


Promoting Proliferation of Streptococcus thermophilus FUA329 and Antioxidant Activity of Enzyme Hydrolysis Products of Larimichthys polyactis
-
摘要: 本研究旨在探索小黄鱼酶解产物对嗜热链球菌FUA329的增殖作用,并测定不同分子量酶解产物的抗氧化活性。在选择不同酶水解小黄鱼加工副产物的基础上,选用木瓜蛋白酶对小黄鱼副产物进行酶解。以酶解产物对嗜热链球菌的增殖效应为评价指标,采用单因素实验和响应面法优化酶解条件,并对酶解产物的体外抗氧化活性进行测定。结果表明,小黄鱼加工副产物酶解的最佳工艺参数为:酶解时间5 h,料液比1:5.1(w/v),加酶量2.8%。将优化条件下制备的小黄鱼酶解液按分子量大小分为4个片段:>10000 Da,5000~10000 Da,3000~5000 Da,<3000 Da,其中分子量<3000 Da的片段促进嗜热链球菌的增殖能力最强。小黄鱼酶解产物对于DPPH、羟自由基和超氧阴离子自由基的清除率分别达到68.75%、56.23%和30.64%。小黄鱼酶解产物对于嗜热链球菌的增殖有一定的促进效果,同时具有一定的抗氧化能力,有开发为嗜热链球菌氮源的潜力。Abstract: To determine the promoting effects of enzyme hydrolysis products of Larimichthys polyactis on the proliferation of Streptococcus thermophilus FUA329. The in vitro antioxidant activity of the enzymatic hydrolysis with different molecular weights were determined. Papain was selected for the enzymatic hydrolysis of its by-products on the basis of selecting different enzymes to hydrolyze the processing by-products of Larimichthys polyactis. Taking the proliferative effect of enzymatic degradation products on Streptococcus thermophilus as an indicator of evaluation. The enzymatic hydrolysis conditions were optimized through single factor and response surface methodology. The in vitro antioxidant activity of the hydrolysis products of the by-products of Larimichthys polyactis (HPBL) were determined. The results showed that the best enzymatic hydrolysis conditions included the hydrolysis time, material-liquid ratio and enzyme loading were 5 h, 1:5.1 (w/v) and 2.8% (w/v), respectively. The HPBL prepared under the optimized conditions was separated by ultrafiltration into four fragments, according to the molecular weight: >10000 Da, 5000~10000 Da, 3000~5000 Da, <3000 Da, among which the fragment with molecular weight of less than 3000 Da promoted the proliferation of Streptococcus thermophilus significantly. The scavenging rate of HPBL for DPPH, hydroxyl radicals and superoxide anion radicals reached 68.75%, 56.23% and 30.64%, respectively. The enzyme hydrolysis products of Larimichthys polyactis significantly promoted the proliferation of Streptococcus thermophilus FUA329 and showed antioxidant activity. HPBL would have a certain promoting effect on the proliferation of Streptococcus thermophilus FUA329, and a certain antioxidant capacity, which would have the potential to be developed as a nitrogen source for Streptococcus thermophilus.
-
小黄鱼(Larimichthys polyactis)栖身于沿岸及近海沙泥底中底层水域,是我国东黄海区重要的经济鱼种[1],常被加工成鱼干、鱼豆腐、鱼丸等产品,其头部和内脏为主要的加工副产物[2]。随着海产品消费量的快速增长,其加工副产品也逐年增多,这些加工副产物的随意丢弃和掩埋造成了资源浪费和严重的环境污染[3]。为了妥善地处理这些加工副产物,提高海产品加工附加值,获得良好的经济和社会效益,王婧[4]利用生物酶解技术结合热反应技术制备水产调味料水产调味品,许韬等[5]采用稀碱水解法、酶解法和微波辅助法提取鱼油并以棕榈酸为指标,研究晶体传质模型以及低温富集内脏油中棕榈酸的条件,刘冲[6]以小黄鱼鱼头为主要原料开发风味鱼糕和小黄鱼风味休闲脆片。这些研究丰富了小黄鱼副产物的处理方式,但对小黄鱼副产物中含有的丰富蛋白资源未进行深入挖掘,在开发其更多的用途方面存在一定局限性。
随着健康消费意识的升级,功能食品市场规模不断扩大,益生菌因其具有多种保健功能而备受青睐[7]。嗜热链球菌(Streptococcus thermophilus)作为一种多功能益生菌,能够改善人体肠道微生态[8]、抗癌[9]、调节血压[10]、缩短酸奶的凝乳时间[11]。但嗜热链球菌普遍对生长条件的要求比较苛刻,尤其是其蛋白质代谢能力较弱,影响了嗜热链球菌的进一步开发利用。因此,挖掘有效促进菌体增殖的物质受到研究者的广泛关注[12]。多种海产品的加工副产物酶解后可以作为氮源促进益生菌生长[13],尤其是一些鱼类加工副产物[14−15]。大西洋鳕鱼(Gadus morhua L.)内脏水解产物作为氮源可促进植物乳杆菌、大肠杆菌、酿酒酵母以及乳酸菌的生长,与商用氮源效果相当或略低[16−17]。Safati等[18]对黄鳍金枪鱼(Thunnus albacares)头部进行酶解获得鱼肽,用10 g/L的鱼肽取代18 g/L肉类和酪蛋白肽,乳酸菌的最大增长率和比增长率较高,但略低于商用的MRS氮源。此外,沙丁鱼(Sardina pilchardus)副产物酶解后也可作为大肠杆菌和酿酒酵母生长的良好氮源,能够显著降低益生菌的生长成本[19-20]。
源于人母乳的嗜热链球菌FUA329能将人体难吸收利用的鞣花酸转化为人体易吸收利用且具有缓解肌肉衰退[21]、抗癌[22]、抗炎[23]、抗衰老[24]等生物活性的尿石素A,具有开发为益生菌的潜力[25−26]。但是,嗜热链球菌FUA329生长较为缓慢。因此,本研究选取小黄鱼加工副产物为原料,选择最适蛋白酶对其进行酶解,研究HPBL对嗜热链球菌FUA329增殖的影响,利用单因素实验及响应面试验对酶解条件进行优化,并对HPBL体外抗氧化活性进行测定。本研究结果为获得促进嗜热链球菌FUA329增长的氮源奠定实验基础,并为小黄鱼的高值化利用提供新思路。
1. 材料与方法
1.1 材料与仪器
冷冻小黄鱼(Larimichthys polyactis) 购自连云港市海鲜蔬菜直销市场;嗜热链球菌Streptococcus thermophilus FUA329 本实验室分离保存;厌氧菌基础肉汤(干粉) 美国赛默飞世尔科技公司;木瓜蛋白酶(2000 U/mg)、胰蛋白酶(250 U/mg)、碱性蛋白酶(200 U/mg)、中性蛋白酶(500 U/mg)、胃蛋白酶(3000 U/mg) 上海阿拉丁试剂有限公司;DPPH自由基清除能力试剂盒、抑制与产生超氧阴离子自由基测定试剂盒、羟自由基测定试剂盒 南京建成有限公司;BCA蛋白浓度测定试剂盒 北京索莱宝科技有限公司;本研究中使用的化学试剂均为分析纯。
SHA-B数显水浴恒温振荡器(双功能) 江苏金坛亿通电子有限公司;YXQ-LS-50S立式高压蒸汽灭菌器 上海博讯实业有限公司医疗设备厂;DPH-9082电热恒温培养箱 上海博珍仪器设备制造厂;FE20 Plust pH计 梅特勒-托利多仪器(上海)有限公司;SQP电子天平 赛多利斯科学仪器(北京)有限公司;Multifuge X 3R台式高速冷冻离心机、Multiskan SkyHigh全波长酶标仪 美国赛默飞世尔科技公司。
1.2 实验方法
1.2.1 小黄鱼蛋白含量的测定
将小黄鱼解冻后,去除肉、骨(保留头和内脏),以1:1的比例加水,放入均质机破碎,收集破碎后液体,按试剂盒说明书操作[27]测定其蛋白质含量。
1.2.2 小黄鱼酶解液的制备
小黄鱼解冻→去除可食部分(保留头和内脏)→加入一定比例的水→均质机破碎→调节pH→加入蛋白酶酶解→沸水浴加热酶解液(10 min)→离心(12000 r/min,10 min)→撇去浮油(去部分脂肪)和沉淀→过滤→小黄鱼酶解液。
1.2.3 酶的选择
在木瓜蛋白酶、胰蛋白酶、碱性蛋白酶、中性蛋白酶、胃蛋白酶这五种酶中进行选择,分别在各自最适pH和最适温度下加入3%(w/v)的酶制剂反应4 h,以其对嗜热链球菌FUA329增殖效果(ΔOD600)为评价指标,确定最佳酶。
1.2.4 单因素实验
鉴于木瓜蛋白酶酶解的最适温度为50~55 ℃,最适pH为5.0~7.0,本实验以料液比、加酶量和酶解时间3个单因素为考察对象,测定其对嗜热链球菌FUA329增殖促进作用的影响。料液比、加酶量、酶解时间分别设计为1:5(w/v)、3%(w/v)和5 h其中2个单因素固定,其他因素水平如下:料液比:1:1、1:5、1:10、1:15;加酶量:1%、2%、3%、4%、5%;酶解时间:1、2、3、4、5、6、7、8 h。
1.2.5 响应面试验
利用Design-Expert 8.0.6软件,以嗜热链球菌FUA329的增殖效果为响应值,选取料液比(A)、加酶量(B)和酶解时间(C)3个参数作为自变量,ΔOD600(Y)作为因变量,通过三因素三水平响应面试验优化酶解工艺,响应面自变量因素与水平见表1。
表 1 响应面自变量因素与水平Table 1. Response surface independent variable factors and levels因素 水平 −1 0 1 A料液比(w/v) 1:2 1:5 1:8 B加酶量(%) 2 3 4 C酶解时间(h) 4 5 6 1.2.6 不同分子量小黄鱼酶解产物的制备
利用响应面试验得到的最优条件制备酶解液,分别用分子量为10000、5000和3000 Da的超滤管4000×g,30 min分离,分别采集了HPBL Ⅰ(>10000 Da)、HPBL Ⅱ(5000~10000 Da)、HPBL Ⅲ(3000~5000 Da)和HPBL Ⅳ(<3000 Da)4种成分。将各组分冷冻干燥,置于−20 ℃保存备用。
1.2.7 小黄鱼酶解产物对嗜热链球菌FUA329增殖促进作用的测定
本试验以宋益善等[14]选择的MRS化学限定培养基为基础,根据嗜热链球菌FUA329的特性进行调整,最终的MRS化学限定培养基成分如下:葡萄糖 20、乙酸钠 5、NaCl 5、柠檬酸二胺 2、K2HPO4·3H2O 2、KH2PO4 2、吐温80 1、L-精氨酸 1、丙酮酸钠 1、MgSO4·7H2O 0.2、MnSO4·7H2O 0.05、肌醇 0.02、核黄素 0.02、吡哆醇 0.02、烟酸 0.02、FeSO4 0.02、生物素 0.01、硫胺素 0.01、钴胺素 0.01、氯化血红素 0.005、维生素K1 0.005和叶酸 0.005(g/L)。维生素和无机盐需用0.22 μm滤膜过滤除菌,配制成溶液后121 ℃高压灭菌30 min。
将甘油保藏菌以1%(v/v)的接种量接入10 mL MRS化学限定液体培养基中,37 ℃厌氧培养24 h后,以1%(v/v)的接种量置于含有HPBL和不含有HPBL的MRS化学限定培养基中,37 ℃厌氧培养。培养48 h之后,分别使用酶标仪测定其在600 nm处的吸光度值,菌株增殖量计算方式如下:
注:ΔOD600为嗜热链球菌FUA329的增殖量;OD600a为嗜热链球菌FUA329在含有HPBL的化学限定培养液中的吸光值;OD600b为嗜热链球菌FUA329在不含HPBL的化学限定培养中的吸光值[15]。
1.2.8 抗氧化活性测定
DPPH自由基清除能力、羟自由基清除能力和超氧阴离子清除能力采用分光光度计法测定,按试剂盒说明书操作。
1.3 数据处理
所有试验均重复3次。作图使用Origin 2019软件,并对试验结果进行整理和分析。利用Design-Expert 8.0.6分析软件进行回归拟合,并进行统计学分析,P<0.05表示结果有统计学意义。
2. 结果与分析
2.1 蛋白含量的测定结果
冷冻小黄鱼均质后的液体蛋白含量为20.41%±0.36%,此可方便后续加酶量的选择。
2.2 酶的选择
用胃蛋白酶、中性蛋白酶、木瓜蛋白酶、胰蛋白酶以及碱性蛋白酶水解小黄鱼加工副产物,结果表明木瓜蛋白酶的酶解产物促进嗜热球菌增殖作用显著优于其它几种酶类(图1)(P<0.05)。丁慧璞等[28]报道木瓜蛋白酶对小黄鱼副产物的酶解效果最好,与本实验结果一致。可能的原因是水解度高的酶解产物中分子量小的肽段含量较多,这些肽段对于嗜热链球菌FUA329有促生长作用。因此,选择木瓜蛋白酶进行后续研究。
![]()
2.3 小黄鱼酶解产物促进嗜热链球菌增殖的单因素实验结果
2.3.1 料液比对嗜热链球菌FUA329增殖的影响
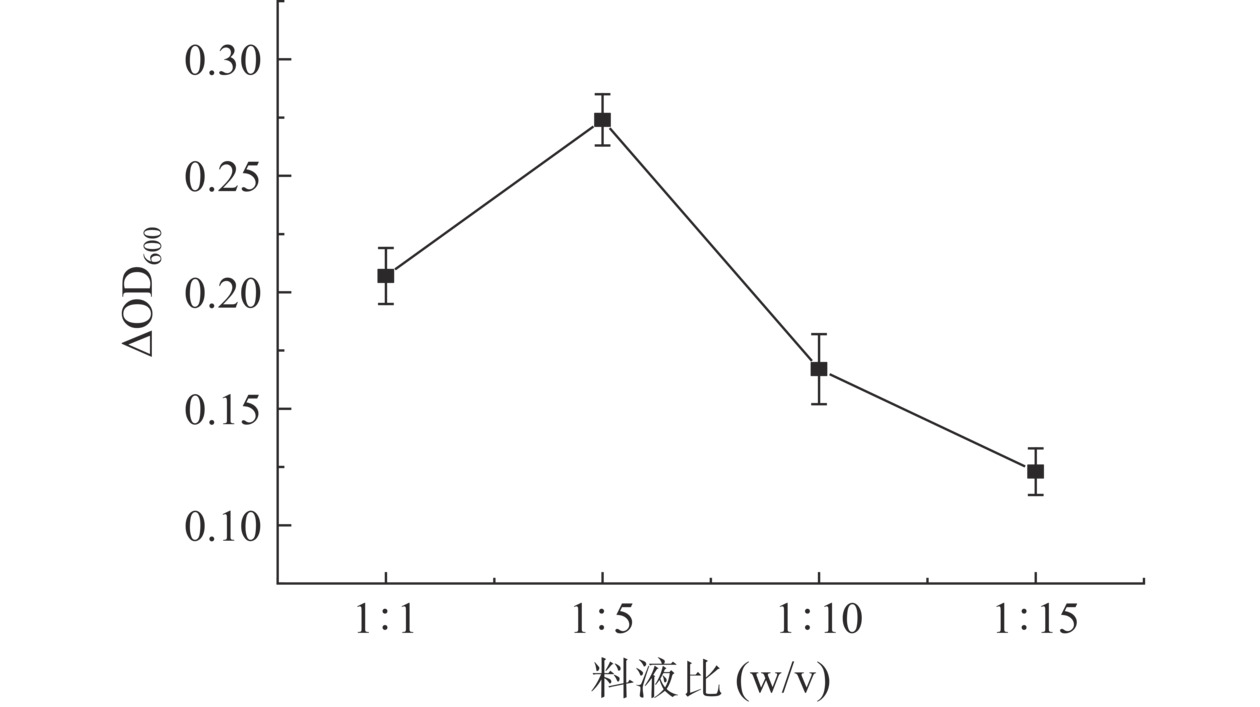
随着料液比的降低,ΔOD600值先增大后减小,当料液比为1:5时,ΔOD600值最高(图2)。当料液比处在较高的状态时,木瓜蛋白酶与小黄鱼副产物的有效接触面积较小,酶解效果差,随着水量的增加,有效接触面积逐渐变大,促进寡肽的生成;随着水量的进一步增加,酶和鱼皮蛋白在水中的浓度降低,不利于酶促反应,降低了寡肽的生成率,因此可供嗜热链球菌FUA329作为有效氮源的肽段相对较少,从而导致促进嗜热链球菌FUA329的增殖效果较差。
![]() 图 2 料液比对嗜热链球菌FUA329增殖的影响Figure 2. Effects of material-liquid ratio on proliferation of Streptococcus thermophilus FUA329
图 2 料液比对嗜热链球菌FUA329增殖的影响Figure 2. Effects of material-liquid ratio on proliferation of Streptococcus thermophilus FUA3292.3.2 加酶量对嗜热链球菌FUA329增殖的影响
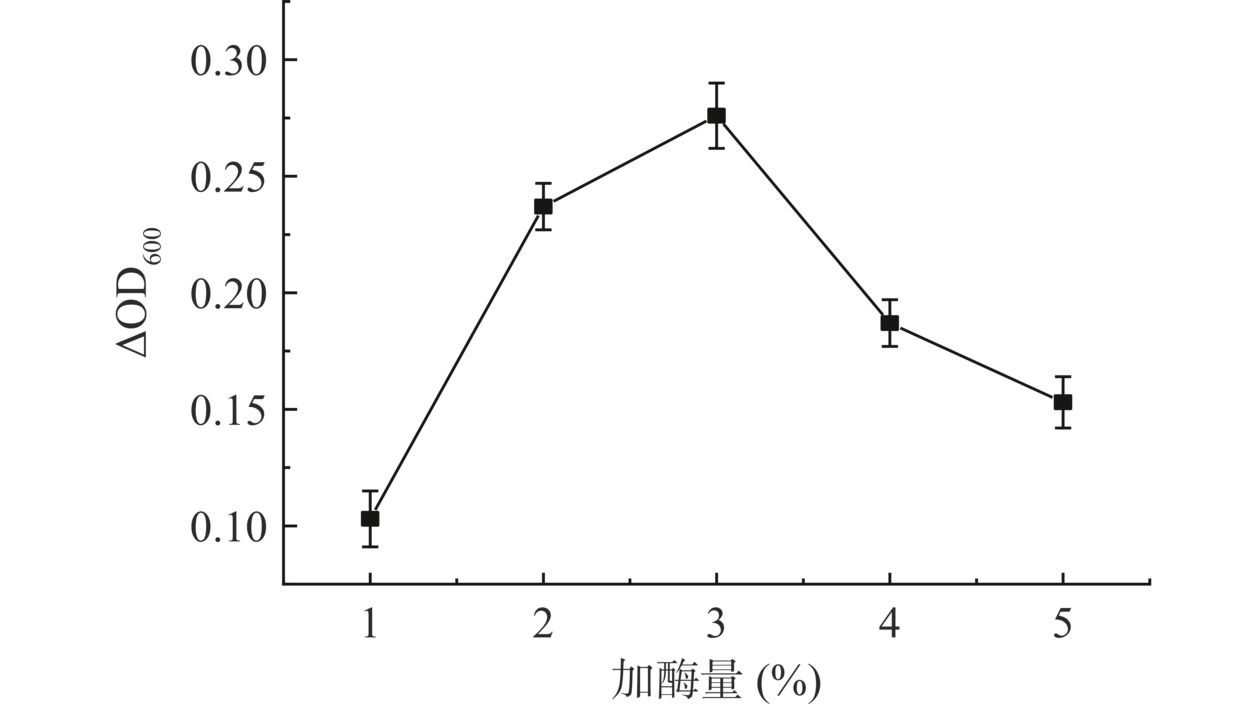
随着加酶量的增加,嗜热链球菌FUA329的ΔOD600值呈现先升高后降低的趋势(图3),且加酶量为3%时ΔOD600值最大。加酶量较少时,酶解小黄鱼副产物生成的促进嗜热链球菌FUA329增殖的寡肽较少,在低加酶量内,加酶量的增加可促进鱼皮蛋白水解生成寡肽;当加酶量达到3%时,水解小黄鱼副产物产生的促进嗜热链球菌FUA329增殖的肽段最多。进一步增加酶量,肽段被水解为增殖效果远不如寡肽的氨基酸,最终导致嗜热链球菌FUA329的ΔOD600值下降。陈建康等[29]研究草鱼皮酶解产物促进嗜热链球菌增殖作用时发现加酶量3.0%对应的OD600值略高于加酶量2.0%对应的OD600值,与本实验结果一致。可能的原因是随着加酶量的进一步增加,二者寡肽处于相同较低水平时,氨基酸促进嗜热链球菌增殖的作用逐步显现。
![]() 图 3 加酶量对嗜热链球菌FUA329增殖的影响Figure 3. Effects of enzyme loading on proliferation of Streptococcus thermophilus FUA329
图 3 加酶量对嗜热链球菌FUA329增殖的影响Figure 3. Effects of enzyme loading on proliferation of Streptococcus thermophilus FUA3292.3.3 酶解时间对嗜热链球菌FUA329增殖的影响
如图4所示,随着反应时间的增加,ΔOD600值先增大后减小,在酶解时间为5 h时达到最大值。Juillard等[30]的研究表明,嗜热链球菌更倾向于利用环境中的寡肽进行增殖,而非体系中的游离氨基酸。在酶促反应早期,随着酶解时间的增加,反应体系中不断产生分子量小的肽段,其增殖效果也越来越好,并在5 h时达到最高值;之后,由于底物的不断减少,这些已经生成的寡肽又被反应体系中的木瓜蛋白酶继续水解成为氨基酸,使得促进嗜热链球菌FUA329增值的效果变差。
![]() 图 4 酶解时间对嗜热链球菌FUA329增殖的影响Figure 4. Effects of hydrolysis time on proliferation of Streptococcus thermophilus FUA329
图 4 酶解时间对嗜热链球菌FUA329增殖的影响Figure 4. Effects of hydrolysis time on proliferation of Streptococcus thermophilus FUA3292.4 促进嗜热链球菌FUA329增殖的响应面优化
2.4.1 响应面试验设计与结果
基于小黄鱼酶解产物促进嗜热链球菌FUA329增殖的单因素实验结果,采用Box-Behnken中心组合试验对小黄鱼副产物的酶解工艺条件进行设计优化。响应面试验设计与结果如表2所示。
表 2 响应面试验设计与结果Table 2. Design and results of response surface experimental实验号 料液比 (A) 加酶量(B) 酶解时间 (C) ΔOD600 (Y) 1 0 1 −1 0.206±0.004 2 0 0 0 0.283±0.004 3 0 0 0 0.278±0.001 4 0 −1 1 0.238±0.005 5 0 −1 −1 0.226±0.002 6 1 0 1 0.242±0.003 7 −1 −1 0 0.193±0.002 8 0 1 1 0.189±0.002 9 1 −1 0 0.224±0.004 10 1 1 0 0.141±0.002 11 −1 1 0 0.191±0.004 12 0 0 0 0.288±0.002 13 −1 0 1 0.236±0.002 14 0 0 0 0.279±0.001 15 0 0 0 0.283±0.007 16 −1 0 −1 0.237±0.006 17 0 0 −1 0.244±0.004 2.4.2 回归模型和方差分析
以嗜热链球菌FUA329的增殖效果ΔOD600为响应值,选择液料比、加酶量和酶解时间3个因素作为自变量进行优化,利用Design-Expert 8.0.6软件进行多元回归分析,拟合后获得ΔOD600随料液比(A)、加酶量(B)和酶解时间(C)变化的二次多项回归方程为下式:
ΔOD600=0.280−7.500×10−4×A−0.019×B−0.001×C−0.020×A×B−2.500×10−4×A×C−7.250×10−4×B×C−0.035×A2−0.060×B2−7.475×10−3×C2
方差分析结果见表3。该模型的F值为93.35,且该模型P<0.0001,表明所得到的数学模型极显著,失拟项P值为0.1336>0.05,表明未知因素对试验结果的影响较小,残差主要源于随机误差;R2=0.9917,说明该模型能够解释99.17%的响应值变化,拟合程度良好,可以用于酶解小黄鱼副产物对嗜热链球菌FUA329的增殖作用进行分析和预测。
表 3 回归模型和方差分析Table 3. Analysis of variance of the regression model方差来源 平方和 自由度 均方 F值 P值 显著性 模型 0.027 9 2.978×10−3 93.35 < 0.0001 ** A 4.500×10−6 1 4.500×10−6 0.14 0.7183 B 2.965×10−3 1 2.965×10−3 92.93 < 0.0001 ** C 8.000×10−6 1 8.000×10−6 0.25 0.6319 AB 1.640×10−3 1 1.640×10−3 51.42 0.0002 ** AC 2.500×10−7 1 2.500×10−7 7.837×10−3 0.9319 BC 2.102×10−4 1 2.102×10−4 6.59 0.0372 * A2 5.151×10−3 1 5.151×10−3 161.46 < 0.0001 ** B2 0.015 1 0.015 474.77 < 0.0001 ** C2 2.353×10−4 1 2.353×10−4 7.38 0.0300 * 残差 2.233×10−4 7 3.190×10−5 失拟误差 1.605×10−4 3 5.350×10−5 3.41 0.1336 不显著 纯误差 6.280×10−5 4 1.570×10−5 总离差 0.027 16 注:*P<0.05表示显著,**P<0.001表示极显著。 回归方程的显著性检验表明模型一次项B,交互项AB,二次项A2、B2对ΔOD600的影响极显著(P<0.01);模型交互项BC,二次项C2显著影响ΔOD600(P<0.05)。F值结果表明3个因素对嗜热链球菌FUA329增殖的影响顺序为:加酶量>酶解时间>料液比。
2.4.3 响应面交互分析
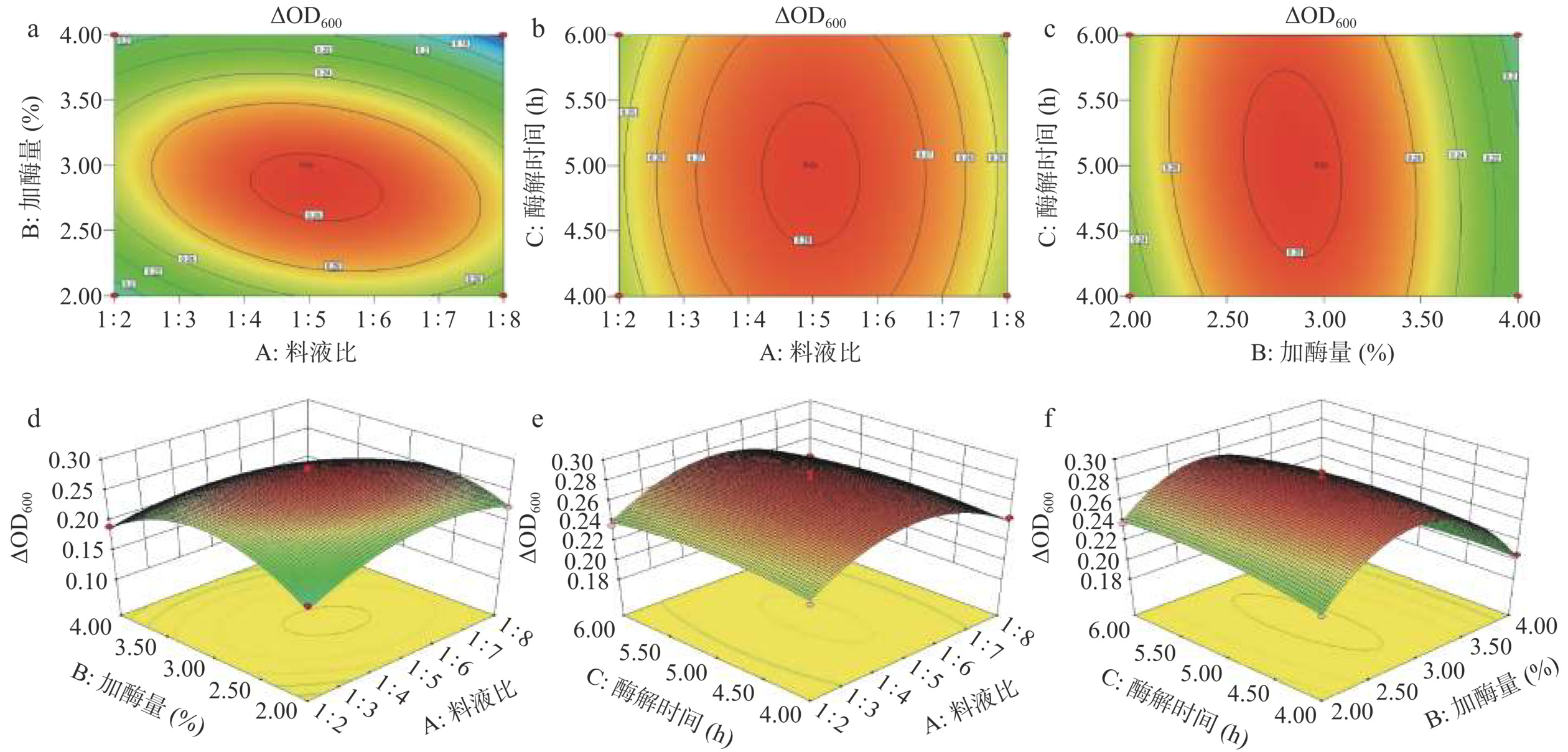
由图5可知,AB、AC交互作用的等高线图线条分布稀疏,趋于圆形,而BC交互作用的等高线图线条分布密集,趋于椭圆形;三维(3D)响应面说明了自变量和因变量之间的关系,响应面坡度表明AB、BC、AC的相互作用依次降低,这与方差分析结果一致。
![]() 图 5 两因素交互作用对ΔOD600影响的响应面图Figure 5. Response surface plot of the interaction between two factors and their impact on ΔOD600
图 5 两因素交互作用对ΔOD600影响的响应面图Figure 5. Response surface plot of the interaction between two factors and their impact on ΔOD6002.4.4 验证试验
通过软件分析获得促进嗜热链球菌FUA329增殖的最佳水解条件为:反应时间5.01 h,料液比1:5.11(w/v),酶浓度2.83%。在此条件下,嗜热链球菌FUA329的理论ΔOD600值为0.283793。结合实际条件,将酶解反应时间修订为5 h,料液比1:5.1(w/v),加酶量2.8%。在修订后的条件下进行3次重复试验验证,ΔOD600值为0.282±0.006,与软件预测值0.283793相近。
2.5 不同分子量HPBL对嗜热链球菌FUA329增殖的影响
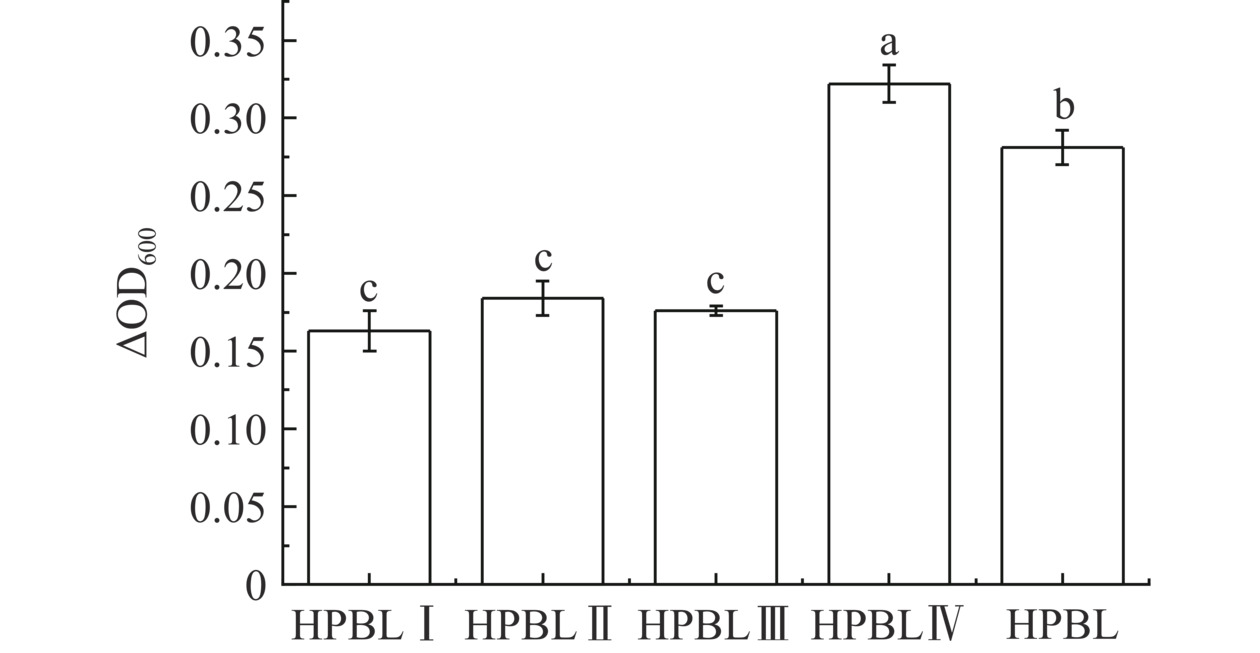
由图6可知,与未分离的酶解产物相比,HPBL Ⅳ对嗜热链球菌FUA329的增殖效果有显著的促进作用(P<0.05)。分子量越小,嗜热链球菌FUA329的增殖效应越明显,证明分子量<3000 Da的肽段是促进嗜热链球菌FUA329增殖的最佳氮源。文献[14-15,31]分别对草鱼皮、凤尾鱼以及大豆进行酶促水解,测定了这三种酶解产物对嗜热链球菌的增殖效果,结果均表明,分子量<3000 Da的肽段促进嗜热链球菌增殖作用效果最好,与本试验的结果一致。初步表明促进嗜热链球菌FUA329增殖的有益肽段的分子量范围<3000 Da,这也验证了Juillard等[30]对于嗜热链球菌更倾向于利用环境中的寡肽进行增殖的结论。
![]() 图 6 不同分子量HPBL对嗜热链球菌FUA329增殖的影响Figure 6. Effects of HPBL with different molecular weight on proliferation of Streptococcus thermophilus FUA329
图 6 不同分子量HPBL对嗜热链球菌FUA329增殖的影响Figure 6. Effects of HPBL with different molecular weight on proliferation of Streptococcus thermophilus FUA3292.6 不同分子量HPBL的抗氧化活性
2.6.1 DPPH自由基清除能力
由图7可知,HPBL I、HPBL Ⅱ、HPBL Ⅲ、HPBL Ⅳ的DPPH自由基清除率分别为48.33%、54.26%、56.73%、68.74%,HPBL对DPPH自由基的清除能力随着分子量的减小而增大,其中HPBL Ⅳ对DPPH自由基的清除能力最强。可能的原因是分子量<3000 Da的HPBL其暴露的氨基酸侧链基团和肽序列更多,能更好地结合DPPH自由基,而分子量较大的HPBL其空间位阻可能较大,从而影响了反应的倾向,使其无法高效地与DPPH自由基结合,对其进行清除[32]。胡凌豪等[15]研究发现分子量<3000 Da的凤尾鱼多肽(AFH1)DPPH清除能力高于未分段的多肽,在溶液中添加的AFH1大于5 mg/mL时,自由基清除率趋于稳定,清除率达到70%,与本文结果相似。
![]() 图 7 不同分子量HPBL对DPPH、羟自由基以及超氧阴离子的清除率Figure 7. Clearance of DPPH, hydroxyl radicals and superoxide anions by different molecular weights HPBL
图 7 不同分子量HPBL对DPPH、羟自由基以及超氧阴离子的清除率Figure 7. Clearance of DPPH, hydroxyl radicals and superoxide anions by different molecular weights HPBL2.6.2 羟自由基清除能力
HPBL I、HPBL Ⅱ、HPBL Ⅲ、HPBL Ⅳ的羟自由基清除率分别为31.35%、34.26%、36.78%和42.23%,HPBL Ⅳ的羟自由基清除率最大。不同分子量多肽对羟基自由基的清除能力不同,分子量越小清除能力越强,这与红花籽多肽的研究结果类似[33]。这是由于分子量较小的多肽可阻断自由基链式反应,促使自由基转化成更加稳定的物质,从而表现出更好的羟基自由基清除能力[34]。
2.6.3 超氧阴离子清除能力
HPBL I、HPBL Ⅱ、HPBL Ⅲ、HPBL Ⅳ的超氧阴离子清除率分别为30.64%、25.16%、27.99%和26.64%,HPBL对超氧阴离子清除能力并未随HPBL分子量的变化显著变化。HPBL清除超氧阴离子自由基的能力可能与酚羟基有关,HPBL I相比于其它三种分子量的肽段含有较多的酚羟基,但总体而言,HPBL含有的酚羟基较少使得其超氧阴离子清除能力较弱。纪莹等[35]的研究结果也发现小黄鱼酶解液对超氧阴离子的清除率均未超过20%且与酶解液的水解度未呈现一定的量效果关系,这可能是由于水解反应的最终产物是氨基酸,而氨基酸抗氧化效果很低,当水解度过高,一些抗氧化能力较强的短肽被水解,不具备抗氧化能力,使得整体抗氧化作用减弱。所以水解程度与抗氧化性的关系并不绝对,控制水解程度,使肽的活性基团暴露出来与自由基发生作用,酶解产物的抗氧化活性才能得到提高。
3. 结论
本文研究了小黄鱼酶解产物对嗜热链球菌FUA329增殖的影响,并对酶解条件进行了优化,发现分子量<3000 Da的肽段促进嗜热链球菌FUA329的增殖效果最优。体外抗氧化实验表明HPBL对于DPPH自由基以及羟自由基的清除效果较好。相较于现有的对于小黄鱼加工副产物的利用方式,本文证明了利用酶解工艺制备促进嗜热链球菌增殖的氮源的可行性,有助于提高水产品加工副产物的经济价值,延长水产品生产的产业链,解决资源浪费与环境污染问题,也对益生菌产业健康可持续发展提供助力。但小黄鱼酶解产物在促进嗜热链球菌增殖的效果上与商用氮源仍有差距,其促进嗜热链球菌增殖的分子机制有待进一步探索。
-
![]()
![]()
图 2 料液比对嗜热链球菌FUA329增殖的影响
Figure 2. Effects of material-liquid ratio on proliferation of Streptococcus thermophilus FUA329
![]()
图 3 加酶量对嗜热链球菌FUA329增殖的影响
Figure 3. Effects of enzyme loading on proliferation of Streptococcus thermophilus FUA329
![]()
图 4 酶解时间对嗜热链球菌FUA329增殖的影响
Figure 4. Effects of hydrolysis time on proliferation of Streptococcus thermophilus FUA329
![]()
图 5 两因素交互作用对ΔOD600影响的响应面图
Figure 5. Response surface plot of the interaction between two factors and their impact on ΔOD600
![]()
图 6 不同分子量HPBL对嗜热链球菌FUA329增殖的影响
Figure 6. Effects of HPBL with different molecular weight on proliferation of Streptococcus thermophilus FUA329
![]()
图 7 不同分子量HPBL对DPPH、羟自由基以及超氧阴离子的清除率
Figure 7. Clearance of DPPH, hydroxyl radicals and superoxide anions by different molecular weights HPBL
表 1 响应面自变量因素与水平
Table 1 Response surface independent variable factors and levels
因素 水平 −1 0 1 A料液比(w/v) 1:2 1:5 1:8 B加酶量(%) 2 3 4 C酶解时间(h) 4 5 6  下载: 导出CSV
下载: 导出CSV
表 2 响应面试验设计与结果
Table 2 Design and results of response surface experimental
实验号 料液比 (A) 加酶量(B) 酶解时间 (C) ΔOD600 (Y) 1 0 1 −1 0.206±0.004 2 0 0 0 0.283±0.004 3 0 0 0 0.278±0.001 4 0 −1 1 0.238±0.005 5 0 −1 −1 0.226±0.002 6 1 0 1 0.242±0.003 7 −1 −1 0 0.193±0.002 8 0 1 1 0.189±0.002 9 1 −1 0 0.224±0.004 10 1 1 0 0.141±0.002 11 −1 1 0 0.191±0.004 12 0 0 0 0.288±0.002 13 −1 0 1 0.236±0.002 14 0 0 0 0.279±0.001 15 0 0 0 0.283±0.007 16 −1 0 −1 0.237±0.006 17 0 0 −1 0.244±0.004
下载: 导出CSV
表 3 回归模型和方差分析
Table 3 Analysis of variance of the regression model
方差来源 平方和 自由度 均方 F值 P值 显著性 模型 0.027 9 2.978×10−3 93.35 < 0.0001 ** A 4.500×10−6 1 4.500×10−6 0.14 0.7183 B 2.965×10−3 1 2.965×10−3 92.93 < 0.0001 ** C 8.000×10−6 1 8.000×10−6 0.25 0.6319 AB 1.640×10−3 1 1.640×10−3 51.42 0.0002 ** AC 2.500×10−7 1 2.500×10−7 7.837×10−3 0.9319 BC 2.102×10−4 1 2.102×10−4 6.59 0.0372 * A2 5.151×10−3 1 5.151×10−3 161.46 < 0.0001 ** B2 0.015 1 0.015 474.77 < 0.0001 ** C2 2.353×10−4 1 2.353×10−4 7.38 0.0300 * 残差 2.233×10−4 7 3.190×10−5 失拟误差 1.605×10−4 3 5.350×10−5 3.41 0.1336 不显著 纯误差 6.280×10−5 4 1.570×10−5 总离差 0.027 16 注:*P<0.05表示显著,**P<0.001表示极显著。
下载: 导出CSV
-
[1] 李国东, 李冬佳, 熊瑛, 等. 基于GAM的黄海南部越冬小黄鱼资源丰度与环境因子关系[J]. 海洋渔业,2023,45(4):403−411. [LI G D, LI D J, XIONG Y, et al. Relationship between environmental factors and abundance of overwintering Larimichthys polyactis in the southern Yellow Sea based on GAM[J]. Marine Fisheries,2023,45(4):403−411.] LI G D, LI D J, XIONG Y, et al. Relationship between environmental factors and abundance of overwintering Larimichthys polyactis in the southern Yellow Sea based on GAM[J]. Marine Fisheries, 2023, 45(4): 403−411.
[2] 沈艳奇, 季广仁, 李学鹏, 等. 低温真空油炸小黄鱼加工工艺优化及包装方式对其贮藏品质的影响[J]. 食品工业科技,2019,40(24):110−119. [SHEN Y Q, JI G R, LI X P, et al. Optimization of low-temperature vacuum fried small yellow croaker processing technology and the effect of packaging method on its quality during storage[J]. Science and Technology of Food Industry,2019,40(24):110−119.] SHEN Y Q, JI G R, LI X P, et al. Optimization of low-temperature vacuum fried small yellow croaker processing technology and the effect of packaging method on its quality during storage[J]. Science and Technology of Food Industry, 2019, 40(24): 110−119.
[3] 张瑜. 水产副产物中功能性成分研究进展[J]. 现代食品,2023,29(11):8−11. [ZHANG Y. Research progress of functional components in aquatic by-products[J]. Modern Food,2023,29(11):8−11.] ZHANG Y. Research progress of functional components in aquatic by-products[J]. Modern Food, 2023, 29(11): 8−11.
[4] 王婧. 利用小黄鱼下脚料开发水产调味料及酶解液生理活性初步研究[D]. 杭州:浙江工商大学, 2011. [WANG J. Preparation of aquatic flavoring and investigation of physiological activity of protein hydrolysate from small yellow croaker scraps[D]. Hangzhou:Zhejiang Gongshang University, 2011.] WANG J. Preparation of aquatic flavoring and investigation of physiological activity of protein hydrolysate from small yellow croaker scraps[D]. Hangzhou: Zhejiang Gongshang University, 2011.
[5] 许韬, 周晓红, 陈玉洁, 等. 小黄鱼内脏油的提取及其棕榈酸甘油酯低温结晶富集工艺的研究[J]. 食品工业科技,2020,41(19):219−225. [XU T, ZHOU X H, CHEN Y J, et al. Extraction of visceral oil from yellow croaker and mass transfer modeling of its palmitic acid enrichment by low temperature crystallization[J]. Science and Technology of Food Industry,2020,41(19):219−225.] XU T, ZHOU X H, CHEN Y J, et al. Extraction of visceral oil from yellow croaker and mass transfer modeling of its palmitic acid enrichment by low temperature crystallization[J]. Science and Technology of Food Industry, 2020, 41(19): 219−225.
[6] 刘冲. 即食小黄鱼制品的开发和研究[D]. 杭州:浙江大学, 2012. [LIU C. Studies on the technology of snack foods with small yellow croaker[D]. Hangzhou:Zhejiang University, 2012.] LIU C. Studies on the technology of snack foods with small yellow croaker[D]. Hangzhou: Zhejiang University, 2012.
[7] ELSOHAIMY S A, HUSSAIN M A. Functional probiotic foods development:trends, concepts, and products[J]. Fermentation,2023,9(3):249−249. doi: 10.3390/fermentation9030249
[8] HARUKA T, KAZUHIKO F, KAZUYA T, et al. Influences of a fermented milk with Lactobacillus bulgaricus and Streptococcus thermophiles on gut associated lymphoid tissue, mucosal IgA, and gut flora in mice[J]. Clinical Nutrition Open Science,2022,42:36−48. doi: 10.1016/j.nutos.2022.01.007
[9] SAFAA A, EBRU G, ILKNUR A C, et al. Lysates from the probiotic bacterium Streptococcus thermophilus enhances the survival of T cells and triggers programmed cell death in neuroblastoma cells[J]. Medical Oncology,2023,40(11):315. doi: 10.1007/s12032-023-02186-4
[10] ITO M, KUSUHARA S, YOKOI W, et al. Streptococcus thermophilus fermented milk reduces serum MDA-LDL and blood pressure in healthy and mildly hypercholesterolaemic adults[J]. Beneficial Microbes,2017,8(2):171−178. doi: 10.3920/BM2016.0102
[11] XUE Z P, CU X, XU K, et al. The effect of glutathione biosynthesis of Streptococcus thermophilus ST-1 on cocultured Lactobacillus delbrueckii ssp. bulgaricus ATCC11842[J]. Journal of Dairy Science,2023,106(2):884−896. doi: 10.3168/jds.2022-22123
[12] 潘晖, 刘欣欣, 孔令慧, 等. 精氨酸代谢调控蛋白ArgR调控嗜热链球菌胞外多糖合成[J]. 微生物学报,2020,60(11):2412−2422. [PAN H, LIU X X, KONG L H, et al. Arginine regulator ArgR regulates exopolysaccharides biosynthesis of Streptococcus thermophilus[J]. Acta Microbiologica Sinica,2020,60(11):2412−2422.] PAN H, LIU X X, KONG L H, et al. Arginine regulator ArgR regulates exopolysaccharides biosynthesis of Streptococcus thermophilus[J]. Acta Microbiologica Sinica, 2020, 60(11): 2412−2422.
[13] PETROVA I, TOLSTOREBROV I, ZHIVLYANTSEVA I, et al. Utilization of fish protein hydrolysates as peptones for microbiological culture medias[J]. Food Bioscience,2021,42(5):101063.
[14] 宋益善, 王肖南, 秦梅, 等. 草鱼皮酶解工艺及水解物对嗜热链球菌增殖作用研究[J]. 科学技术与工程,2016,16(33):249−254. [SONG Y S, WANG X N, QIN M, et al. Study on enzymatic hydrolysis of grass carp Fish skin and promotion of Streptococcus Thermopilus proliferation by the hydrolysates[J]. Science Technology and Engineering,2016,16(33):249−254.] SONG Y S, WANG X N, QIN M, et al. Study on enzymatic hydrolysis of grass carp Fish skin and promotion of Streptococcus Thermopilus proliferation by the hydrolysates[J]. Science Technology and Engineering, 2016, 16(33): 249−254.
[15] 胡凌豪, 葛俊苗, 宋益善, 等. 凤尾鱼酶解工艺优化及其产物促进嗜热链球菌增殖作用[J]. 食品工业科技,2019,40(2):189−194. [HU L H, GE J M, SONG Y S, et al. Optimization on enzymolysis conditions and the proliferation effect on Streptococcus thermophiles growth by the anchovy hydrolysate[J]. Science and Technology of Food Industry,2019,40(2):189−194.] HU L H, GE J M, SONG Y S, et al. Optimization on enzymolysis conditions and the proliferation effect on Streptococcus thermophiles growth by the anchovy hydrolysate[J]. Science and Technology of Food Industry, 2019, 40(2): 189−194.
[16] HORN S, ASPMO S I, EIJSINK V. Growth of Lactobacillus plantarum in media containing hydrolysates of fish viscera[J]. Journal of applied microbilogy,2005,99(5):1082−1089. doi: 10.1111/j.1365-2672.2005.02702.x
[17] IVAR A S, JARLE H S, EIJSINK V G H. Use of hydrolysates from Atlantic cod (Gadus morhua L.) viscera as a complex nitrogen source for lactic acid bacteria[J]. Fems Microbiology Letters,2005,248(1):65−68. doi: 10.1016/j.femsle.2005.05.021
[18] SAFATI R, MOTAMEDZADEGAN A, OVISSIPOUR M, et al. Use of hydrolysates from yellow fin tuna (Thunnus albacares) heads as a complex nitrogen source for lactic acid Bacteria[J]. Food & Bioprocess Technology,2012,5(1):73−79.
[19] FALLAN M, BAHRAM S, JAVADIAN S R. Fish peptone development using enzymatic hydrolysis of silver carp by-products as a nitrogen source in Staphylococcus aureus media[J]. Food Science & Nutrition,2015,3(2):153−157.
[20] SOUISSI N, JRIDI M, SLAMA R B, et al. Sardinelle by-product hydrolyzates as nitrogenI source for microbial growth and protease production[J]. Environmental Engineering and Management Journal,2018,17(11):2677−2684. doi: 10.30638/eemj.2018.266
[21] 芦宇婷. 尿石素B提升肌肉衰减综合征小鼠模型肌肉功能的实验研究[D]. 锦州:锦州医科大学, 2020. [LU Y T. Experimental study of urolithin B in improving muscle function in Sarcopenia mice model[D]. Jinzhou:Jinzhou Medical University, 2020.] LU Y T. Experimental study of urolithin B in improving muscle function in Sarcopenia mice model[D]. Jinzhou: Jinzhou Medical University, 2020.
[22] 田弥, 赵丽蓉, 兰昱, 等. 尿石素A抗癌作用及其机制研究进展[J]. 药物流行病学杂志,2023,32(2):213−220. [TIAN M, ZHAO L R, LAN Y, et al. Anti-cancer effect and its mechanism of urolithin A:Research progress[J]. Chinese Journal of Pharmacoepidemiology,2023,32(2):213−220.] TIAN M, ZHAO L R, LAN Y, et al. Anti-cancer effect and its mechanism of urolithin A: Research progress[J]. Chinese Journal of Pharmacoepidemiology, 2023, 32(2): 213−220.
[23] LIU Q T, LIU S, YE Q W, et al. A novel Streptococcus thermophilus FUA329 isolated from human breast milk capable of producing urolithin A from allagic acid[J]. Foods,2022,11(20):3280. doi: 10.3390/foods11203280
[24] 张孟炜. 转化鞣花单宁产尿石素的菌株筛选及抗衰老功效研究[D]. 无锡:江南大学, 2023. [ZHANG M W. Screening of strains for converting ellagitannin to produce urolithin and study on its anti-aging effect[D]. Wuxi:Jiangnan University, 2023.] ZHANG M W. Screening of strains for converting ellagitannin to produce urolithin and study on its anti-aging effect[D]. Wuxi: Jiangnan University, 2023.
[25] LIU Q T, BIAN Y Y, MU S T, et al. Genomic and phenotypic-based safety assessment and probiotic properties of Streptococcus thermophilus FUA329, a urolithin A-producing bacterium of human milk origin[J]. Genomics,2023,115(6):110724. doi: 10.1016/j.ygeno.2023.110724
[26] LIU Q T, HUA Z Y, CHEN M, et al. Changes in polyphenols and antioxidant properties of pomegranate peels fermented by urolithin A-producing Streptococcus thermophilus FUA329[J]. ACS Food Science & Technology,2023,3(4):1383−1392.
[27] GAO S C, SONG Q, LIU J, et al. E2F1 mediates the downregulation of POLD1 in replicative senescence[J]. Cellular and Molecular Life Sciences:CMLS,2019,76(14):2833−2850. doi: 10.1007/s00018-019-03070-z
[28] 丁慧璞, 欧阳伟虹, 黄玉婷, 等. 小黄鱼边角料的酶解工艺及酶解液性能研究[J]. 核农学报,2020,34(9):2021−2031. [DING H P, OUYANG W H, HUANG Y T, et al. Enzymatic hydrolysis technology of small yellow croaker scrapsand properties of its enzymatic hydrolysis solution[J]. Journal of Nuclear Agricultural Sciences,2020,34(9):2021−2031.] DING H P, OUYANG W H, HUANG Y T, et al. Enzymatic hydrolysis technology of small yellow croaker scrapsand properties of its enzymatic hydrolysis solution[J]. Journal of Nuclear Agricultural Sciences, 2020, 34(9): 2021−2031.
[29] 陈建康, 包建强. 酶解法制备草鱼皮蛋白水解物增殖嗜热链球菌[J]. 食品科学,2016,37(5):144−148. [CHEN J K, BAO J Q. Enzymatic preparation of protein hydrolysate from grass carp skin for use as a mitrogen source for Streptococcus thermophilus growth[J]. Food Science,2016,37(5):144−148.] CHEN J K, BAO J Q. Enzymatic preparation of protein hydrolysate from grass carp skin for use as a mitrogen source for Streptococcus thermophilus growth[J]. Food Science, 2016, 37(5): 144−148.
[30] JUILLARD N V, LAAN H, KUNJI E, et al. The extracellular P-I-Type proteinase of Lactococcus lactis hydrolyzes Beta-Casein into more than 100 different oligopetides[J]. Journal of Bacteriology,1995,177(12):3472−3478. doi: 10.1128/jb.177.12.3472-3478.1995
[31] 支潇, 田晓宇, 田辉, 等. 水解大豆蛋白对嗜热链球菌增殖的促进作用[J]. 食品科技,2012,37(8):13−17. [ZHI X, TIAN X Y, TIAN H, et al. Effect of soybean protein hydrolysates on promotion of Streptococcus thermophilus[J]. Food Science and Technology,2012,37(8):13−17.] ZHI X, TIAN X Y, TIAN H, et al. Effect of soybean protein hydrolysates on promotion of Streptococcus thermophilus[J]. Food Science and Technology, 2012, 37(8): 13−17.
[32] 石晓岩. 蓝莓中花青素的抗氧化活性研究[J]. 中国食品添加剂,2023,34(12):162−168. [SHI X Y. Study on antioxidant activity of anthocyanins from blueberries[J]. China Food Additives,2023,34(12):162−168.] SHI X Y. Study on antioxidant activity of anthocyanins from blueberries[J]. China Food Additives, 2023, 34(12): 162−168.
[33] 刘晓艺, 周玉岩, 过利敏, 等. 不同分子量红花籽抗氧化肽稳定性研究[J]. 食品工业科技,2022,43(13):94−102. [LIU X Y, ZHOU Y Y, GUO L M, et al. Study on the stability of antioxidant peptides from safflower seeds with different molecular weight[J]. Science and Technology of Food Industry,2022,43(13):94−102.] LIU X Y, ZHOU Y Y, GUO L M, et al. Study on the stability of antioxidant peptides from safflower seeds with different molecular weight[J]. Science and Technology of Food Industry, 2022, 43(13): 94−102.
[34] 张雯雯, 张艳妮, 杨丽荣, 等. 4种天然物质抗氧化能力的研究[J]. 食品研究与开发,2022,43(2):43−50. [ZHANG W W, ZHANG Y N, YANG L R, et al. Study of the antioxidant capacities of four natural substances[J]. Food Research and Development,2022,43(2):43−50.] ZHANG W W, ZHANG Y N, YANG L R, et al. Study of the antioxidant capacities of four natural substances[J]. Food Research and Development, 2022, 43(2): 43−50.
[35] 纪莹, 张国增. 小黄鱼蛋白酶解液抗氧化活性研究[J]. 粮油加工(电子版),2014(1):72−74,79. [JI Y, ZHANG G Z. Antioxidative activity of enzymatic hydrolysate from little yellow croaker protein[J]. Cereals and Oils Processing(Electronic Version),2014(1):72−74,79.] JI Y, ZHANG G Z. Antioxidative activity of enzymatic hydrolysate from little yellow croaker protein[J]. Cereals and Oils Processing(Electronic Version), 2014(1): 72−74,79.
 下载:
下载:










计量
- 文章访问数: 82
- HTML全文浏览量: 11
- PDF下载量: 14



