


Bioutilization of Arca subcrenata Myofibrillar Protein by Four Probiotics
-
摘要: 以毛蚶为研究对象,考察植物乳植杆菌(Lactiplantibacillus plantarun,LP45)、两歧双歧杆菌BBi32(Bifidobacterium bifidobacterial,BBi32)、鼠李糖乳酪杆菌LGG(Lacticaseibacillus rhamnosus,LGG)以及动物双歧杆菌乳亚种Probio-M8(Bifidobacterium animalis subsp. lactis Probio-M8,Probio-M8)四种益生菌对毛蚶肌原纤维蛋白模拟消化产物生物利用的差异性。将毛蚶肌原纤维蛋白进行体外模拟消化,以消化产物作为氮源,分别采用氮源替换和氮源添加两种方式培养四种益生菌,通过监测四种益生菌的生长曲线、pH、菌落数等变化,评价不同益生菌对毛蚶肌原纤维蛋白模拟消化产物的利用特性。结果表明,以MRS组作为对照组,氮源替换组可显著抑制四种益生菌的增殖(P<0.05),但对动物双歧杆菌乳亚种Probio-M8的抑制作用较低;当毛蚶肌原纤维蛋白模拟消化产物作为氮源添加至培养基中,该蛋白模拟消化产物可显著促进动物双歧杆菌乳亚种Probio-M8的增殖;当添加氮源浓度为0.03 mg/mL时,动物双歧杆菌乳亚种Probio-M8活菌数为对照组的1.2倍(P<0.05),但对胞外聚合物的产量无显著影响。研究显示,氮源替换时,毛蚶肌原纤维蛋白模拟消化产物可抑制益生菌的增殖;氮源添加时,毛蚶肌原纤维蛋白模拟消化产物在低浓度时对四种益生菌的增殖均有一定的促进作用,但对不同益生菌的促进程度不同,表明不同种类益生菌对氮源的需求不同,该研究为益生菌的氮源生物利用提供了理论指导。Abstract: Using Arca subcrenata as the research object, this study explored the differences in the utilization of simulated digestion products of Arca subcrenata myofibrillar protein by probiotics, namely Lactiplantibacillus plantarun, Bifidobacterium bifidobacterial, Lacticaseibacillus rhamnosus and Bifidobacterium animalis subsp. Lactis Probio-M8. The study first simulated the in vitro digestion of myofibrillar protein from Arca subcrenata and used it as a nitrogen source to co-culture with probiotics by nitrogen source replacement and nitrogen source addition, respectively. Subsequently, the utilization characteristics by the probiotics on the digestion products of myofibrillar protein from Arca subcrenata were evaluated by monitoring the growth curve, pH and colony number among others. The results showed that, compared to the de Man-Rogosa-Sharpe group, the nitrogen source replacement group could significantly inhibit the proliferation of the four probiotic species (P<0.05), but the inhibitory effect on Probio-M8 was weak. When the simulated digestion products of Arca subcrenata myofibrillar protein were added to the medium as the nitrogen source, the protein products could significantly promote the proliferation of Probio-M8. At the protein concentration of 0.03 mg/mL, the number of Probio-M8 viable bacteria was 1.2 times higher than that of the control group, implying the protein significantly promoted the growth of Probio-M8 (P<0.05). However, Arca subcrenata myofibrillar protein products had no significant effect on extracellular polymer substances. The results showed that, when the nitrogen source was replaced, simulated digestion products of Arca subcrenata myofibrillar protein inhibited the proliferation of probiotics. When the nitrogen source was added, the simulated digestion products of Arca subcrenata myofibrillar protein at low concentrations had probiotic-specific growth-promoting effect on the four probiotics. It indicated that different probiotics have different nitrogen source requirements. These results would provide theoretical guidance for the bio-utilization of nitrogen sources by probiotics.
-
双歧杆菌和乳杆菌是人体肠道中的主要益生菌,对宿主健康具有很大的影响[1],包括免疫系统的调节、与肠道微生物的互作、肠黏膜屏障功能的改善以及参与营养物质的分解和合成[2]等多方面功能,其中外源摄入营养物质可作为益生菌生长底物[3]。蛋白质经口摄入,在胃肠消化后转化为肽和氨基酸等消化产物,此类消化产物可作为微生物群的氮源和能源[4−6]。大多数乳酸菌是氨基酸营养缺陷型菌株[7],不能利用外源蛋白质供自身的营养需求,必须通过蛋白水解系统将蛋白水解成小肽和氨基酸[8]。该蛋白水解系统在不同的乳酸菌株中具有很强的菌株特异性,乳酸菌在适应不同的生长环境时会通过自身基因如点突变、重组或者重复序列等变异方式来获得新基因[9],进而导致益生菌对同一氮源的利用效率存在差异,这意味着未被利用的氮源成分将通过增加培养基的渗透压,对菌株的生长产生抑制作用。因此,氮源是否被利用是制约其生长的一个关键因素。
水产品作为一种广泛易得的大宗原料,可为微生物提供丰富的氮源。为增加鱼类废物的商业价值,研究表明,鱼蛋白水解物可作为细菌培养基的营养替代物[10];鱼粉可促进大鼠体内丁酸杆菌和乳酸杆菌增殖[11];鳓鱼蛋白可促进植物乳植杆菌的黏附[12]。蚶是一类高蛋白低脂肪的水产品[13],其粗蛋白含量高达14.8%[14]。基于本课题组研究发现,毛蚶蛋白主要由肌原纤维蛋白组成[15],毛蚶蛋白和多肽具有丰富的生物活性。毛蚶蛋白水解物ASAH通过调控小鼠巨噬细胞中MAPK依赖性HO-1/Nrf2通路显示出潜在的抗炎活性[16];从毛蚶中鉴定的多肽MCLDSCLL和HPLDSLCL,显示出较强的抗氧化性,IC50值为19.47±0.35和74.36±0.54 μmol/L[17];毛蚶蛋白ASP-3具有抗肿瘤活性,对HepG2细胞具有很强的增殖抑制作用,IC50值为171.18±18.59 μg/mL[18]。目前已有学者对毛蚶蛋白的部分功能活性进行了研究,但其对益生菌的作用尚未被明确,不同益生菌对其氮源的利用方式存在差异,而氮源的利用一直是制约乳酸菌生长的关键因素。因此,本研究深入探究四种益生菌对毛蚶肌原纤维蛋白的利用特征,可为益生菌的生产提供氮源的理论优选策略。
不同乳酸菌氮代谢系统的底物特异性决定了其可能对同一氮源具有不同的利用效率。本文以毛蚶肌原纤维蛋白为研究对象,探究该蛋白模拟消化产物对四种肠道益生菌(植物乳植杆菌LP45、两歧双歧杆菌BBi32、鼠李糖乳酪杆菌LGG以及动物双歧杆菌乳亚种Probio-M8)的生长特性的影响,该研究可为益生菌的氮源利用提供理论指导。
1. 材料与方法
1.1 材料与仪器
毛蚶(新鲜活体) 广州市黄沙水产市场;植物乳植杆菌LP45(LP45,编号YMC1005)、动物双歧杆菌乳亚种Probio-M8(Probio-M8)、鼠李糖乳酪杆菌LGG(LGG)、两歧双歧杆菌(BBi32) 中国工业微生物菌种保藏中心:胃蛋白酶(3000 U/mg)、胰蛋白酶(32.6 U/mg) 浙江中牧药业有限公司;考马斯亮蓝G250、牛血清白蛋白 上海伯奥生物科技有限公司;酵母浸膏、葡萄糖、吐温-80、蛋白胨、牛肉浸膏 生工生物工程(上海)股份有限公司;其余试剂均为国产分析纯。
SCIENTZ-18N冷冻干燥机 宁波新芝生物科技股份有限公司;HYQX-ΙΙΙ厌氧培养箱 上海新诺仪器集团有限公司;Synergy H1M多功能酶标仪 美国伯腾仪器有限公司;台式G6 Phenom Pro扫描电镜 复纳科学仪器(上海)有限公司;UV-1800紫外分光光度计 日本岛津公司;HYQX-ΙΙΙ厌氧培养箱 上海新诺仪器集团有限公司。
1.2 实验方法
1.2.1 毛蚶肌原纤维蛋白的提取
毛蚶去壳、去内脏,按照1:4(W/V)的比例加入4 ℃超纯水,均质机均质90 s,制成样品匀浆,4 ℃静置90 min,4000 r/min离心10 min,保留上清液除去沉淀,沉淀重复上述步骤,将两次上清液合并即为毛蚶肌原纤维蛋白,冷冻干燥后保存备用。
1.2.2 毛蚶肌原纤维蛋白的SDS-PAGE电泳分析
制样:取适量样品配制成浓度为2 mg/mL的溶液,蛋白样品与5×蛋白上样品缓冲液以4:1的体积混合,沸水浴5 min,上样量为10 μL。制胶:配制12%分离胶、5%浓缩胶。电泳:恒定电压85 V。染色与脱色:用0.125%考马斯亮蓝R-250染色4 h,脱色液(冰醋酸10%,甲酸30%)脱色至条带清晰。使用凝胶成像系统收集凝胶图像并对电泳条带进行分析。
1.2.3 毛蚶肌原纤维蛋白体外模拟消化
毛蚶肌原纤维蛋白质体外模拟消化参考André等[19]的方法。消化液电解质储备液(1.25×)的配制见表1。
表 1 消化液电解质储备液的配制Table 1. Preparation of electrolyte stock solutions of digestion fluids盐溶液 储备液浓度
(mol/L)SSF(pH7)
1.25×(mmol/L)SGF(pH3)
1.25×(mmol/L)SIF(pH7)
1.25×(mmol/L)KCl 0.50 15.10 6.90 6.80 KH2PO4 0.50 3.70 0.90 0.80 NaHCO3 1.00 13.60 25.00 85.00 NaCl 2.00 − 47.20 38.40 MgCl·6H2O 0.15 0.15 0.12 0.33 (NH4)2CO3 0.50 0.06 0.50 − HCl 6.00 0.09 15.60 8.40 CaCl2·2H2O 0.30 1.50 0.15 0.60 模拟口服期消化:口腔消化储备液(Simulated Salivary Fluid,SSF)预热到37 ℃,记录食物体积为V0,加入SSF,0.3 mol/L CaCl2至1.5 mmol/L,补加水至SSF浓度为1×,最终体积为2V0,37 ℃恒温振荡培养箱,140 r/min,消化2 min。
模拟胃期消化:模拟胃消化储备液(Simulated Gastric Fluid,SGF)预热到37 ℃,加入0.3 mol/L CaCl2至0.15 mmol/L,调节pH为3.0,加入胃蛋白酶至2000 U/mL,补加水至SGF浓度为1×,最终体积为4V0,37 ℃恒温振荡培养箱,140 r/min,消化120 min;将pH调至8.0,终止胃消化。
模拟肠期消化:模拟肠消化储备液(Simulated Intestinal Fluid,SIF)预热到37℃,加入0.3 mol/L CaCl2至0.6 mmol/L,胆汁至10 mmol/L,调节pH至7.0,加入胰蛋白酶至100 U/mL,补加水至SIF浓度为1×,最终体积为8V0,37 ℃恒温振荡培养箱,140 r/min,消化120 min;100 ℃沸水浴5 min,停止肠消化,放置室温,4000 r/min,离心10 min,收集上清液,于4 ℃冷藏备用。
1.2.4 毛蚶肌原纤维蛋白模拟消化产物蛋白含量的测定
毛蚶肌原纤维蛋白模拟消化产物中的蛋白含量测定参照考马斯亮蓝法。得蛋白质标准曲线方程y=0.001483x+0.6931,R2=0.9908,毛蚶蛋白模拟消化液蛋白含量为4.43 mg/mL。
1.2.5 毛蚶肌原纤维蛋白模拟消化产物对益生菌生长的影响
1.2.5.1 培养基的配制
MRS(de Man-Rogosa-Sharpe,MRS)培养基(10×)的配制:蛋白胨10 g、牛肉浸膏5 g、葡萄糖20 g、吐温-80 1 g、磷酸二氢钠2 g、无水乙酸钠5 g、酵母浸膏5 g、柠檬酸三铵2 g、硫酸锰0.02 g、硫酸镁0.1 g、蒸馏水100 mL,121℃,灭菌30 min。
氮源缺乏MRS(10×)培养基的配制:牛肉浸膏5 g、葡萄糖20 g、吐温-80 1 g、磷酸二氢钠2 g、无水乙酸钠5 g、酵母浸膏5 g、柠檬酸三铵2 g、硫酸锰0.02 g、硫酸镁0.1 g、蒸馏水100 mL,121 ℃,灭菌30 min。
MRS(1×)培养基的配制:MRS培养基(10×)与无菌水以1:9(v/v)混匀。
氮源缺乏MRS(1×)培养基(BMRS)的配制:氮源缺乏MRS(10×)培养基与无菌水以1:9(v/v)混匀。
氮源替换MRS(1×)培养基(TMRS)的配制:向氮源缺乏MRS(10×)培养基添加与蛋白胨等氮含量的毛蚶肌原纤维蛋白模拟消化液,配制成与MRS(1×)培养基等氮含量的培养基。
氮源添加MRS培养基的配制:MRS培养基(10×)、无菌水和毛蚶肌原纤维蛋白模拟消化液以适当比例混合,配制含有毛蚶肌原纤维蛋白模拟消化液蛋白浓度为0.01、0.03、0.05、0.1、0.5 mg/mL的MRS(1×)培养基。
以MRS(1×)为对照组,氮源缺乏MRS(1×)培养基(BMRS)、氮源替换MRS(1×)培养基(TMRS)和含有不同浓度氮源添加的MRS培养基为实验组。
1.2.5.2 益生菌培养条件
在超净工作台中,取菌液以2%(v/v)接种量接种于MRS培养基中,37 ℃,200 r/min,培养12 h制得新鲜菌液。
将新鲜菌液以2%比例接种于含有毛蚶肌原纤维蛋白模拟消化液蛋白浓度为0.01、0.03、0.05、0.1、0.5 mg/mL的MRS培养基中,混匀后以200 μL/孔接种于96孔板中,每组设置3个平行,用封口膜密封,置于多功能酶标仪中。在37 ℃、OD600 nm下检测24 h益生菌的生长情况。绘制生长曲线,计算代时和曲线下面积(Area under Curve,AUC)。
G=ln2k 式中:k表示lnOD600与时间T拟合直线的斜率;G表示代时,h。
1.2.6 毛蚶肌原纤维蛋白模拟消化产物对益生菌菌液pH的影响
测定动物双歧杆菌乳亚种Probio-M8在毛蚶肌原纤维蛋白添加浓度为0.03 mg/mL时培养基的pH的变化,以MRS培养基为对照。恒温振荡培养箱37 ℃、200 r/min恒温振荡培养箱培养24 h。每4 h在超净工作台中取2 mL菌液,用pH计测定对应时间菌液的pH。
1.2.7 毛蚶肌原纤维蛋白模拟消化产物对益生菌活菌数的影响
将动物双歧杆菌乳亚种Probio-M8菌菌液以2%(v/v)接种量接种于毛蚶肌原纤维蛋白添加浓度为0.03 mg/mL的MRS培养基,以MRS培养基为对照。37 ℃、200 r/min恒温振荡培养箱培养24 h,稀释106倍后,取100 μL菌液接种于MRS固体培养基,用涂布棒涂布均匀,37℃厌氧培养箱培养24 h,使用Image J软件计算菌落数。
N=10C10−D 式中:N表示菌落数,CFU/mL;C表示培养基菌落数;D表示稀释倍数。
1.2.8 毛蚶肌原纤维蛋白模拟消化产物对益生菌胞外聚合物的影响
动物双歧杆菌乳亚种Probio-M8菌菌液以2%(v/v)接种量接种于毛蚶肌原纤维蛋白添加浓度为0.03 mg/mL的MRS培养基中,以MRS培养基为对照,37 ℃、200 r/min培养24 h,菌悬液7000 r/min离心5 min,用PBS洗涤3次(pH=7.4),放置于烘箱,35℃过夜。成像前,对样品喷金180 s以增加导电性,扫描电子显微镜下拍摄图像:加速电压5 kV,探测器模式为Mix 50%,光束强度为Image。
1.3 数据处理
每组实验独立重复3次。实验数据采用Image J和prism 8.0中单因素方差分析(one-way ANOVA)进行统计学处理,P<0.05为统计学意义上有显著差异。绘图采用prism 8.0软件进行。
2. 结果与分析
2.1 毛蚶肌原纤维蛋白的蛋白质组成分析
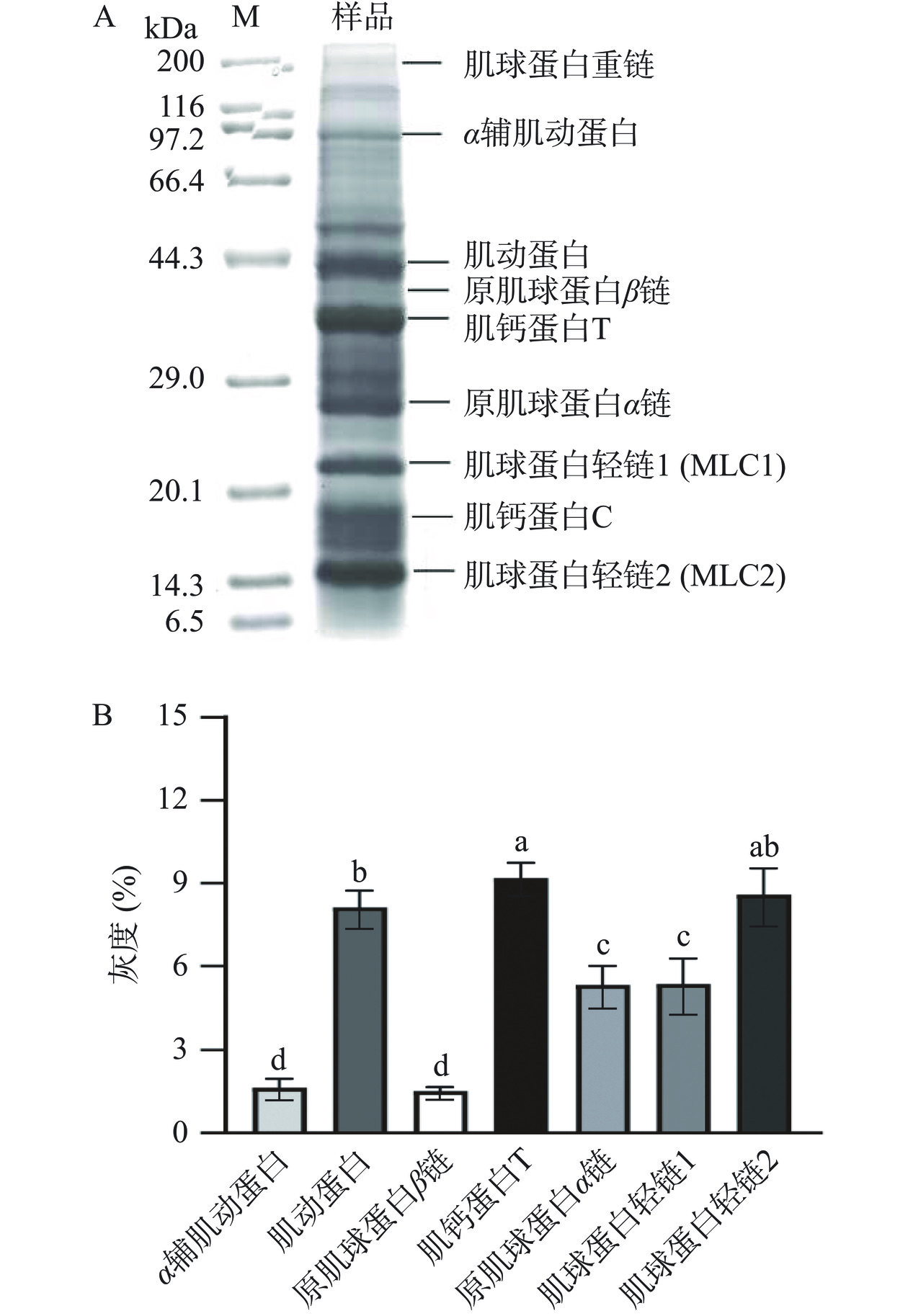
对毛蚶肌原纤维蛋白进行SDS-PAGE电泳分析,并对肌原纤维蛋白的电泳谱图进行定量分析。如图1A所示,毛蚶肌原纤维蛋白包括粗丝蛋白和细丝蛋白,粗丝蛋白包括形成核心的副肌球蛋白;细丝蛋白包括肌动蛋白(约42 kDa)和肌钙蛋白T(约35 kDa)等,此外,毛蚶肌原纤维蛋白还包括几个低强度条带,如原肌球蛋白α链(约27 kDa)、原肌球蛋白β链(37 kDa)和α辅肌动蛋白(97 kDa)等,与Jin等[20]结果相似。如图1B所示,毛蚶肌原纤维蛋白中肌钙蛋白T和肌球蛋白轻链2占比较大,约为α辅肌动蛋白和原肌球蛋白β链的3倍。上述结果表明,肌钙蛋白T和肌球蛋白轻链在毛蚶肌原纤维蛋白含量较多。
![]() 图 1 毛蚶蛋白的蛋白质组成分析注:A. 毛蚶蛋白的电泳图谱;B. 电泳条带灰度分析;不同小写字母代表两组差异显著(P<0.05)。Figure 1. Analysis the protein composition of Arca subcrenata protein
图 1 毛蚶蛋白的蛋白质组成分析注:A. 毛蚶蛋白的电泳图谱;B. 电泳条带灰度分析;不同小写字母代表两组差异显著(P<0.05)。Figure 1. Analysis the protein composition of Arca subcrenata protein2.2 毛蚶肌原纤维蛋白模拟消化产物对益生菌生长特性的影响
2.2.1 毛蚶肌原纤维蛋白模拟消化产物对益生菌生长状态的影响
生长曲线是描述微生物生长状况的基本指标,反映微生物生长与时间的对应关系。图2为探究毛蚶肌原纤维蛋白模拟消化产物等氮替换MRS培养基中的蛋白胨对四种益生菌生长状态的影响。OD值可反应益生菌增殖情况,OD值越高,细菌的增殖效果越明显。从生长曲线可知,MRS组与BMRS组相比,BMRS组并未对四种益生菌的增殖产生显著影响(P>0.05),可能BMRS培养基已为益生菌提供充足的氮源;当用毛蚶肌原纤维蛋白模拟消化产物等氮替换时,其OD值和AUC均小于MRS组和BMRS组,除动物双歧杆菌乳亚种Probio-M8外,其余三种益生菌TMRS组AUC为对照组的0.5倍,四种菌均指数期时间缩短,提前到达稳定期,可能BMRS组的培养基固形物较多,导致培养基渗透压增大,抑制微生物的生长[21]。
![]() 图 2 四种益生菌在毛蚶肌原纤维蛋白模拟消化产物存在条件下的生长曲线及其积分面积注:不同小写字母代表两组差异显著(P<0.05)。Figure 2. Growth curves and area growth curves of four probiotics in the presence of simulated digestion products of Arca subcrenata myofibrillar protein
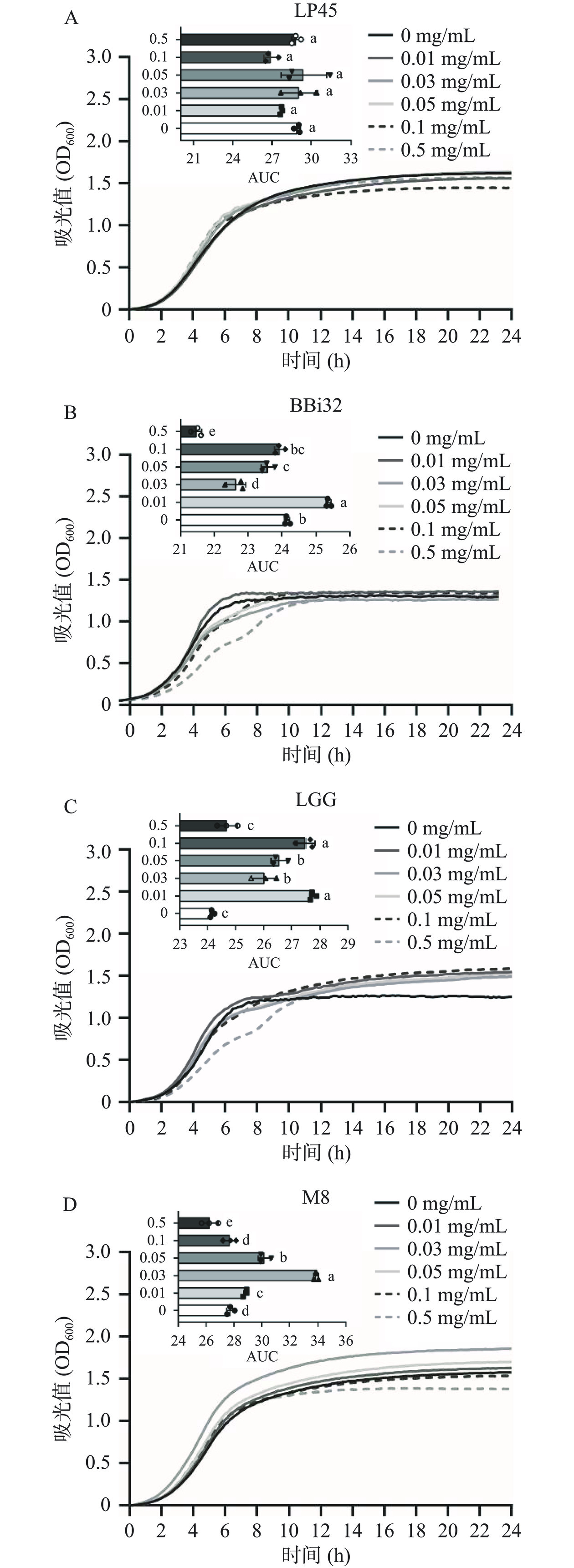
图 2 四种益生菌在毛蚶肌原纤维蛋白模拟消化产物存在条件下的生长曲线及其积分面积注:不同小写字母代表两组差异显著(P<0.05)。Figure 2. Growth curves and area growth curves of four probiotics in the presence of simulated digestion products of Arca subcrenata myofibrillar protein图3展示了不同浓度毛蚶肌原纤维蛋白模拟消化产物对四种益生菌生长状态的影响。从生长曲线可知,指数期后,益生菌生长迅速,导致菌液浊度升高,OD600增长速度最快;随着代谢产物的积累,生长曲线趋于平稳,即益生菌生长状态进入稳定期。当肌原纤维蛋白模拟消化产物浓度较低时,四种益生菌在延滞期和指数期的生长曲线形态与其对照组相似,稳定期略有不同:植物乳植杆菌LP45稳定期的OD600低于对照组,表明毛蚶肌原纤维蛋白模拟消化产物对植物乳植杆菌LP45的增殖作用不显著,其余三种菌稳定期的OD600略高于对照组,且当蛋白浓度为0.03 mg/mL时,动物双歧杆菌乳亚种Probio-M8菌的OD600显著(P<0.05)高于对照组。研究表明不同益生菌对同种蛋白质的利用能力不同,当MRS培养基或较低浓度的毛蚶肌原纤维蛋白模拟消化产物为益生菌提供充足的氮源时,此类微生物不需要额外的蛋白质[22]。AUC可全面评估四种益生菌生长过程中的变化[23],从图3A~图3D可知,与对照组相比,蛋白浓度为0.01、0.03、0.05、0.1 mg/mL均对益生菌的增殖有不同程度的促进作用。图3A显示,植物乳植杆菌LP45的AUC与蛋白浓度无关;图3B和图3C显示,两歧双歧杆菌BBi32和鼠李糖乳酪杆菌LGG在毛蚶蛋白浓度为0.01 mg/mL的培养基下生长最佳;图3D显示,当蛋白浓度为0.03 mg/mL时,AUC为对照组的1.2倍,该浓度蛋白可显著促进动物双歧杆菌乳亚种Probio-M8的生长(P<0.05)。除植物乳植杆菌LP45外,当蛋白浓度超过0.5 mg/mL时,AUC较对照组显著降低(P<0.05),可能由于蛋白含量较高,导致培养基渗透压过高[21],细菌产生脱水反应,抑制了菌体的增殖。
![]() 图 3 四种益生菌在不同浓度毛蚶肌原纤维蛋白模拟消化产物存在条件下的生长曲线及其积分面积注:不同小写字母代表两组差异显著(P<0.05)。Figure 3. Growth curves and area growth curves of four probiotics in the presence of different concentrations of simulated digestion products of Arca subcrenata myofibrillar protein
图 3 四种益生菌在不同浓度毛蚶肌原纤维蛋白模拟消化产物存在条件下的生长曲线及其积分面积注:不同小写字母代表两组差异显著(P<0.05)。Figure 3. Growth curves and area growth curves of four probiotics in the presence of different concentrations of simulated digestion products of Arca subcrenata myofibrillar protein2.2.2 毛蚶肌原纤维蛋白模拟消化产物对益生菌代时的影响
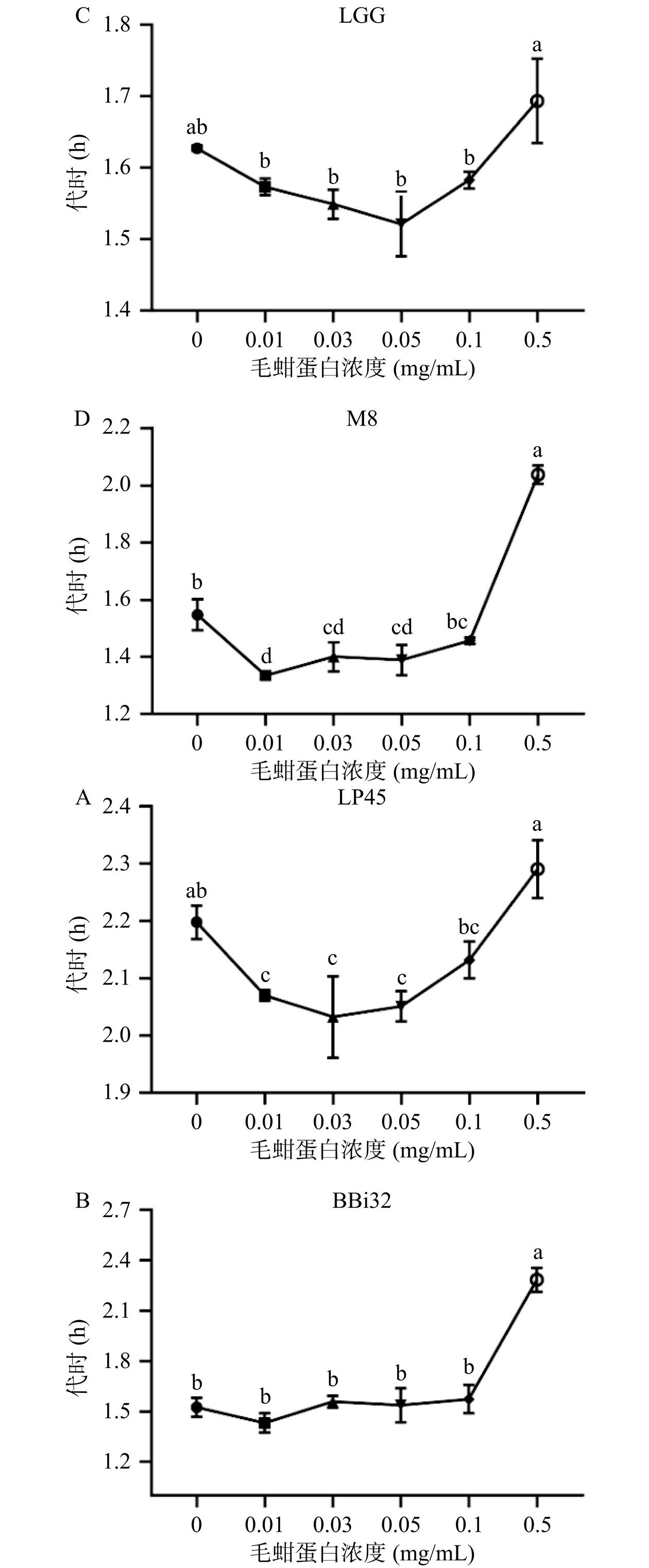
每个微生物分裂繁殖一代所需的时间称为代时,因此,代时是评价微生物生长特性的重要指标之一。图4显示,毛蚶肌原纤维蛋白浓度对益生菌代时的影响并不呈浓度依赖性,低浓度蛋白(<0.05 mg/mL)可缩短代时,而高浓度蛋白(>0.05 mg/mL)则会延长代时。这表明低浓度蛋白的添加即可满足益生菌生长氮源需求,而高浓度蛋白的加入反而不利于益生菌生长。低浓度的毛蚶肌原纤维蛋白可缩短植物乳植杆菌LP45、鼠李糖乳酪杆菌LGG和动物双歧杆菌乳亚种Probio-M8的代时,高浓度的蛋白均可显著延长四种益生菌的代时(P<0.05)。研究人员[24]发现竹虫蛋白酶解液可缩短益生菌的代时,促进益生菌的复制;相比之下,低浓度蛋白对动物双歧杆菌乳亚种Probio-M8的促生长效果较为显著(P<0.05),蛋白浓度为0.03 mg/mL和0.01 mg/mL时,动物双歧杆菌乳亚种Probio-M8代时分别缩短为对照组的0.90倍和0.86倍,且结合图3D可知,蛋白浓度为0.03 mg/mL和0.01 mg/mL时,动物双歧杆菌乳亚种Probio-M8代时分别增加AUC为对照组的1.21倍1.04倍。因此,选择动物双歧杆菌乳亚种Probio-M8菌以及0.03 mg/mL的蛋白浓度进行后续实验。
![]() 图 4 四种益生菌在不同浓度毛蚶肌原纤维蛋白模拟消化产物存在条件下的代时注:不同小写字母代表两组差异显著(P<0.05)。Figure 4. Generation time of four probiotics in the presence of different concentrations of simulated digestion products of Arca subcrenata myofibrillar protein
图 4 四种益生菌在不同浓度毛蚶肌原纤维蛋白模拟消化产物存在条件下的代时注:不同小写字母代表两组差异显著(P<0.05)。Figure 4. Generation time of four probiotics in the presence of different concentrations of simulated digestion products of Arca subcrenata myofibrillar protein2.3 毛蚶肌原纤维蛋白模拟消化产物对益生菌菌液pH的影响
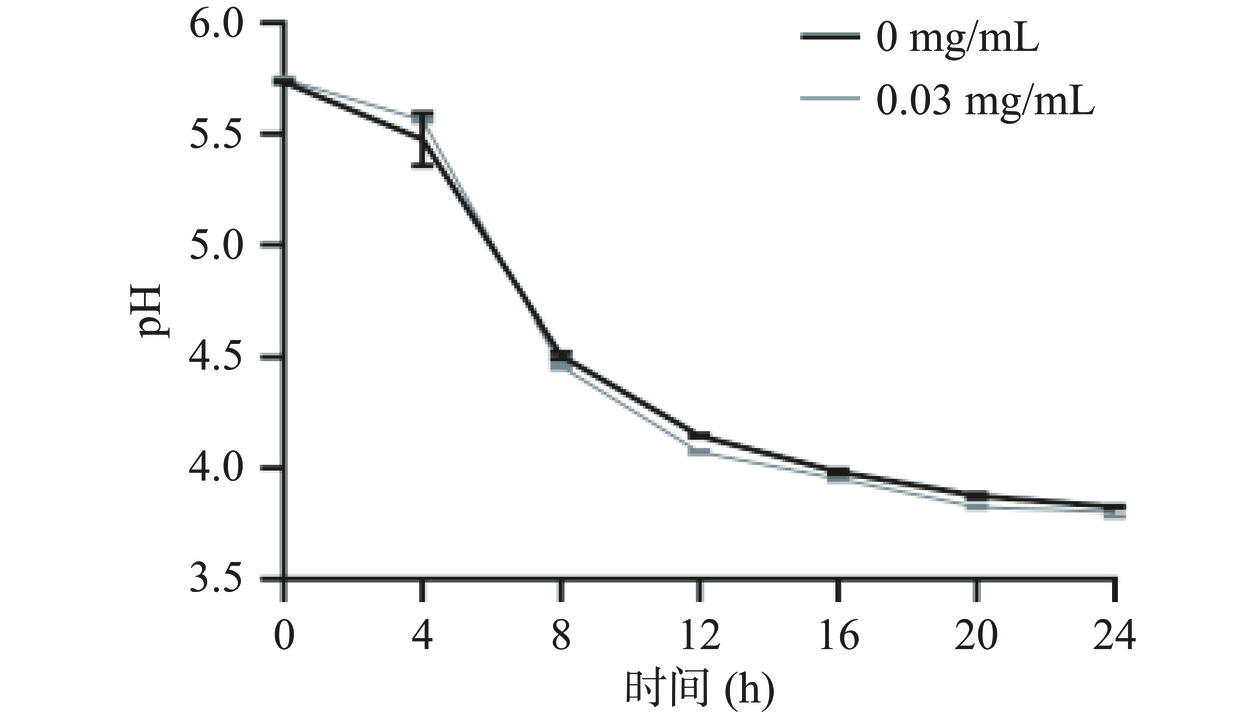
益生菌在培养基中进行生长和繁殖时,产生乳酸等有机酸,从而导致菌液的pH下降。因此,菌液pH变化是益生菌生长特性的重要评价指标之一,可通过培养基pH变化反映毛蚶蛋白对动物双歧杆菌乳亚种Probio-M8生长的影响[25]。由图5可知,益生菌培养体系pH在0~4 h基本无变化,pH在4~8 h明显降低,在8 h后pH变化趋于平缓。4~8 h益生菌生长处于活跃期,该阶段益生菌产酸量最多;稳定期由于培养基pH过低,动物双歧杆菌乳亚种Probio-M8增殖受到抑制,生长曲线变化趋于平缓,表明pH过低不利于毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8的益生作用;总体而言,该蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8菌液pH无显著性影响,结果与Liu等[12]基本一致。
![]() 图 5 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8菌液pH的影响Figure 5. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata myofibrillar protein on the pH of bacterial solution of Probio-M8
图 5 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8菌液pH的影响Figure 5. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata myofibrillar protein on the pH of bacterial solution of Probio-M82.4 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8活菌数的影响
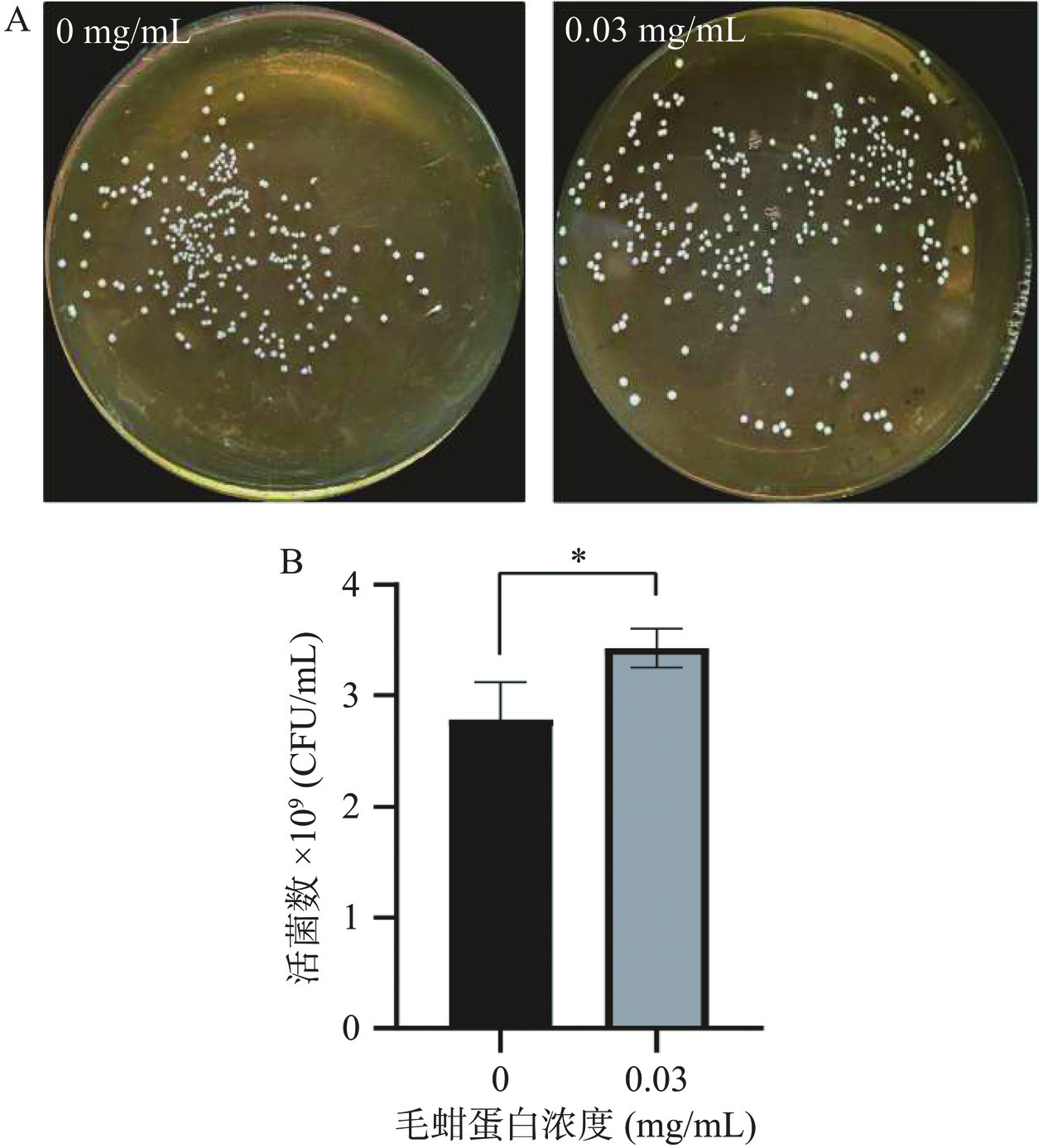
AUC可反映细菌总数,但无法反映培养体系中的活菌数。为进一步确定0.03 mg/mL毛蚶肌原纤维蛋白对动物双歧杆菌乳亚种Probio-M8生长特性的影响,采用平板涂布法测定0.03 mg/mL毛蚶蛋白培养体系中Probio-M8的活菌数。图6A可观察到在MRS固体培养基中,动物双歧杆菌乳亚种Probio-M8菌落表面光滑,呈乳白色,形态呈凸圆不透明状,与王元驰等[26]研究中动物双歧杆菌乳亚种Probio-M8形态相似。此外,与对照组相比(菌落数2.82×109 CFU/mL),加入毛蚶蛋白可促进益生菌的增长(菌落数3.43×109 CFU/mL)(P<0.05),菌落数为对照组的1.2倍,由图6B可知。结果表明,毛蚶肌原纤维蛋白浓度为0.03 mg/mL可促进动物双歧杆菌乳亚种Probio-M8的生长(P<0.05),与生长曲线结果一致。
![]() 图 6 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8活菌数的影响注:A. 益生菌菌落照片;B. 益生菌菌落数(*P<0.05)。Figure 6. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata myofibrillar protein on Probio-M8 viable bacteria count
图 6 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8活菌数的影响注:A. 益生菌菌落照片;B. 益生菌菌落数(*P<0.05)。Figure 6. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata myofibrillar protein on Probio-M8 viable bacteria count研究报道,双歧杆菌水解蛋白酶活性低以MRS作为培养基无法满足动物双歧杆菌乳亚种Probio-M8对肽及氨基酸营养需求,导致生长缓慢,因此,向培养基中补充氨基酸或肽可促进乳酸菌的增长[27]。本研究毛蚶蛋白为模拟消化产物,可能含有丰富的氨基酸,其可作为促进双歧杆菌增殖的生长因子。
2.5 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8胞外聚合物的影响
胞外聚合物(Extracellular Polymeric Substances,EPS)是由微生物分泌的一种聚合物,它包裹在细菌的外部,用于抵抗外界环境的压力和有害物质的侵害。利用扫描电镜观察毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8菌EPS的影响由图7所示。EPS表面呈颗粒状,整体上呈致密的网状结构,内部分布着许多大小形状相似的短杆菌;进一步放大可看出,EPS结构紧致光滑,动物双歧杆菌乳亚种Probio-M8被EPS包裹,形成复杂的生物膜。综上,毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8的EPS无明显影响。
![]() 图 7 毛蚶蛋白肌原纤维蛋白模拟消化产物对M8 胞外聚合物的影响Figure 7. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata on M8 extracellular polymeric substances
图 7 毛蚶蛋白肌原纤维蛋白模拟消化产物对M8 胞外聚合物的影响Figure 7. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata on M8 extracellular polymeric substances3. 结论
本文研究了植物乳植杆菌LP45、两歧双歧杆菌BBi32、鼠李糖乳酪杆菌LGG、动物双歧杆菌乳亚种Probio-M8四种益生菌对毛蚶肌原纤维蛋白模拟消化产物利用特性的影响。研究发现,毛蚶肌原纤维蛋白模拟消化产物在低浓度时对四种益生菌的生长均有一定的促进作用,高浓度会导致培养基渗透压增大,抑制微生物的生长且毛蚶肌原纤维蛋白模拟消化产物对不同益生菌的促进程度有显著差异,表明不同种类益生菌对氮源的需求不同。该研究可为益生菌的氮源生物利用提供理论指导。后续研究将聚焦于毛蚶肌原纤维蛋白模拟消化产物的分离纯化,探究不同益生菌对不同类型蛋白的偏好性。
-
![]()
图 1 毛蚶蛋白的蛋白质组成分析
注:A. 毛蚶蛋白的电泳图谱;B. 电泳条带灰度分析;不同小写字母代表两组差异显著(P<0.05)。
Figure 1. Analysis the protein composition of Arca subcrenata protein
![]()
图 2 四种益生菌在毛蚶肌原纤维蛋白模拟消化产物存在条件下的生长曲线及其积分面积
注:不同小写字母代表两组差异显著(P<0.05)。
Figure 2. Growth curves and area growth curves of four probiotics in the presence of simulated digestion products of Arca subcrenata myofibrillar protein
![]()
图 3 四种益生菌在不同浓度毛蚶肌原纤维蛋白模拟消化产物存在条件下的生长曲线及其积分面积
注:不同小写字母代表两组差异显著(P<0.05)。
Figure 3. Growth curves and area growth curves of four probiotics in the presence of different concentrations of simulated digestion products of Arca subcrenata myofibrillar protein
![]()
图 4 四种益生菌在不同浓度毛蚶肌原纤维蛋白模拟消化产物存在条件下的代时
注:不同小写字母代表两组差异显著(P<0.05)。
Figure 4. Generation time of four probiotics in the presence of different concentrations of simulated digestion products of Arca subcrenata myofibrillar protein
![]()
图 5 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8菌液pH的影响
Figure 5. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata myofibrillar protein on the pH of bacterial solution of Probio-M8
![]()
图 6 毛蚶肌原纤维蛋白模拟消化产物对动物双歧杆菌乳亚种Probio-M8活菌数的影响
注:A. 益生菌菌落照片;B. 益生菌菌落数(*P<0.05)。
Figure 6. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata myofibrillar protein on Probio-M8 viable bacteria count
![]()
图 7 毛蚶蛋白肌原纤维蛋白模拟消化产物对M8 胞外聚合物的影响
Figure 7. Effects of myofibrillar protein of simulated digestion products of Arca subcrenata on M8 extracellular polymeric substances
表 1 消化液电解质储备液的配制
Table 1 Preparation of electrolyte stock solutions of digestion fluids
盐溶液 储备液浓度
(mol/L)SSF(pH7)
1.25×(mmol/L)SGF(pH3)
1.25×(mmol/L)SIF(pH7)
1.25×(mmol/L)KCl 0.50 15.10 6.90 6.80 KH2PO4 0.50 3.70 0.90 0.80 NaHCO3 1.00 13.60 25.00 85.00 NaCl 2.00 − 47.20 38.40 MgCl·6H2O 0.15 0.15 0.12 0.33 (NH4)2CO3 0.50 0.06 0.50 − HCl 6.00 0.09 15.60 8.40 CaCl2·2H2O 0.30 1.50 0.15 0.60  下载: 导出CSV
下载: 导出CSV
-
[1] 储梦. 不同食源性蛋白对益生菌生长特性及肠道黏附的影响[D]. 扬州:扬州大学, 2022. [CHU Meng. Effects of different foodborne proteins on growth characteristics and intestinal adhesion of probiotics[D]. Yangzhou:Yangzhou University, 2022.] CHU Meng. Effects of different foodborne proteins on growth characteristics and intestinal adhesion of probiotics[D]. Yangzhou: Yangzhou University, 2022.
[2] ELLEN S M, J M D, GREGOR R, et al. Probiotics and prebiotics in intestinal health and disease:from biology to the clinic[J]. Nature Reviews Gastroenterology and Hepatology,2019,16(10):605−616. doi: 10.1038/s41575-019-0173-3
[3] 杨立娜, 吴凯为, 朱力杰, 等. 益生元、多酚、蛋白质和多不饱和脂肪酸对肠道健康的影响[J]. 食品工业科技,2017,38(22):336−340. [YANG Lina, WU Kaiwei, ZHU Lijie, et al. Effect of prebiotics, phytochemicals, protein and polyunsaturated fatty acids on intestinal health[J]. Science and Technology of Food Industry,2017,38(22):336−340.] YANG Lina, WU Kaiwei, ZHU Lijie, et al. Effect of prebiotics, phytochemicals, protein and polyunsaturated fatty acids on intestinal health[J]. Science and Technology of Food Industry, 2017, 38(22): 336−340.
[4] BROWN T D, WHITEHEAD K A, MITRAGOTRI S. Materials for oral delivery of proteins and peptides[J]. Nature Reviews Materials,2020,5(7):127−148.
[5] PORTUNE K J, BEAUMONT M, DAVILA A-M, et al. Gut microbiota role in dietary protein metabolism and health-related outcomes:The two sides of the coin[J]. Trends in Food Science and Technology,2016,57:213−232. doi: 10.1016/j.jpgs.2016.08.011
[6] 矫春娜, 张健, 刘芳, 等. 体外模拟消化在水产品营养活性物质研究中的应用进展[J]. 食品工业科技,2023,44(8):421−428. [JIAO Chunna, ZHANG Jian, LIU Fang, et al. Research progress on the application of in vitro digestion model in nutritional active substances of aquatic products[J]. Science and Technology of Food Industry,2023,44(8):421−428.] JIAO Chunna, ZHANG Jian, LIU Fang, et al. Research progress on the application of in vitro digestion model in nutritional active substances of aquatic products[J]. Science and Technology of Food Industry, 2023, 44(8): 421−428.
[7] 杜越欧, 侯俊财. 乳酸菌蛋白水解体系及相关基因表达的研究进展[J]. 食品工业科技, 2013, 34(3):383-386, 391. [DU Yueou, HOU Juncai. Research progress in proteolysis system of lactic acid bacteria and related gene expression[J]. Science and Technology of Food Industry, 2013, 34(3):383-386, 191.] DU Yueou, HOU Juncai. Research progress in proteolysis system of lactic acid bacteria and related gene expression[J]. Science and Technology of Food Industry, 2013, 34(3): 383-386, 191.
[8] CHI Z, SHAOQI X, YINXIAO Z, et al. Identification of soybean peptides and their effect on the growth and metabolism of Limosi Lactobacillus reuteri LR08[J]. Food Chemistry,2022,369:130923−130923. doi: 10.1016/j.foodchem.2021.130923
[9] MAKAROVA K S, KOONIN E V. Evolutionary genomics of lactic acid bacteria. Journal of bacteriology[J]. Journal of Bacteriology,2007,189(4):1199−1208. doi: 10.1128/JB.01351-06
[10] MARTONE C B, BRLA O P, SáNCHEZ J J. Fishery by-product as a nutrient source for bacteria and archaea growth media[J]. Bioresource Technology,2005,96(3):383−387. doi: 10.1016/j.biortech.2004.04.008
[11] AN C, KUDA T, YAZAKI T, et al. Caecal fermentation, putrefaction and microbiotas in rats fed milk casein, soy protein or fish meal[J]. Applied Microbiology and Biotechnology,2014,98:2779−2787. doi: 10.1007/s00253-013-5271-5
[12] LIU G Y, CHU M, XU P, et al. Effects of Ilisha elongata proteins on proliferation and adhesion of Lactobacillus plantarum[J]. Food Chemistry X, 2022,13:100206.
[13] 于志鹏, 武思佳, 赵文竹, 等. 海洋贝类蛋白源生物活性肽及肽组学的研究进展[J]. 食品工业科技,2015,26(22):384−388. [YU Zhipeng, WU Sijia, ZHAO Wenzhu, et al. Research progress on the study of bioactive peptides from shellfish proteins and peptidomics[J]. Science and Technology of Food Industry,2015,26(22):384−388.] YU Zhipeng, WU Sijia, ZHAO Wenzhu, et al. Research progress on the study of bioactive peptides from shellfish proteins and peptidomics[J]. Science and Technology of Food Industry, 2015, 26(22): 384−388.
[14] 李春磊, 朱建华, 宋丽艳, 等. 药用蚶的活性蛋白成分及药理作用研究[J]. 中国生化药物杂志,2016,36(10):155−160,164. [LI Chunlei, ZHU Jianhua, SONG Liyan, et al. Studies on bioactive proteins and pharmacological activities of Arca[J]. Chinese Journal of Biochemical Pharmaceutics,2016,36(10):155−160,164.] LI Chunlei, ZHU Jianhua, SONG Liyan, et al. Studies on bioactive proteins and pharmacological activities of Arca[J]. Chinese Journal of Biochemical Pharmaceutics, 2016, 36(10): 155−160,164.
[15] 梁铮洋, 周苗, 陶春霖, 等. 毛蚶蛋白对益生菌抗生素胁迫的保护作用[J]. 现代食品科技,2023,39(12):1−8. [LIANG Z Y, ZHOU M, TAO C L, et al. Protective effect of Arca subcrenata protein on probiotics against antibiotic damage[J]. Modern Food Science and Technology,2023,39(12):1−8.] LIANG Z Y, ZHOU M, TAO C L, et al. Protective effect of Arca subcrenata protein on probiotics against antibiotic damage[J]. Modern Food Science and Technology, 2023, 39(12): 1−8.
[16] KAUSHALYA C M, WONKYO J, JAEYOUNG J. Anti-inflammatory action of ark shell (Scapharca subcrenata) protein hydrolysate in LPS-stimulated RAW264.7 murine macrophages[J]. Journal of Food Biochemistry,2022,46(12):e14493.
[17] JIN J, AHN C, JE J. Purification and characterization of antioxidant peptides from enzymatically hydrolyzed ark shell (Scapharca subcrenata)[J]. Process Biochemistry, 2018:72170−72176.
[18] GUO Z, SHI H, LI C, et al. Identification and characterization of a novel protein ASP-3 purified from Arca subcrenata and its antitumor mechanism[J]. Marine Drugs,2019,17(9):528−528. doi: 10.3390/md17090528
[19] ANDRÉ B, LOTTI E, MARIE A, et al. INFOGEST static in vitro simulation of gastrointestinal food digestion[J]. Nature Protocols,2019,14(4):991−1014. doi: 10.1038/s41596-018-0119-1
[20] JIN D L, WEI J L, HE F Y, et al. Effect of tea polyphenols on sturgeon myofibrillar protein structure in the in vitro anti-glycation model mediated by low temperature vacuum heating food[J]. Chemistry,2023,407:135133.
[21] GOMES M A, MALCATA X F, KLLAVER A F. Growth enhancement of Bifidobacterium lactis Bo and Lactobacillus acidophilus Ki by milk hydrolyzates[J]. Journal of Dairy Science,1998,81(11):2817−2825. doi: 10.3168/jds.S0022-0302(98)75840-0
[22] ZHAO H, BAI F, ZHOU F, et al. Characterization of soybean protein hydrolysates able to promote the proliferation of Streptococcus thermophilus ST[J]. Journal of Food Science,2013,78(4):575−581.
[23] SU Y, SEGUINOT P, SANCHEZ I, et al. Nitrogen sources preferences of non- Saccharomyces yeasts to sustain growth and fermentation under winemaking conditions[J]. Food Microbiology,2020,85(C):103287.
[24] 向雄, 廖林锋, 王敏, 等. 竹虫蛋白酶解产物对肠道益生菌增殖作用的影响[J]. 食品研究与开发,2022,43(14):62−69. [XIANG Xiong, LIAO Linfeng, WANG Min, et al. Effects of protease hydrolysates of Chilo frusciden-talis Hampson on proliferation of intestinal probiotics[J]. Food Research and Development,2022,43(14):62−69.] XIANG Xiong, LIAO Linfeng, WANG Min, et al. Effects of protease hydrolysates of Chilo frusciden-talis Hampson on proliferation of intestinal probiotics[J]. Food Research and Development, 2022, 43(14): 62−69.
[25] ESTEFANIA V, JOSE M F. Effect of Inulin on the viability of L. plantarum during storage and in vitro digestion and on composition parameters of vegetable fermented juices[J]. Plant Foods for Human Nutrition (Dordrecht, Netherlands),2017,72(2):161−167. doi: 10.1007/s11130-017-0601-x
[26] 王元弛, 姚国强, 张文羿, 等. 乳双歧杆菌Probio-M8氧气耐受性驯化研究[J]. 中国食品学报,2020,20(12):1−7. [WANG Yuanchi, YAO Guoqiang, ZHANG Wenyi, et al. Studies on oxygen-tolerant domestication of Bifidobacterium animalis subsp. lactis Probio-M8[J]. Journal of Chinese Institute of Food Science and Technology,2020,20(12):1−7.] WANG Yuanchi, YAO Guoqiang, ZHANG Wenyi, et al. Studies on oxygen-tolerant domestication of Bifidobacterium animalis subsp. lactis Probio-M8[J]. Journal of Chinese Institute of Food Science and Technology, 2020, 20(12): 1−7.
[27] SAIJA N, WELMAN D A, BENNETT J R. Development of a dairy-based exopolysaccharide bioingredient[J]. International Dairy Journal,2010,20(9):603−608. doi: 10.1016/j.idairyj.2010.03.011
 下载:
下载:









计量
- 文章访问数: 76
- HTML全文浏览量: 14
- PDF下载量: 16



