-enriched conditions[J]. China Brewing,2018,37(6):103−108.] CAI F, GAO F F, WANG B, et al. Isolation and identification of selenium-enriched yeast and optimization of selenium-enriched conditions[J]. China Brewing, 2018, 37(6): 103−108.
[30] 凡敏, 黄珍, 李欣, 等. 富硒酵母的筛选鉴定及其作为鱼饲料的应用研究[J]. 湖北农业科学,2017,56(8):1416−1419. [FAN M, HUANG Z, LI X, et al. Isolation, identification and application research of fish feed of Selenium-rich yeast[J]. Hubei Agricultural Sciences,2017,56(8):1416−1419.] FAN M, HUANG Z, LI X, et al. Isolation, identification and application research of fish feed of Selenium-rich yeast[J]. Hubei Agricultural Sciences, 2017, 56(8): 1416−1419.
-
期刊类型引用(0)
其他类型引用(2)
 下载:
下载:











计量
- 文章访问数: 72
- HTML全文浏览量: 9
- PDF下载量: 13
- 被引次数: 2
