


Effect of Chlorophylls on the Bioaccessibility of Lutein
-
摘要: 叶黄素具有多种生理活性,它在人体内不能合成,只能从外界获取,但叶黄素因其脂溶性的特点导致生物可给率低,如何高效补充叶黄素成为研究热点。叶绿素常与叶黄素共同出现在人类饮食中,且二者均为脂溶性物质,会产生一定的相互作用。因此研究膳食叶绿素及其结构变化对叶黄素生物可给率的影响具有重要意义。通过制备膳食中常见的8种不同结构的叶绿素(叶绿素a和b、脱镁叶绿素a和b、脱植基叶绿素a和b、脱镁叶绿酸a和b),采用体外静态消化模型和胶束化实验研究它们对叶黄素生物可给率的影响,并测定叶黄素和叶绿素形成的混合胶束粒径和电位的变化。结果表明,在共消化时不同结构的叶绿素能够显著提高叶黄素的回收率和生物可给率(P<0.05),其中脱镁叶绿素b的作用最为显著,所对应的叶黄素回收率和生物可给率分别为90.48%和80.44%。在消化过程中,未检测到明显的叶黄素降解产物,而叶绿素的结构则发生了明显的变化。通过消化物和胶束的平均粒径、荧光图像以及Zeta电位值可以确定胶束液体系比消化液体系更稳定,并且叶绿素可以通过与叶黄素形成复合物来保护叶黄素。本研究对如何提高叶黄素的生物可给率及消化利用性能具有一定的指导意义。Abstract: Lutein has a variety of physiological activities, which can not be synthesized by human body and can only be obtained through diet. Dietary lutein shows low bioaccessibility due to its lipo-soluble nature and thus the efficient supplementation of lutein has become a research hotspot. Chlorophylls, another type of lipo-soluble phytochemicals, are always ingested along with lutein in human diet where interaction between chlorophylls and lutein may occur. Therefore, it is of great significance to study the effect of dietary chlorophylls compounds and their structural changes on the bioaccessibility of lutein. Based on this, in this paper, eight different structures of chlorophylls (chlorophyll a, chlorophyll b, pheophytin a, pheophytin b, chlorophyllide a, chlorophyllide b, pheophorbide a and pheophorbide b) were prepared, and their effects on lutein bioaccessibility and related indexes were studied by in vitro static digestion model and micellization investigations. Changes of particle size and zeta-potential of mixed micelles formed by lutein and chlorophylls were also determined. The results showed that all the chlorophylls structures significantly improved the recovery rate and bioaccessibility (P<0.05) from in vitro digestion of lutein. Among them, pheophytin b exerted the most significant influence, with the corresponding recovery rate and bioaccessibility index of lutein being 90.48% and 80.44%, respectively. During digestion, no significant lutein degradation products were detected, while the structures of chlorophylls underwent significant changes. It could be found that the micellar fraction was more stable than the digesta fraction by the comparison of average particle size, fluorescence images and zeta potential value. Chlorophylls could protect lutein from degradation during in vitro digestion by forming some complex. This study provided important information on how to improve the bioaccessibility and digestion and utilization performance of dietary lutein.
-
Keywords:
- lutein /
- chlorophylls /
- interaction /
- bioaccessibility /
- in vitro digestion /
- micellization
-
叶黄素广泛存在于植物中,是一种优良的抗氧化剂。人类视网膜中含有大量的叶黄素,可在一定程度上过滤进入人眼视网膜的蓝光,有效降低视觉损害[1],还可以降低老年性黄斑变性的风险[2]。叶黄素结构中的C40类异戊二烯碳骨架和10个共轭双键使得叶黄素的溶解度较差,并且对光和氧非常敏感[3]。另外在胃肠道消化过程中也会发生降解,最终导致叶黄素在肠道中的吸收很少,生物可给率低。考虑到从食物中摄取是获得叶黄素的唯一来源,因此如何高效补充叶黄素成为研究热点。目前常采用递送系统(用脂质体、纳米颗粒、乳液、微胶囊等包埋)[4]来提高叶黄素的生物可给率,焦岩等[5]通过纳米脂质体的载体作用将叶黄素包囊形成纳米结构,改变其疏水性状态,能有效提高叶黄素的稳定性;王晓[6]采用微胶囊技术,制备高稳定性和高生物利用度的负载叶黄素微胶囊。此外,石晓晴等[7]还通过物理包埋和化学改性(如酯化反应)等方法改变剂型,并利用超微粉碎和微囊化等技术将叶黄素制备成油悬浮液、水分散性干粉、微胶囊和脂质体。
在消化过程中有许多因素会影响叶黄素的生物可给率,包括叶黄素本身的物理/化学性质、食物材料的微观结构、与其他食物成分的相互作用等[8]。研究表明叶黄素与部分食物成分的相互作用可有效防止其氧化降解[9],如多酚、脂质、膳食纤维等[10]。叶绿素与叶黄素共存于植物的叶绿体中,叶绿素也是一种脂溶性物质且结构多变,其分子由一个卟啉环和一个脂肪烃侧链(植醇)组成,卟啉环的中央含有一个镁原子[11]。叶绿素本身在光、热、酸等理化因素下极不稳定,易分解或氧化生成不同结构的衍生物,在酸性条件下叶绿素卟啉环中央的镁原子容易被氢离子取代生成脱镁叶绿素,而在碱性或酸性作用下叶绿素容易水解脱去植基变成脱植基叶绿素[12]。高等植物中绝大多数叶绿素为叶绿素a和叶绿素b,在酸、热等过程中会形成脱镁叶绿素a和脱镁叶绿素b,同时在可食用海藻如紫菜、海带等中还存在大量的不含植醇链的叶绿素衍生物,如脱植基叶绿素和脱镁叶绿酸[13]。目前国内外关于叶绿素与其他脂溶性成分相互作用的研究较少,而大多集中在叶绿素的代谢[14]、脂质的添加对叶绿素降解的作用[15]以及保护叶绿素不被氧化降解的措施[16]等。与叶黄素一样,叶绿素需要被掺入胶束后才能有效地进行肠吸收[17],然而关于膳食叶绿素是否影响叶黄素生物利用还知之甚少,因此研究叶绿素及其结构变化对叶黄素生物可给率的影响以及两者共消化及胶束化过程中产生的相互作用具有重要意义。
本研究制备了8种膳食中常见的叶绿素及其衍生物(叶绿素a和b、脱镁叶绿素a和b、脱植基叶绿素a和b、脱镁叶绿酸a和b,化学结构见图1和表1),利用体外静态消化模型及胶束化实验研究它们对叶黄素生物可给率的影响,为研究如何提高叶黄素的生物利用度提供一定的参考。
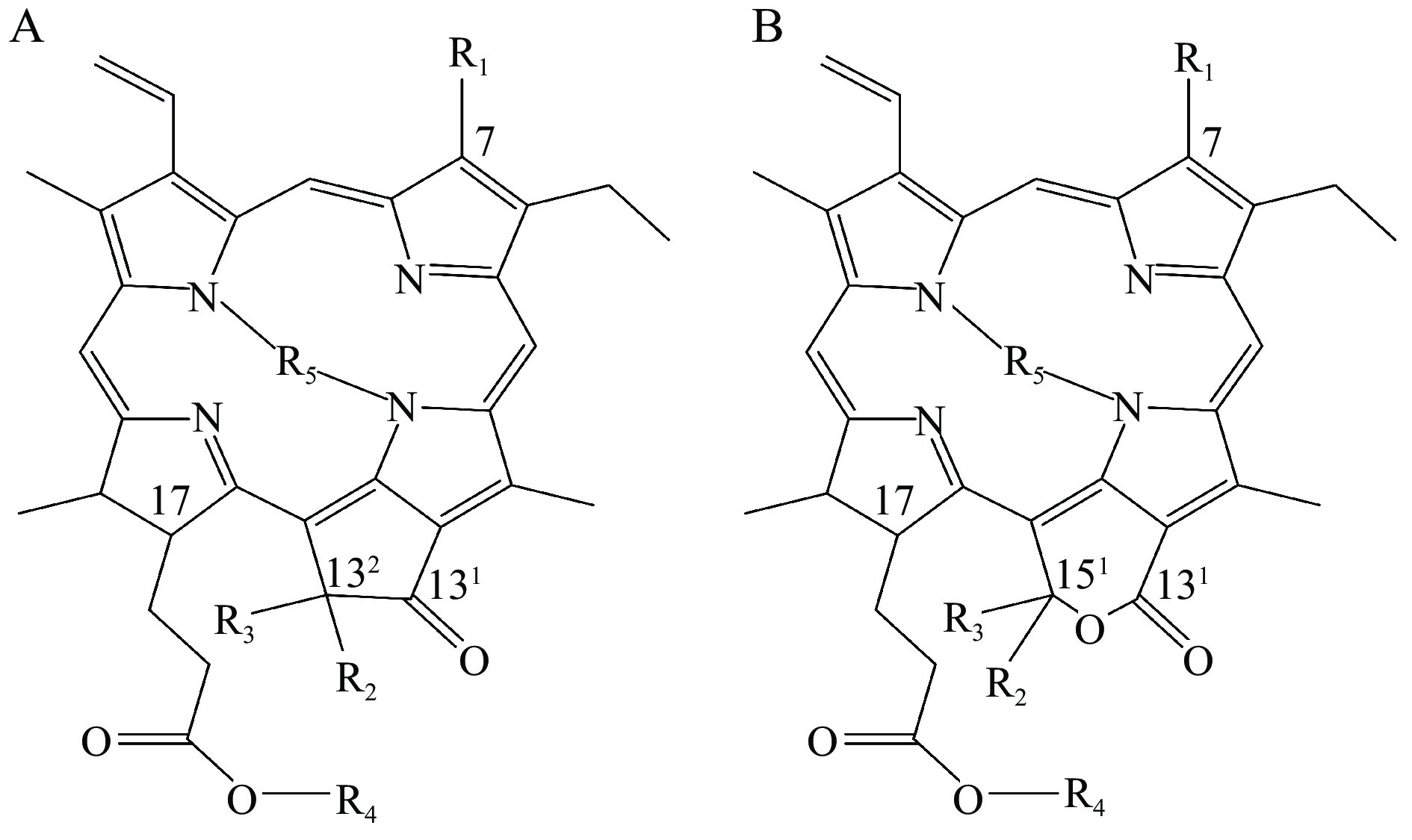
表 1 8种常见叶绿素及其衍生物的化学结构Table 1. Chemical structures of eight common chlorophylls and their derivatives名称 结构类型 R1 R2 R3 R4 R5 叶绿素a(Chlorophyll a) A CH3 H COOCH3 C20H39 Mg 132-羟基叶绿素a(132-Hydroxy chlorophyll a) A CH3 OH COOCH3 C20H39 Mg 151-羟基内酯叶绿素a(151-Hydroxy-lactone chlorophyll a) B CH3 OH (152)COOCH3 C20H39 Mg 叶绿素b(Chlorophyll b) A CHO H COOCH3 C20H39 Mg 132-羟基叶绿素b(132-Hydroxy chlorophyll b) A CHO OH COOCH3 C20H39 Mg 151-羟基内酯叶绿素b(151-Hydroxy-lactone chlorophyll b) B CHO OH (152)COOCH3 C20H39 Mg 脱镁叶绿素a(Pheophytin a) A CH3 H COOCH3 C20H39 2H 132-羟基脱镁叶绿素a(132-Hydroxy pheophytin a) A CH3 OH COOCH3 C20H39 2H 151-羟基内酯脱镁叶绿素a(151-Hydroxy-lactone pheophytin a) B CH3 OH (152)COOCH3 C20H39 2H 脱镁叶绿素b(Pheophytin b) A CHO H COOCH3 C20H39 2H 132-羟基脱镁叶绿素b(132-Hydroxy pheophytin b) A CHO OH ZZCOOCH3 C20H39 2H 脱植基叶绿素a(Chlorophyllide a) B CHO OH (152)COOCH3 C20H39 2H 132-羟基脱植基叶绿素a(132-Hydroxy chlorophyllide a) A CH3 H COOCH3 H Mg 151-羟基内酯脱植基叶绿素a(151-Hydroxy-lactone chlorophyllide a) A CH3 OH COOCH3 H Mg 脱植基叶绿素b(Chlorophyllide b) B CH3 OH (152)COOCH3 H Mg 132-羟基脱植基叶绿素b(132-Hydroxy chlorophyllide b) A CHO H COOCH3 H Mg 151-羟基内酯脱植基叶绿素b(151-Hydroxy-lactone chlorophyllide b) A CHO OH COOCH3 H Mg 脱镁叶绿酸a(Pheophorbide a) B CHO OH (152)COOCH3 H Mg 132-羟基脱镁叶绿酸a(132-Hydroxy pheophorbide a) A CH3 H COOCH3 H 2H 151-羟基内酯脱镁叶绿酸a(151-Hydroxy-lactone pheophorbide a) A CH3 OH COOCH3 H 2H 脱镁叶绿酸b(Pheophorbide b) B CH3 OH (152)COOCH3 H 2H 132-羟基脱镁叶绿酸b(132-Hydroxy pheophorbide b) A CHO H COOCH3 H 2H 151-羟基内酯脱镁叶绿酸b(151-Hydroxy-lactone pheophorbide b) A CHO OH COOCH3 H 2H 脱植基叶绿素a(Chlorophyllide a) B CHO OH (152)COOCH3 H 2H 1. 材料与方法
1.1 材料与仪器
新鲜菠菜、成熟酸橙(Citrus×aurantium(Lour.)Engl.) 重庆当地市场;长寿花金胚玉米油(1 L) 山东三星玉米产业科技有限公司;纯品叶黄素(CAS:127-40-2,纯度≥90%) 上海麦克林生化有限公司;乙醇(分析纯)、丙酮(分析纯)、无水硫酸钠(分析纯)、盐酸 重庆川东化工有限公司;N,N-二甲基甲酰胺 万盛川东化工有限公司;2,6-二叔丁基对甲酚(BHT) 阿拉丁生物化学技术有限公司;乙酸铵、丙酮(色谱纯)、甲醇(色谱纯)、猪胰腺α-淀粉酶(≥5 U/mg)、猪胰腺胰蛋白酶(4 U/mg)、猪胃粘膜胃蛋白酶(≥250 U/mg)、猪胰腺脂肪酶(≥125 U/mg)、猪胆汁提取物(≥200 U/mg) Sigma-Aldrich(中国上海);氯化钾、氯化钠、碳酸氢钠 成都科隆化工有限公司。
SPH-200B恒温摇床 上海世平实验设备有限公司;TGL-16M台式高速冷冻离心机 湖南湘仪实验室仪器开发有限公司;PB-10标准型PH计 赛多利斯科学仪器(北京)有限公司;BSA124S分析天平 德国Sartorious公司;KQ5200DE型数控超声波清洗器 昆山市超声仪器有限公司;SHZ-II型循环水真空泵、RE-52AA旋转蒸发器 上海亚荣生化仪器厂;DHG-9240A电热恒温鼓风干燥箱 上海齐欣科学仪器有限公司;Agilent 1290 infinity II超高效液相色谱仪 美国Agilent公司;XP-DCY-12SL圆形电动水浴氮吹仪 上海析谱仪器有限公司;DW-HL398超低温冷冻储存箱 中科美菱低温科技股份有限公司;Malvern Zetasizer Nano ZS90纳米粒度电位仪 英国马尔文仪器有限公司;Vortex-2涡旋混匀仪 上海沪析实业有限公司;BX53正置荧光显微镜 日本Olympus公司。
1.2 实验方法
1.2.1 8种不同结构叶绿素的制备
参考实验室已有的方法[18]提取叶绿素并进行一些修改,本实验中的所有操作均在安装有绿光灯且避光的实验室内进行,以避免色素的光降解。将新鲜菠菜叶片洗净、剪碎并打浆,与等比例的石油醚(30~60 ℃)进行充分混合,用八层纱布过滤,弃去含有叶黄素和少部分叶绿素的第一次滤液,加入等体积的丙酮:乙醇=1:1混合液提取滤渣中的叶绿素,重复操作直至绿色的菠菜滤渣转变为黄色。再用等体积的乙醚进行复提,经无水硫酸钠过滤后,通过旋转蒸发器在常温下减压旋转挥干乙醚及残留的水分,最后用少量丙酮溶解并经超声3 min后过滤(0.22 μm),即可得到叶绿素原液。将叶绿素原液均匀地点样于1 mm厚的滤纸板上,用配比为石油醚(60~90 ℃):乙醇:甲醇=60:20:1的层析液洗脱,进行薄层层析。层析产生的蓝绿色条带为叶绿素a,黄绿色条带为叶绿素b。将来自层析的叶绿素条切割并浸入等体积丙酮中以获得相应的叶绿素-丙酮溶液。
用植物组织制备的丙酮粉可以实现叶绿素的脱植基化,其中成熟的橙皮具有较高的叶绿素酶活性[18]。取成熟酸橙(C. aurantium(Lour.)Engl.)的外皮并用蒸馏水清洗,沥干水分后放入−20 ℃冰箱中储存,使用时将酸橙皮剪碎后进行破碎,加入等比例的冷丙酮(−20 ℃)进行混匀,抽滤去除丙酮并置于通风橱内晾干,之后将其研磨并用30目网筛进行过筛,即可得到橙皮丙酮粉。称取4 g橙皮丙酮粉用30 mL,5 mmol/L的PBS(pH7.0,50 mmol/L KCl,0.24% Trition-X-100)提取叶绿素酶,经磁力搅拌器700 r/min搅拌1 h后用八层纱布过滤,滤液经离心机5000 r/min离心8 min后,取上清液备用。将50 μL,1 μmol/mL的叶绿素与0.2 mL酶液以及0.2 mL,0.1 mol/L PBS(pH7.0,50 mmol/L MgCl2)混合并搅拌均匀。将叶绿素b反应液在摇床上于30 ℃下酶解1 h,将叶绿素a反应液在摇床上于40 ℃下酶解3 h。反应结束后加入4倍体积的丙酮终止其反应,反应液用等体积乙醚复提,经无水硫酸钠除水后,进行常温旋转挥干,用少量丙酮溶解,得到的叶绿素丙酮溶液采用纸层析法可制备得到脱植基叶绿素a、b溶液。
将5 mL,1 µmol/mL的叶绿素a/b或脱植基叶绿素a/b丙酮溶液置于50 mL离心管中,滴入2~3滴0.1 mol/L的盐酸溶液。不断摇晃观察到液体颜色由绿变褐后迅速加入纯水20 mL,用乙醚复提。向有机相中加入等体积的10%氯化钠水溶液,洗去有机相中的氢离子,重复3~4次。将有机相减压旋转挥干,并用少量丙酮溶液进行溶解,即可得到脱镁叶绿素a/b或脱镁叶绿酸a/b溶液。
1.2.2 标准曲线的建立
参考实验室已有的研究[19]和方法[20]并加以修改,对1.2.1中所制备的8种叶绿素溶液的具体浓度进行测定,同时用叶黄素标准品配制摩尔浓度分别为0.01、0.1、1.0 μmol/mL的叶黄素-乙醇溶液,均采用与光电二极管阵列检测器相偶联的超高效液相色谱法(ultra-high performance liquid phase chromatography-diode array detector,UPLC-DAD),配备有自动进样系统。分析柱为C18柱(Poroshell 120,1.9 μm,2.1×100 mm),并配有同等材料的保护柱。流动相A为甲醇:水=8:2混合液,流动相B为甲醇:丙酮=1:1混合液,向两流动相中分别加入10 mmol/L乙酸铵,流速为0.2 mL/min。使用梯度洗脱程序:0~15 min,0%~100% B;15~20 min,100% B;20~25 min,0% B。最终根据保留时间及光谱特征对叶黄素和叶绿素进行定性分析,得到的不同色素浓度的标准曲线方程结果见表2。
表 2 不同色素浓度的标准曲线方程Table 2. Standard curve equations for different pigments concentrations色素种类 DAD检测波长
(nm)标准曲线方程 决定系数
(R2)叶黄素 445 y=0.000315x−0.001214 0.9993 叶绿素a 664 y=0.000006x−0.002447 0.9969 叶绿素b 647 y=0.000005x+0.000973 0.9979 脱镁叶绿素a 408 y=0.000005x−0.003201 0.9935 脱镁叶绿素b 436 y=0.000003x+0.010738 0.9937 脱植基叶绿素a 664 y=0.000006x−0.002447 0.9969 脱植基叶绿素b 647 y=0.000005x+0.000973 0.9979 脱镁叶绿酸a 408 y=0.000003x−0.004508 0.9989 脱镁叶绿酸b 436 y=0.000003x−0.006572 0.9992 注:标准曲线方程中y为色素的浓度(μmol/mL),x为峰面积。 1.2.3 体外静态消化
参考标准植物化学物质的体外消化实验方法[19,21−23],模拟口腔、胃和肠道三个阶段的消化过程,设置三个消化实验组:叶黄素和叶绿素共消化、叶绿素单独消化及叶黄素单独消化。根据相关研究及人类日常膳食摄入量标准,类胡萝卜的日均摄入量为14 mg[24],总叶绿素的日均摄入量为50~65 mg[25],以此确定叶黄素体外消化的起始浓度为2.5~50 μmol/L,叶绿素的起始浓度应为相应叶黄素的3.5~4.5倍,最终确定本研究中叶黄素的初始浓度为7.5 μmol/L,而叶绿素的初始浓度为叶黄素的4.2倍,即31.5 μmol/L,并按照最终消化液体积为10 mL来计算所需的叶黄素和叶绿素贮备液体积。每组均设置6次独立平行实验。
前处理:分别将叶黄素和不同结构的叶绿素按照比例单独或共同加到15 mL样品管中,进行氮吹晾干后加入50 μL玉米油,将其在功率为200 W的超声波清洗器中超声10 min。其中玉米油为精炼油,以减少油中含有的少量叶黄素对反应体系的影响。
口腔阶段:将0.568 g α-淀粉酶溶于生理盐水(140 mmol/L NaCl和5 mmol/L KCl)中,最终体积为25 mL作为模拟唾液。向每个样品管中加入750 μL模拟唾液和6.5 μL,0.3 mmol/L CaCl2溶液,用0.9 mol/L NaHCO3溶液将pH调至7.0,补加生理盐水使总体积为2.5 mL,37 ℃恒温摇床中85 r/min反应2 min。
胃消化阶段:将0.496 g胃蛋白酶溶于生理盐水(140 mmol/L NaCl和5 mmol/L KCl)中,最终体积为25 mL作为模拟胃液。向每个口腔阶段消化结束后的样品管中加入750 μL模拟胃液和2.5 μL,0.3 mmol/L CaCl2溶液,用5 mmol/L HCl溶液将pH调至3.0,补加生理盐水使总体积为5 mL,37 ℃恒温摇床中85 r/min反应2 h。
肠消化阶段:将8.333 g胰蛋白酶、5 g胆盐和2.222 g胰脂肪酶溶于0.1 mol/L NaHCO3溶液中,最终体积为125 mL作为模拟肠液。首先用0.9 mol/L NaHCO3溶液将pH调至6.5以终止胃消化,再向每个胃消化阶段结束后的样品管中加入3.75 mL模拟肠液和10 μL,0.3 mmol/L CaCl2溶液,之后用0.9 mol/L NaHCO3溶液将pH调至7.0,补加生理盐水使总体积为10 mL,其它后续操作同上。
1.2.4 体外离心共胶束化
体外消化完成后,分别取1 mL消化液于15、2.5 mL样品管,并分别置于−20 ℃(用于色素提取)和4 ℃(用于电位和粒径检测)下待用以抑制所有消化酶的作用。另取5 mL消化液于15 mL加厚离心管,置于离心机中,在4 ℃、12000 r/min条件下离心1.5 h。离心完成后用0.2 μm尼龙膜过滤,得到混合胶束,分别取1 mL胶束液于15、2.5 mL样品管,并分别置于−20 ℃(用于色素提取)和4 ℃(用于电位和粒径检测)下待用。
1.2.5 从消化液和胶束中提取色素
以消化液为例,向消化液中加入2 mL饱和食盐水和2 mL丙酮(含0.1% BHT),摇匀后加入2 mL乙醚提取有机相,吸取上层有机相于另一干净样品管,重复操作(至少3次)直至上层有机相变为无色,通过旋转蒸发器在常温下减压旋转挥干有机溶剂及残留水分,最后用1 mL丙酮溶解粘附在蒸发瓶上的色素,涡旋2 min后用0.22 μm尼龙膜过滤,用于超高效液相色谱分析,并根据1.2.2中建立的标准曲线对消化液及胶束液中的叶黄素和叶绿素进行定量测定。
1.2.6 Zeta电位和粒径的测定
对经过体外消化后消化液和胶束液的粒径和Zeta电位进行测定。分别将经体外消化后置于4 ℃下的消化液和胶束液进行样品制备,采用纳米粒度电位仪分别测定Zeta电位和平均粒径,使用纯水作为分散体系,折射率为1.33。每个样品进行6次独立重复测定。
1.2.7 荧光显微镜观察消化液和胶束液形貌
叶绿素会吸收短波长的激发光,发射出更大波长的光作为荧光[26],据此可以利用叶绿素荧光的特性,将与叶黄素共消化或单独消化后叶绿素在消化液和胶束液中的分布可视化。参考实验室已有的方法[19,21]并加以改进,将叶绿素与叶黄素共消化或单独消化后的少量消化液和胶束液分别均匀地涂布在载玻片上,用盖玻片覆盖。使用的激发射线波长为633 nm,收集的发射荧光波长范围为650~700 nm[27],用正置荧光显微镜以10×的放大率在荧光模式下观察样品。每个样品进行3次独立重复测定。
1.2.8 消化特性指标的计算
体外消化特性指标的相关计算如下:
回收率(%)=消化液中色素的质量(µg)初始色素的质量(µg)×100 胶束化率(%)=胶束中色素的质量(µg)消化液中色素的质量(µg)×100 生物可给率(%)=胶束中色素的质量(µg)初始色素的质量(µg)×100 1.3 数据处理
本研究利用超高效液相色谱配备的Bruker Compass Data Analysis 5.1数据处理软件对体外消化前后不同结构的叶绿素及其衍生物和叶黄素进行定性定量分析,采用Origin 2022软件对数据进行绘图,运用SPSS 26.0统计分析软件进行单因素方差分析和t检验,P<0.05表示具有显著性差异。每组数据均进行6次独立平行实验,最终结果表示为平均值±标准差。
2. 结果与分析
2.1 体外消化过程中叶黄素和叶绿素各项指标的变化
2.1.1 回收率分析
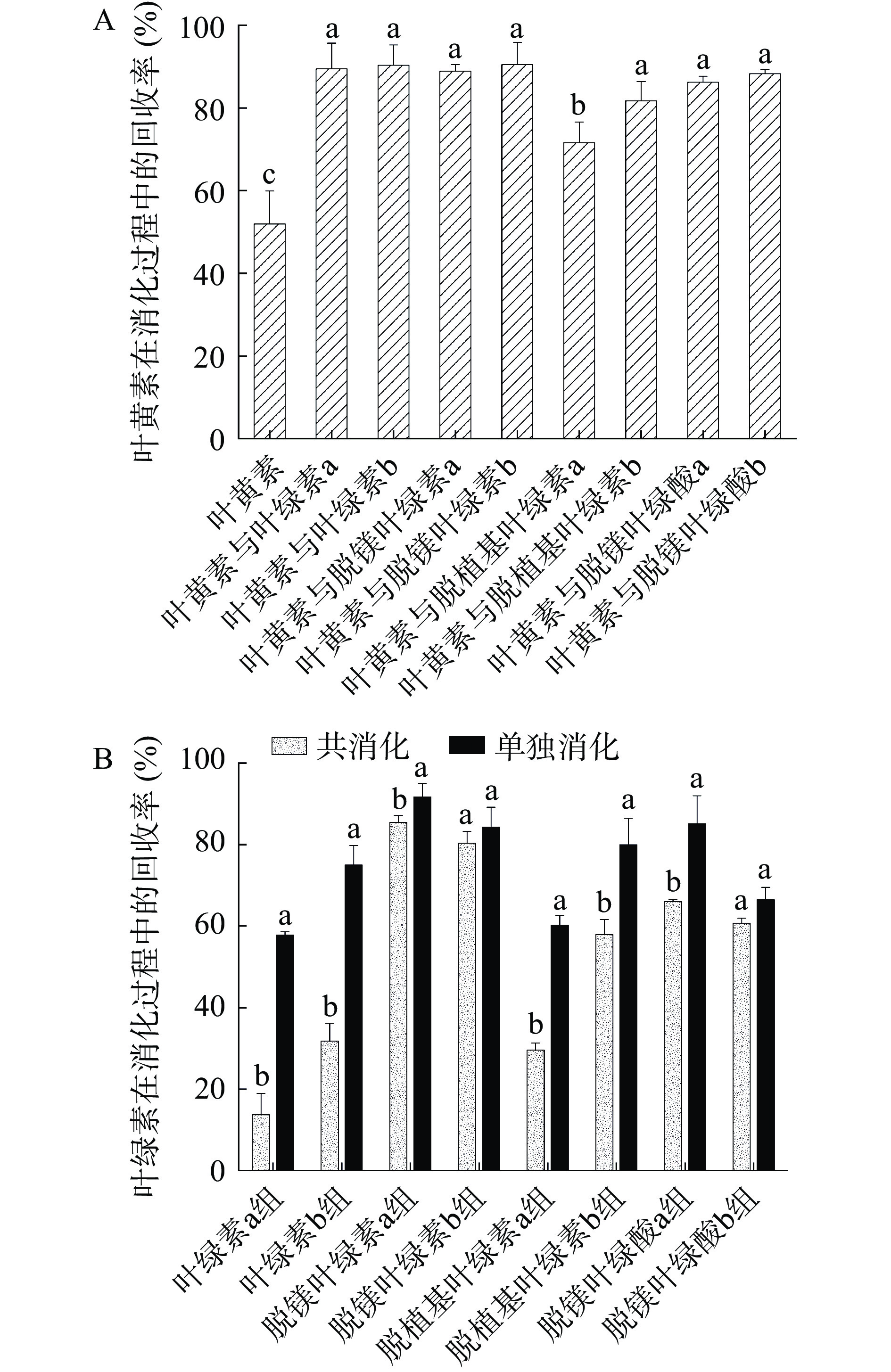
回收率能够反映体外消化后叶黄素和叶绿素被消化降解为无色物质而损失的程度[21],图2(A)为叶黄素的回收率,其中叶黄素单独消化时的回收率最低(51.91%),与不同结构叶绿素共消化时叶黄素的回收率均升高,且与单独消化时相比具有显著差异(P<0.05),说明不同结构叶绿素均能对叶黄素的消化损失起到一定的抑制作用。其中脱镁叶绿素b对叶黄素的保护作用最强,两者共消化时叶黄素的回收率高达90.48%,而脱植基叶绿素a对叶黄素的保护作用最弱(71.60%)。图2(B)为叶绿素及其衍生物的回收率,其中脱镁叶绿素a组在单独消化及与叶黄素共消化时的回收率均最大,分别为91.67%和85.45%。初始投入叶绿素a、b进行消化,最终叶绿素b组的回收率总体上高于叶绿素a组,与之前的研究结果相一致[23],这是因为在结构上叶绿素 b 中 C7 位上的-CHO 比叶绿素 a 中的 -CH3 更有助于稳定叶绿素的卟啉结构[21]。
![]()
2.1.2 胶束化率分析
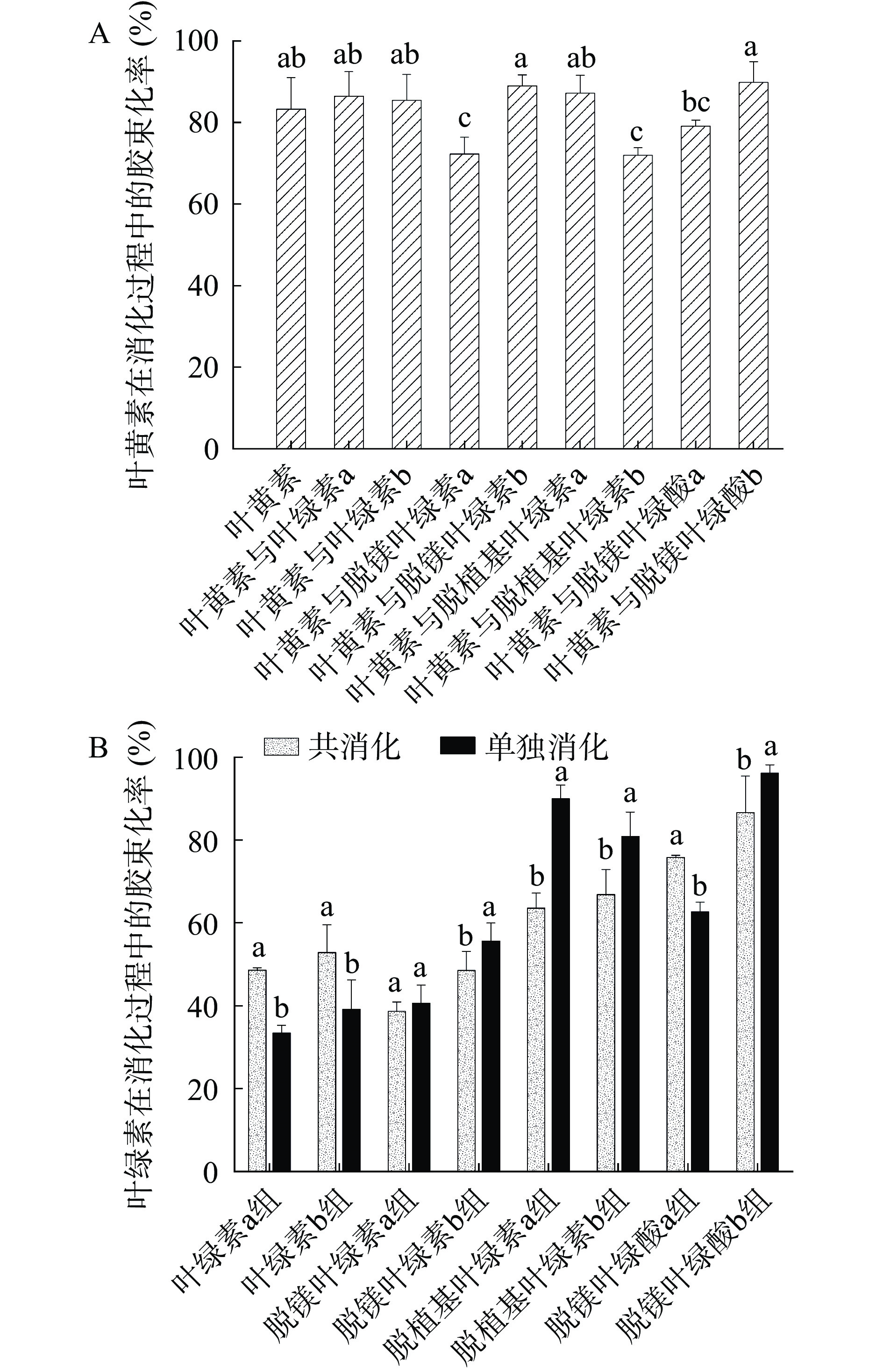
混合胶束由胆盐和脂肪消化产物等形成[28],胶束化率是评价脂溶性化合物能被肠上皮细胞吸收的有效指标[21]。图3(A)为叶黄素的胶束化率,叶黄素单独消化时的胶束化率可达83.27%,这是因为叶黄素本身的极性较大,很容易通过脂滴表面进入小肠内与胆盐等其他物质形成混合胶束[29]。在共消化时,脱镁叶绿素a、脱植基叶绿素b以及脱镁叶绿酸a均使叶黄素的胶束化率降低,可能原因是这三种叶绿素与叶黄素在胶束化过程中发生复合形成了致密的结构而导致叶黄素胶束化率降低[21]。而其他结构的叶绿素则使叶黄素的胶束化率升高,其中脱镁叶绿酸b所对应的叶黄素胶束化率最大,为89.80%。图3(B)为叶绿素及其衍生物的胶束化率,脱镁叶绿酸b组在单独消化及与叶黄素共消化时均呈现出最大的胶束化率,分别为96.16%和86.64%,且两者间具有显著性差异(P<0.05)。脱镁叶绿酸a组的胶束化率总体上低于脱镁叶绿酸b组,与先前的研究[17,21]不一致,可能是受到食物基质及消化环境条件等的影响。大部分结构的叶绿素在总体上均呈现b系列比a系列更利于胶束化,这与之前的研究结果[19]相一致,可能原因是b系列叶绿素具有能增强亲水性的甲酰基[19]。脱植基叶绿素a、b组和脱镁叶绿酸a、b组的胶束化率总体上比叶绿素a、b组和脱镁叶绿素a、b组高,即无植醇链叶绿素的胶束化率较含植醇链叶绿素的高,这是因为植醇的存在能够影响胶束化过程,并且极性叶绿素比非极性叶绿素优先胶束化和被吸收[17]。另外,叶绿素a、b组的胶束化率与脱镁叶绿素a、b组的差异不大,可能是因为中心镁的存在与否对掺入胶束的影响并不显著[17]。
![]() 图 3 体外消化后叶黄素和叶绿素及其衍生物的胶束化率Figure 3. Micellization rate of lutein, chlorophylls and their derivatives after in vitro digestion
图 3 体外消化后叶黄素和叶绿素及其衍生物的胶束化率Figure 3. Micellization rate of lutein, chlorophylls and their derivatives after in vitro digestion2.1.3 生物可给率分析
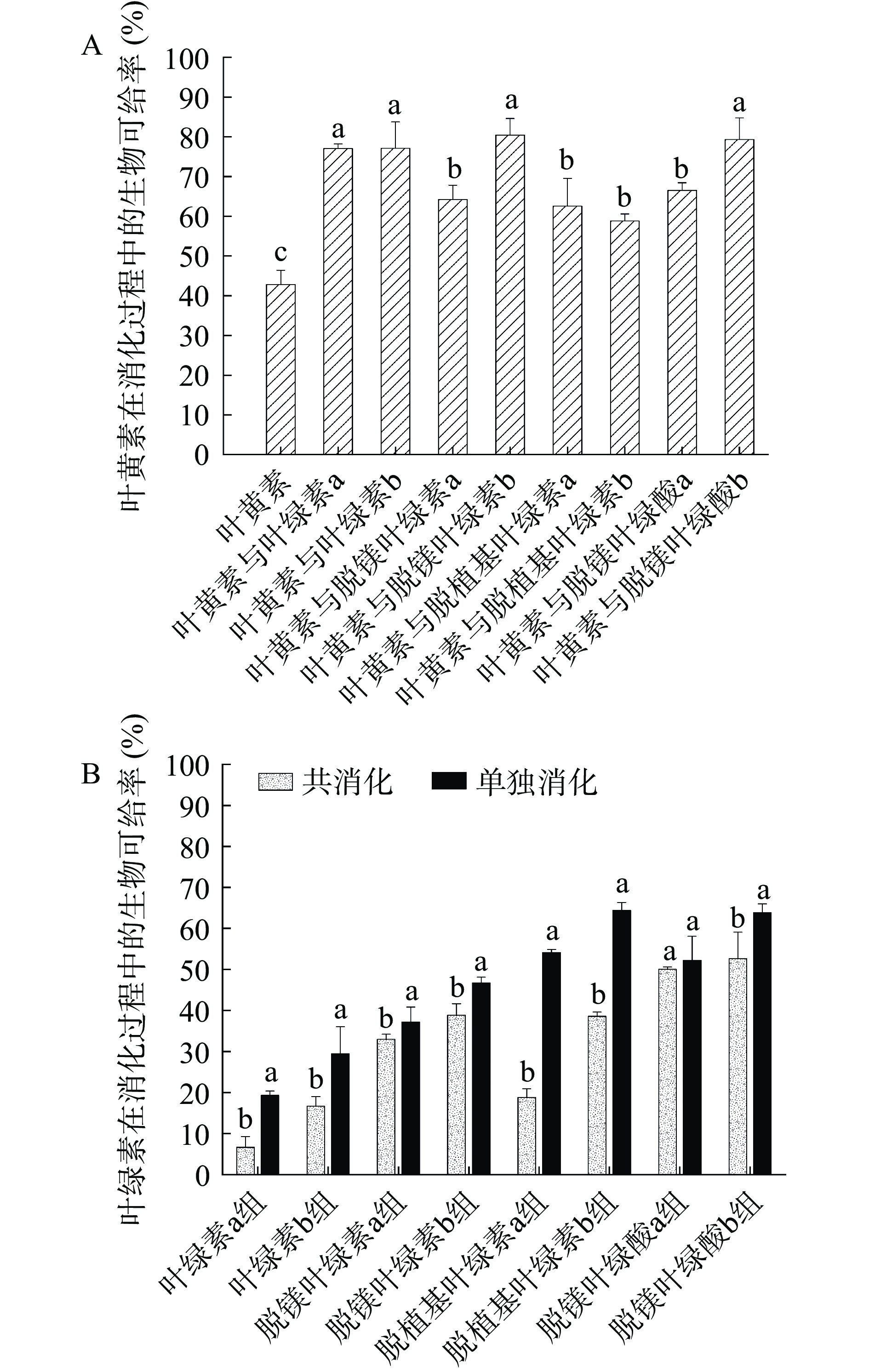
生物可给率是对营养物质在摄入后能够被吸收利用程度的综合评价,可用来评估消化期间从食物基质转移到胶束的摄入化合物的量[23]。图4(A)为叶黄素的生物可给率,可知叶黄素单独消化时其生物可给率最小(42.83%),与不同结构叶绿素共消化时的叶黄素生物可给率均增大,且两者之间均具有显著差异(P<0.05),说明不同结构的叶绿素均能提高叶黄素的生物利用程度。其中脱镁叶绿素b与叶黄素共消化时所对应的叶黄素生物可给率最大,为80.44%。结合叶黄素的回收率(图2A)和胶束化率(图3A)可知,在确定叶黄素生物可给率方面的主导因素是回收率,这是因为相比于胶束化过程,叶绿素能更多地在消化过程中保护叶黄素不被破坏。图4(B)为叶绿素及其衍生物的生物可给率,可知与叶黄素共消化时叶绿素组的生物可给率均更低,可能原因是叶黄素从食物基质中释放进入胃肠环境后,被油脂包埋其中,必须将表层脂质水解才能使叶黄素暴露[10],而在与叶绿素共消化时,叶绿素的脂溶性特性能抑制其表层脂质水解以减少叶黄素的暴露及消解,同时叶绿素还具有一定的抗氧化性[30],在消化过程中可抑制叶黄素被氧化,从而提高叶黄素在体内的生物可给率。其中单独消化时脱植基叶绿素b组的生物可给率最大,为64.40%;与叶黄素共消化时脱镁叶绿酸b组的生物可给率最大,为52.65%。b系列叶绿素的生物可给率均比a系列的大,是因为b系列叶绿素在总体上具有更大的胶束化率,而生物可给率会受到胶束化过程的影响。另外,在总体上呈现出无植醇链叶绿素(脱植基叶绿素a和b组、脱镁叶绿酸a和b组)的生物可给率较含植醇链叶绿素(叶绿素a和b组、脱镁叶绿素a和b组)的高,这与叶绿素胶束化率(图3B)的趋势一致,可能原因是叶绿素的脱植醇反应可显著提高其胶束化能力,从而进一步提高体外消化过程中的生物可给率[18]。
![]() 图 4 体外消化后叶黄素和叶绿素及其衍生物的生物可给率Figure 4. Bioaccessibility index of lutein, chlorophylls and their derivatives after in vitro digestion
图 4 体外消化后叶黄素和叶绿素及其衍生物的生物可给率Figure 4. Bioaccessibility index of lutein, chlorophylls and their derivatives after in vitro digestion2.2 叶绿素及其衍生物在体外消化过程中的组成变化
表3为单独消化或与叶黄素共消化前后叶绿素及其衍生物的组成变化。叶黄素在体外消化前后的结构未发生变化,而叶绿素在食品加工和其他条件下容易发生氧化,形成各种叶绿素氧化产物,如132-羟基衍生物和151-羟基内酯衍生物等[31],在本研究中这两类叶绿素氧化产物均能被检测到,这与体外消化过程中的氧化环境有关[23]。胃中的酸性条件会将叶绿素a和叶绿素b分别转化为脱镁叶绿素a和脱镁叶绿素b,而玉米油对此具有一定的保护作用,会抑制部分叶绿素发生转化[23]。同时与单独消化相比,叶黄素共消化时叶绿素能更多地转化为脱镁叶绿素,这是因为在消化过程中叶黄素较叶绿素更加稳定,两者共存时叶绿素的结构会优先发生变化,说明共消化时叶绿素在一定程度上能为叶黄素提供保护作用。另外相比于叶绿素b组,叶绿素a组在消化过程中能更多地转变为脱镁叶绿素,并且先前的研究也表明在相同的消化时间内叶绿素a转化为脱镁叶绿素的速度比叶绿素b快[23]。脱镁叶绿素a、b组在消化前后均未转变为其他结构的叶绿素,只在自身内部发生结构变化。先前研究发现脱镁叶绿素的植基链可以水解形成脱镁叶绿酸[21,23],而在本研究中并未发现这种现象,可能原因是食物基质玉米油会对脱镁叶绿素的水解起到一定的抑制作用。在胃酸的作用下,脱植基叶绿素可以转化为脱镁叶绿酸[17],本研究中发现脱植基叶绿素a、b组中均有这样的变化。有研究表明氧化的叶绿素衍生物具有更高的胶束化率,并且羟基的增加有助于提高其在胶束中的溶解度[32],本研究中脱镁叶绿素a和b组、脱植基叶绿素a和b组以及脱镁叶绿酸b组中均具有这样的趋势,与叶黄素共消化相比,它们单独消化后能产生更多的132-羟基衍生物或151-羟基内酯衍生物(表3),所对应的胶束化率(图3B)也在单独消化时比与叶黄素共消化时更高。
表 3 消化前后叶绿素及其衍生物的组成含量变化(%)Table 3. Changes in the composition contents of chlorophylls and their derivatives before and after digestion (%)叶绿 素种类 消化前 消化后 叶绿素单独消化 叶绿素与叶黄素共消化 叶绿素a组 叶绿素a 91.02±0.82 30.49±5.21 叶绿素a' 5.81±0.51 7.18±1.22 132-羟基叶绿素a 0.76±0.10 2.23±0.26 151-羟基内酯叶绿素a 2.41±0.54 0.64±0.30 脱镁叶绿素a 48.92±5.25 75.92±0.76 脱镁叶绿素a' 8.24±0.91 9.57±0.35 132-羟基脱镁叶绿素a 2.04±0.29 14.50±0.41 151-羟基内酯脱镁叶绿素a 0.26±0.01 叶绿素a系列 100.00 40.54±6.44 脱镁叶绿素a系列 59.46±6.44 100.00 叶绿素b组 叶绿素b 81.12±1.21 64.46±1.90 17.74±1.74 叶绿素b' 13.06±1.10 13.93±0.65 132-羟基叶绿素b 3.07±0.56 6.19±0.18 14.05±2.50 151-羟基内酯叶绿素b 2.76±0.23 4.75±0.80 5.29±0.62 脱镁叶绿素b 8.71±1.28 56.45±2.87 脱镁叶绿素b' 1.44±0.22 2.66±0.37 132-羟基脱镁叶绿素b 0.51±0.15 3.81±0.48 叶绿素b系列 100.00 89.34±1.64 37.08±3.56 脱镁叶绿素b系列 10.66±1.64 62.92±3.56 脱镁叶绿素a组 脱镁叶绿素a 74.03±3.22 70.65±0.08 90.38±0.18 脱镁叶绿素a' 12.34±1.34 12.06±0.05 7.17±0.15 脱镁叶绿素a组 132-羟基脱镁叶绿素a 12.18±0.89 17.29±0.08 2.45±0.06 151-羟基内酯脱镁叶绿素a 1.46±0.03 脱镁叶绿素a系列 100.00 100.00 100.00 脱镁叶绿素b组 脱镁叶绿素b 82.04±2.25 73.83±0.38 87.20±0.20 脱镁叶绿素b' 5.42±1.10 11.17±0.05 3.09±0.09 132-羟基脱镁叶绿素b 1.95±0.50 15.01±0.39 3.08±0.05 151-羟基内酯脱镁叶绿素b 10.59±1.02 6.63±0.22 脱镁叶绿素b系列 100.00 100.00 100.00 脱植基叶绿素a组 脱植基叶绿素a 73.30±2.05 132-羟基脱植基叶绿素a 26.70±1.15 脱镁叶绿酸a 52.95±1.18 69.98±3.95 132-羟基脱镁叶绿酸a 30.02±3.95 151-羟基内酯脱镁叶绿酸a 47.05±1.18 脱植基叶绿素a系列 100.00 脱镁叶绿酸a系列 100.00 100.00 脱植基叶绿素b组 脱植基叶绿素b 76.06±2.87 17.39±1.36 132-羟基脱植基叶绿素b 23.94±1.32 脱镁叶绿酸b 54.36±1.53 82.61±2.26 脱镁叶绿酸b' 4.58±0.20 132-羟基脱镁叶绿酸b 12.89±0.88 151-羟基内酯脱镁叶绿酸b 28.17±0.97 脱植基叶绿素b系列 100.00 17.39±1.36 脱镁叶绿酸b系列 100.00 82.61±2.26 脱镁叶绿酸a组 脱镁叶绿酸a 75.21±1.50 76.57±2.27 56.96±0.10 脱镁叶绿酸a' 5.71±0.60 132-羟基脱镁叶绿酸a 19.09±0.45 12.35±0.45 43.04±0.06 151-羟基内酯脱镁叶绿酸a 11.08±0.16 脱镁叶绿酸a系列 100.00 100.00 100.00 脱镁叶绿酸b组 脱镁叶绿酸b 64.13±1.20 63.45±1.71 100.00 脱镁叶绿酸b' 4.67±0.31 1.97±0.17 132-羟基脱镁叶绿酸b 31.20±0.62 12.38±1.71 151-羟基内酯脱镁叶绿酸b 22.20±0.91 脱镁叶绿酸b系列 100.00 100.00 100.00 注:“'”表示相应叶绿素的同分异构体。 2.3 消化液和胶束液的体外消化特性
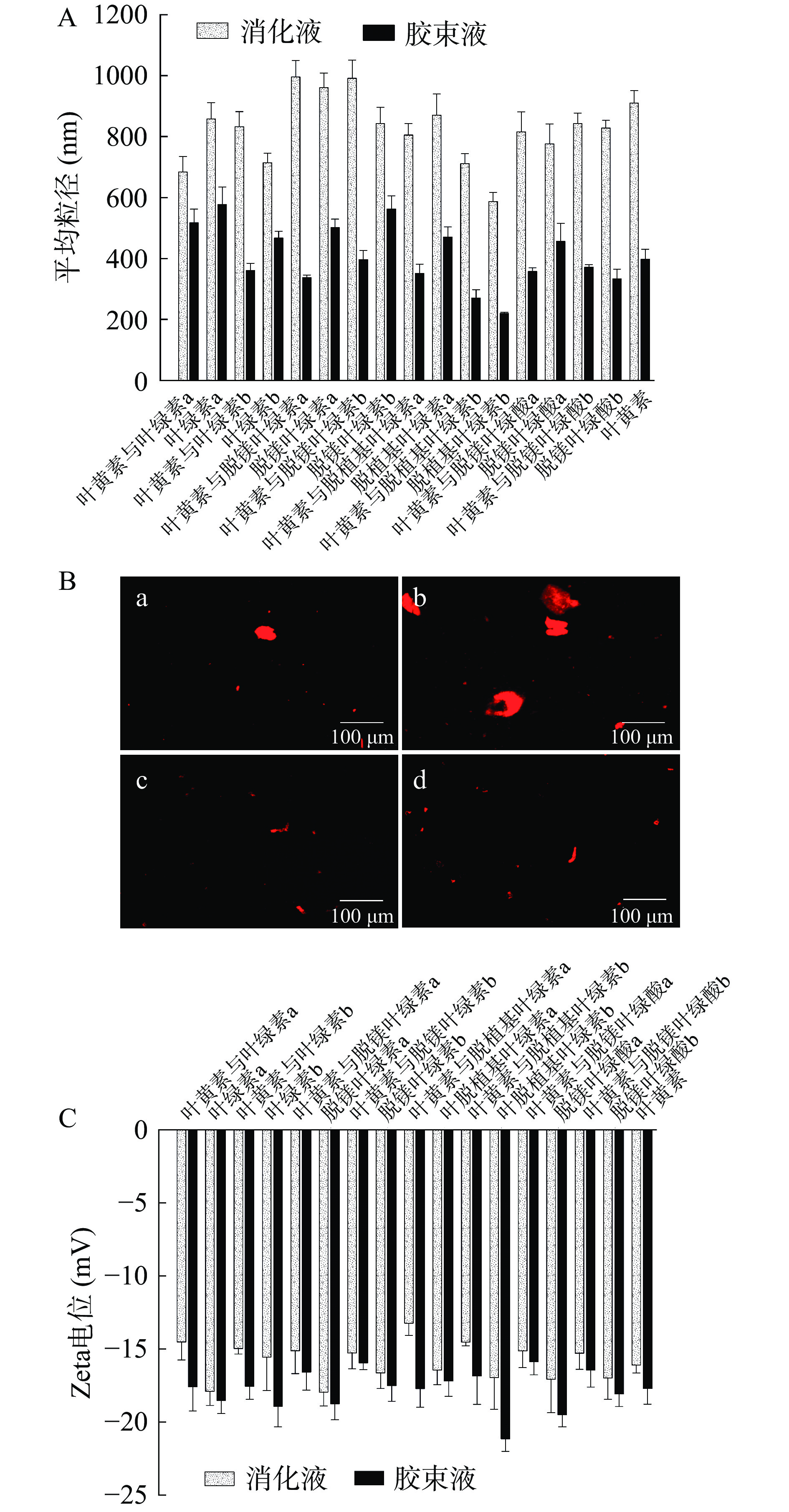
消化液和胶束液中颗粒的平均粒径、荧光图像及Zeta电位见图5。如图5(A)所示,消化液中颗粒的平均粒径为587.17~995.82 nm,胶束液颗粒的平均粒径为220.17~577.68 nm,可见消化液颗粒的平均粒径更大,说明消化液经过胶束化处理变成胶束液的过程会使溶液中的颗粒变小,可能是因为胶束化过程需经过高速离心处理,造成颗粒分散,而颗粒大小是决定微纳米颗粒乳液稳定性的一个重要因素[33],因此颗粒小的胶束液具有更好的稳定性和乳化能力[34]。同时,如图5(C)所示,消化液和胶束液的Zeta电位均为负值,且胶束液的Zeta电位绝对值普遍是大于消化液的,而Zeta电位的绝对值能代表悬浮液的稳定性,绝对值越大,稳定性就越好[35],因此说明胶束液的稳定性比消化液的更好。此外,图5(A)中脱镁叶绿酸a的平均粒径比脱镁叶绿素a小,这是因为脱镁叶绿酸a缺乏植醇导致分子尺寸减小[17]。对于消化液,共消化后的平均粒径总体上大于单独消化时的平均粒径,说明大部分结构的叶绿素在消化过程中能与叶黄素形成复合物,而部分叶绿素在与叶黄素共消化时的平均粒径减小,可能是因为受到食物基质或其他消化成分的影响;对于胶束液,总体上呈现叶绿素单独消化时的平均粒径更大,可能是因为相比于单独消化,叶绿素在与叶黄素共消化时自身更多地被分解,胶束化过程通过高速离心将两者分离,从而得到体积更小的色素颗粒。叶绿素具有荧光特性,会在一定程度上受到同处在一起的高极性叶黄素的调节[11]。以脱镁叶绿酸b为代表,图5(B)为其单独消化及与叶黄素共消化后在消化液和胶束液中分布的荧光图像,能直观反映叶绿素颗粒在消化液及胶束液中的分布和数量。在荧光模式下,可以看到消化液颗粒(a、b图)的形状和数量均比胶束液颗粒(c、d图)的更大更多,并且叶绿素与叶黄素共消化后的消化液和胶束液颗粒(b、d图)均明显大于叶绿素单独消化时的(a、c图),这与A图中脱镁叶绿酸b平均粒径的变化相一致。
![]() 图 5 消化液和胶束液中颗粒的平均粒径(A)、荧光图像(B)及Zeta电位(C)注:图B中,a:脱镁叶绿酸b单独消化后的消化液颗粒;b:脱镁叶绿酸b与叶黄素共消化后的消化液颗粒;c:脱镁叶绿酸b单独消化后的胶束液颗粒;d:脱镁叶绿酸b与叶黄素共消化后的胶束液颗粒。Figure 5. Average particle size (A), fluorescence images (B) and Zeta potential (C) of the particles in the digestiveand micellar solutions
图 5 消化液和胶束液中颗粒的平均粒径(A)、荧光图像(B)及Zeta电位(C)注:图B中,a:脱镁叶绿酸b单独消化后的消化液颗粒;b:脱镁叶绿酸b与叶黄素共消化后的消化液颗粒;c:脱镁叶绿酸b单独消化后的胶束液颗粒;d:脱镁叶绿酸b与叶黄素共消化后的胶束液颗粒。Figure 5. Average particle size (A), fluorescence images (B) and Zeta potential (C) of the particles in the digestiveand micellar solutions3. 结论
本研究通过制备8种不同结构的叶绿素,并与叶黄素共同或单独进行体外消化及胶束化实验,探究不同结构叶绿素对叶黄素生物可给率的影响规律。结果表明,在消化过程中不同结构的叶绿素均会对叶黄素产生一定程度的保护作用。通过体外消化过程中叶黄素和叶绿素各项指标的变化发现,相比于叶黄素单独消化,在与所有结构叶绿素共消化时叶黄素的回收率和生物可给率均能显著升高(P<0.05),其中脱镁叶绿素b在与叶黄素共消化时所对应的叶黄素回收率和生物可给率均为最大(90.48%和80.44%),说明脱镁叶绿素b在消化过程中对叶黄素的保护效果最佳。在本实验中,未发现叶黄素结构的变化,而叶绿素结构发生了变化,这与自身分子的化学性质有关,还会受到食物基质、环境条件等其他因素的影响。由消化液和胶束液中颗粒的平均粒径、荧光图像以及Zeta电位值可以确定胶束液体系比消化液体系更加稳定,同时叶绿素能通过与叶黄素形成复合物来提高叶黄素的生物可给率。本研究为叶黄素的生物利用增效提供了一定的参考,同时不同结构的叶绿素在与叶黄素相互作用的同时,也会受到其他因素的影响,如膳食成分、消化条件及其它脂溶性植物化学物质(如固醇)等,这些影响机制还有待进一步研究。
-
![]()
![]()
图 3 体外消化后叶黄素和叶绿素及其衍生物的胶束化率
Figure 3. Micellization rate of lutein, chlorophylls and their derivatives after in vitro digestion
![]()
图 4 体外消化后叶黄素和叶绿素及其衍生物的生物可给率
Figure 4. Bioaccessibility index of lutein, chlorophylls and their derivatives after in vitro digestion
![]()
图 5 消化液和胶束液中颗粒的平均粒径(A)、荧光图像(B)及Zeta电位(C)
注:图B中,a:脱镁叶绿酸b单独消化后的消化液颗粒;b:脱镁叶绿酸b与叶黄素共消化后的消化液颗粒;c:脱镁叶绿酸b单独消化后的胶束液颗粒;d:脱镁叶绿酸b与叶黄素共消化后的胶束液颗粒。
Figure 5. Average particle size (A), fluorescence images (B) and Zeta potential (C) of the particles in the digestiveand micellar solutions
表 1 8种常见叶绿素及其衍生物的化学结构
Table 1 Chemical structures of eight common chlorophylls and their derivatives
名称 结构类型 R1 R2 R3 R4 R5 叶绿素a(Chlorophyll a) A CH3 H COOCH3 C20H39 Mg 132-羟基叶绿素a(132-Hydroxy chlorophyll a) A CH3 OH COOCH3 C20H39 Mg 151-羟基内酯叶绿素a(151-Hydroxy-lactone chlorophyll a) B CH3 OH (152)COOCH3 C20H39 Mg 叶绿素b(Chlorophyll b) A CHO H COOCH3 C20H39 Mg 132-羟基叶绿素b(132-Hydroxy chlorophyll b) A CHO OH COOCH3 C20H39 Mg 151-羟基内酯叶绿素b(151-Hydroxy-lactone chlorophyll b) B CHO OH (152)COOCH3 C20H39 Mg 脱镁叶绿素a(Pheophytin a) A CH3 H COOCH3 C20H39 2H 132-羟基脱镁叶绿素a(132-Hydroxy pheophytin a) A CH3 OH COOCH3 C20H39 2H 151-羟基内酯脱镁叶绿素a(151-Hydroxy-lactone pheophytin a) B CH3 OH (152)COOCH3 C20H39 2H 脱镁叶绿素b(Pheophytin b) A CHO H COOCH3 C20H39 2H 132-羟基脱镁叶绿素b(132-Hydroxy pheophytin b) A CHO OH ZZCOOCH3 C20H39 2H 脱植基叶绿素a(Chlorophyllide a) B CHO OH (152)COOCH3 C20H39 2H 132-羟基脱植基叶绿素a(132-Hydroxy chlorophyllide a) A CH3 H COOCH3 H Mg 151-羟基内酯脱植基叶绿素a(151-Hydroxy-lactone chlorophyllide a) A CH3 OH COOCH3 H Mg 脱植基叶绿素b(Chlorophyllide b) B CH3 OH (152)COOCH3 H Mg 132-羟基脱植基叶绿素b(132-Hydroxy chlorophyllide b) A CHO H COOCH3 H Mg 151-羟基内酯脱植基叶绿素b(151-Hydroxy-lactone chlorophyllide b) A CHO OH COOCH3 H Mg 脱镁叶绿酸a(Pheophorbide a) B CHO OH (152)COOCH3 H Mg 132-羟基脱镁叶绿酸a(132-Hydroxy pheophorbide a) A CH3 H COOCH3 H 2H 151-羟基内酯脱镁叶绿酸a(151-Hydroxy-lactone pheophorbide a) A CH3 OH COOCH3 H 2H 脱镁叶绿酸b(Pheophorbide b) B CH3 OH (152)COOCH3 H 2H 132-羟基脱镁叶绿酸b(132-Hydroxy pheophorbide b) A CHO H COOCH3 H 2H 151-羟基内酯脱镁叶绿酸b(151-Hydroxy-lactone pheophorbide b) A CHO OH COOCH3 H 2H 脱植基叶绿素a(Chlorophyllide a) B CHO OH (152)COOCH3 H 2H  下载: 导出CSV
下载: 导出CSV
表 2 不同色素浓度的标准曲线方程
Table 2 Standard curve equations for different pigments concentrations
色素种类 DAD检测波长
(nm)标准曲线方程 决定系数
(R2)叶黄素 445 y=0.000315x−0.001214 0.9993 叶绿素a 664 y=0.000006x−0.002447 0.9969 叶绿素b 647 y=0.000005x+0.000973 0.9979 脱镁叶绿素a 408 y=0.000005x−0.003201 0.9935 脱镁叶绿素b 436 y=0.000003x+0.010738 0.9937 脱植基叶绿素a 664 y=0.000006x−0.002447 0.9969 脱植基叶绿素b 647 y=0.000005x+0.000973 0.9979 脱镁叶绿酸a 408 y=0.000003x−0.004508 0.9989 脱镁叶绿酸b 436 y=0.000003x−0.006572 0.9992 注:标准曲线方程中y为色素的浓度(μmol/mL),x为峰面积。
下载: 导出CSV
表 3 消化前后叶绿素及其衍生物的组成含量变化(%)
Table 3 Changes in the composition contents of chlorophylls and their derivatives before and after digestion (%)
叶绿 素种类 消化前 消化后 叶绿素单独消化 叶绿素与叶黄素共消化 叶绿素a组 叶绿素a 91.02±0.82 30.49±5.21 叶绿素a' 5.81±0.51 7.18±1.22 132-羟基叶绿素a 0.76±0.10 2.23±0.26 151-羟基内酯叶绿素a 2.41±0.54 0.64±0.30 脱镁叶绿素a 48.92±5.25 75.92±0.76 脱镁叶绿素a' 8.24±0.91 9.57±0.35 132-羟基脱镁叶绿素a 2.04±0.29 14.50±0.41 151-羟基内酯脱镁叶绿素a 0.26±0.01 叶绿素a系列 100.00 40.54±6.44 脱镁叶绿素a系列 59.46±6.44 100.00 叶绿素b组 叶绿素b 81.12±1.21 64.46±1.90 17.74±1.74 叶绿素b' 13.06±1.10 13.93±0.65 132-羟基叶绿素b 3.07±0.56 6.19±0.18 14.05±2.50 151-羟基内酯叶绿素b 2.76±0.23 4.75±0.80 5.29±0.62 脱镁叶绿素b 8.71±1.28 56.45±2.87 脱镁叶绿素b' 1.44±0.22 2.66±0.37 132-羟基脱镁叶绿素b 0.51±0.15 3.81±0.48 叶绿素b系列 100.00 89.34±1.64 37.08±3.56 脱镁叶绿素b系列 10.66±1.64 62.92±3.56 脱镁叶绿素a组 脱镁叶绿素a 74.03±3.22 70.65±0.08 90.38±0.18 脱镁叶绿素a' 12.34±1.34 12.06±0.05 7.17±0.15 脱镁叶绿素a组 132-羟基脱镁叶绿素a 12.18±0.89 17.29±0.08 2.45±0.06 151-羟基内酯脱镁叶绿素a 1.46±0.03 脱镁叶绿素a系列 100.00 100.00 100.00 脱镁叶绿素b组 脱镁叶绿素b 82.04±2.25 73.83±0.38 87.20±0.20 脱镁叶绿素b' 5.42±1.10 11.17±0.05 3.09±0.09 132-羟基脱镁叶绿素b 1.95±0.50 15.01±0.39 3.08±0.05 151-羟基内酯脱镁叶绿素b 10.59±1.02 6.63±0.22 脱镁叶绿素b系列 100.00 100.00 100.00 脱植基叶绿素a组 脱植基叶绿素a 73.30±2.05 132-羟基脱植基叶绿素a 26.70±1.15 脱镁叶绿酸a 52.95±1.18 69.98±3.95 132-羟基脱镁叶绿酸a 30.02±3.95 151-羟基内酯脱镁叶绿酸a 47.05±1.18 脱植基叶绿素a系列 100.00 脱镁叶绿酸a系列 100.00 100.00 脱植基叶绿素b组 脱植基叶绿素b 76.06±2.87 17.39±1.36 132-羟基脱植基叶绿素b 23.94±1.32 脱镁叶绿酸b 54.36±1.53 82.61±2.26 脱镁叶绿酸b' 4.58±0.20 132-羟基脱镁叶绿酸b 12.89±0.88 151-羟基内酯脱镁叶绿酸b 28.17±0.97 脱植基叶绿素b系列 100.00 17.39±1.36 脱镁叶绿酸b系列 100.00 82.61±2.26 脱镁叶绿酸a组 脱镁叶绿酸a 75.21±1.50 76.57±2.27 56.96±0.10 脱镁叶绿酸a' 5.71±0.60 132-羟基脱镁叶绿酸a 19.09±0.45 12.35±0.45 43.04±0.06 151-羟基内酯脱镁叶绿酸a 11.08±0.16 脱镁叶绿酸a系列 100.00 100.00 100.00 脱镁叶绿酸b组 脱镁叶绿酸b 64.13±1.20 63.45±1.71 100.00 脱镁叶绿酸b' 4.67±0.31 1.97±0.17 132-羟基脱镁叶绿酸b 31.20±0.62 12.38±1.71 151-羟基内酯脱镁叶绿酸b 22.20±0.91 脱镁叶绿酸b系列 100.00 100.00 100.00 注:“'”表示相应叶绿素的同分异构体。
下载: 导出CSV
-
[1] 郑樱, 陈新彬, 马岩, 等. 叶黄素生物学功能与相关慢性疾病的研究进展[J]. 中国食物与营养,2023,29(3):51−55. [ZHENG Y, CHEN X B, MA Y, et al. Research progress on the biological function of lutein and related chronic diseases[J]. Food and Nutrition in China,2023,29(3):51−55.] doi: 10.3969/j.issn.1006-9577.2023.03.010 ZHENG Y, CHEN X B, MA Y, et al . Research progress on the biological function of lutein and related chronic diseases[J]. Food and Nutrition in China,2023 ,29 (3 ):51 −55 . doi: 10.3969/j.issn.1006-9577.2023.03.010[2] 吴静, 张美霞, 严密. 叶黄素及玉米黄素与老年性黄斑变性的研究进展[J]. 眼科新进展,2009,29(10):791−795. [WU J, ZHANG M X, YAN M. Research advances on lutein and zeaxanthin for age-related macular degeneration[J]. Recent Advances in Ophthalmology,2009,29(10):791−795.] WU J, ZHANG M X, YAN M . Research advances on lutein and zeaxanthin for age-related macular degeneration[J]. Recent Advances in Ophthalmology,2009 ,29 (10 ):791 −795 .[3] DAI Z, SONG J, CHEN Y, et al. Study on the bioavailability of stevioside-encapsulized lutein and its mechanism[J]. Food Chemistry,2021,354(2):129528.
[4] 郭静, 胡坦, 潘思轶. 食品运载体系包埋叶黄素的研究进展[J]. 食品科学,2022,43(1):313−320. [GUO J, HU T, PAN S Y. Progress in research on food delivery systems for lutein encapsulation[J]. Food Science,2022,43(1):313−320.] GUO J, HU T, PAN S Y . Progress in research on food delivery systems for lutein encapsulation[J]. Food Science,2022 ,43 (1 ):313 −320 .[5] 焦岩, 李大婧, 刘春泉, 等. 纳米脂质体提高叶黄素的稳定性[J]. 食品工业,2019,40(4):24−27. [JIAO Y, LI D J, LIU C Q, et al. The improvement of stability of lutein by nanoliposomes[J]. The Food Industry,2019,40(4):24−27.] JIAO Y, LI D J, LIU C Q, et al . The improvement of stability of lutein by nanoliposomes[J]. The Food Industry,2019 ,40 (4 ):24 −27 .[6] 王晓. 高稳定性高生物利用度叶黄素初乳及其微胶囊制备优化研究[D]. 聊城:聊城大学, 2022. [WANG X. Study on optimized preparation of lutein-loaded emulsionsand microcapsules with high stability and bioavailability[D]. Liaocheng:Liaocheng University, 2022.] WANG X. Study on optimized preparation of lutein-loaded emulsionsand microcapsules with high stability and bioavailability[D]. Liaocheng: Liaocheng University, 2022.
[7] 石晓晴, 黄月英, 沈一唯, 等. 叶黄素的生物学作用及制剂研究进展[J]. 中国药房,2017,28(4):561−565. [SHI X Q, HUANG Y Y, SHEN Y W, et al. Research progress on biological effects and preparations of lutein[J]. China Pharmacy,2017,28(4):561−565.] doi: 10.6039/j.issn.1001-0408.2017.04.36 SHI X Q, HUANG Y Y, SHEN Y W, et al . Research progress on biological effects and preparations of lutein[J]. China Pharmacy,2017 ,28 (4 ):561 −565 . doi: 10.6039/j.issn.1001-0408.2017.04.36[8] MINH A T P, BUCKNALL M P, ARCOT J. Co-ingestion of red cabbage with cherry tomato enhances digestive bioaccessibility of anthocyanins but decreases carotenoid bioaccessibility after simulated in vitro gastro-intestinal digestion[J]. Food Chemistry,2019,298:125040. doi: 10.1016/j.foodchem.2019.125040
[9] MOJICA L, JUAN C M D, CASTILLO G. Lutein as a functional food ingredient:Stability and bioavailability[J]. Journal of Functional Foods,2020,66:103771. doi: 10.1016/j.jff.2019.103771
[10] 肖亚茹. 多酚在不同消化阶段对叶黄素肠道吸收的影响[D]. 镇江:江苏大学, 2022. [XIAO Y R. Effects of polyphenols on intestinal absorption of lutein in different digestive stages[D]. Zhenjiang:Jiangsu University, 2022.] XIAO Y R. Effects of polyphenols on intestinal absorption of lutein in different digestive stages[D]. Zhenjiang: Jiangsu University, 2022.
[11] 曲方. 环境对类胡萝卜素聚集体吸收及荧光光谱的影响[D]. 长春:吉林大学, 2020. [QU F. Effects of environment on absorption and fluorescence spectra of carotenoid aggregates[D]. Changchun:Jilin University, 2020.] QU F. Effects of environment on absorption and fluorescence spectra of carotenoid aggregates[D]. Changchun: Jilin University, 2020.
[12] 段智红, 袁圣亮, 吕应年, 等. 可食性大型海藻的叶绿素及其衍生物研究进展[J]. 食品工业科技,2018,39(20):337−342. [DUAN Z H, YUAN S L, LÜ Y N, et al. Research progress of chlorophyll and its derivatives from edible macroalga[J]. Science and Technology of Food Industry,2018,39(20):337−342.] DUAN Z H, YUAN S L, LÜ Y N, et al . Research progress of chlorophyll and its derivatives from edible macroalga[J]. Science and Technology of Food Industry,2018 ,39 (20 ):337 −342 .[13] CHEN K, RIOS J J, PEREZ G A, et al. Comprehensive chlorophyll composition in the main edible seaweeds[J]. Food Chemistry,2017,228:625−633. doi: 10.1016/j.foodchem.2017.02.036
[14] 董书琦, 陈达, 秦巧平, 等. 高等植物叶绿素和类胡萝卜素代谢研究进展[J]. 植物生理学报,2023,59(5):793−802. [DONG S Q, CHEN D, QIN Q P, et al. Advances in metabolism of chlorophylls and carotenoids in higher plants[J]. Plant Physiology Journal,2023,59(5):793−802.] DONG S Q, CHEN D, QIN Q P, et al . Advances in metabolism of chlorophylls and carotenoids in higher plants[J]. Plant Physiology Journal,2023 ,59 (5 ):793 −802 .[15] LEE E, AHN H, CHOE E. Effects of light and lipids on chlorophyll degradation[J]. Food Science and Biotechnology,2014,23(4):1061−1065. doi: 10.1007/s10068-014-0145-x
[16] 李根, 张成, 王强, 等. 植物叶绿素代谢途径及其分子调控[J]. 四川农业科技,2021(4):41−45. [LI G, ZHANG C, WANG Q, et al. Plant chlorophyll metabolic pathway and its molecular regulation[J]. Sichuan Agricultural Science and Technology,2021(4):41−45.] doi: 10.3969/j.issn.1004-1028.2021.04.014 LI G, ZHANG C, WANG Q, et al . Plant chlorophyll metabolic pathway and its molecular regulation[J]. Sichuan Agricultural Science and Technology,2021 (4 ):41 −45 . doi: 10.3969/j.issn.1004-1028.2021.04.014[17] CHEN K, ROCA M. In vitro bioavailability of chlorophyll pigments from edible seaweeds[J]. Journal of Functional Foods,2018,41:25−33. doi: 10.1016/j.jff.2017.12.029
[18] LI Y, AGARRY I E, DING D, et al. Screening of dephytinization reaction of chlorophyll pigments with citrus acetone powder by UPLC-DAD-MS[J]. Journal of Food Science,2023,88(1):147−160. doi: 10.1111/1750-3841.16411
[19] AGARRY I E, DING D S, LI Y C, et al. In vitro bioaccessibility evaluation of chlorophyll pigments in single and binary carriers[J]. Food Chemistry,2023,415:135757. doi: 10.1016/j.foodchem.2023.135757
[20] CHEN K, ROCA M. Cooking effects on bioaccessibility of chlorophyll pigments of the main edible seaweeds[J]. Food Chemistry,2019,295:101−109. doi: 10.1016/j.foodchem.2019.05.092
[21] JIN Z, AGARRY I E, LI Y, et al. In vitro bioaccessibility evaluation of pheophytins in gelatin/polysaccharides carrier[J]. Food Chemistry,2023,408:135252. doi: 10.1016/j.foodchem.2022.135252
[22] VIERA I, HERRERA M, ROCA M. In vitro bioaccessibility protocol for chlorophylls[J]. Journal of Agricultural and Food Chemistry,2021,69(31):8777−8786. doi: 10.1021/acs.jafc.1c02815
[23] CHEN K, ROCA M. In vitro digestion of chlorophyll pigments from edible seaweeds[J]. Journal of Functional Foods,2018,40:400−407. doi: 10.1016/j.jff.2017.11.030
[24] BOEHM V, LIETZ G, OLMEDILLA A B, et al. From carotenoid intake to carotenoid blood and tissue concentrations-implications for dietary intake recommendations[J]. Nutrition Reviews,2021,79(5):544−573. doi: 10.1093/nutrit/nuaa008
[25] VIERA I, CHEN K, RIOS J J, et al. First-pass metabolism of chlorophylls in mice[J]. Molecular Nutrition & Food Research,2018,62(17):1800562.
[26] YANG X, TANG J, MUSTARD J F, et al. Solar-induced chlorophyll fluorescence that correlates with canopy photosynthesis on diurnal and seasonal scales in a temperate deciduous forest[J]. Geophysical Research Letters,2015,42(8):2977−2987. doi: 10.1002/2015GL063201
[27] EGEA I, BIAN W, BAESAN C, et al. Chloroplast to chromoplast transition in tomato fruit:Spectral confocal microscopy analyses of carotenoids and chlorophylls in isolated plastids and time-lapse recording on intact live tissue[J]. Annals of Botany,2011,108(2):291−297. doi: 10.1093/aob/mcr140
[28] 李新甜, 徐亚元, 张钟元, 等. 基质结构对纳米结构脂质载体中叶黄素生物利用度的调控机制[J]. 食品科学,2022,43(12):130−138. [LI X T, XU Y Y, ZHANG Z Y, et al. Regulatory effect of matrix structures on bioaccessibility of lutein loaded into nanostructured lipid carriers[J]. Food Science,2022,43(12):130−138.] doi: 10.7506/spkx1002-6630-20210421-300 LI X T, XU Y Y, ZHANG Z Y, et al . Regulatory effect of matrix structures on bioaccessibility of lutein loaded into nanostructured lipid carriers[J]. Food Science,2022 ,43 (12 ):130 −138 . doi: 10.7506/spkx1002-6630-20210421-300[29] 李梦杰. 膳食结构对柑橘类胡萝卜素生物利用度和抗氧化活性的影响研究[D]. 武汉:华中农业大学, 2022. [LI M J. Effects of dietary structure on bioavailability and antioxidant activity of citrus carotenoids[D]. Wuhan:Huazhong Agricultural University, 2022.] LI M J. Effects of dietary structure on bioavailability and antioxidant activity of citrus carotenoids[D]. Wuhan: Huazhong Agricultural University, 2022.
[30] 葛俊珂, 孙丽娜, 赵涵, 等. 叶绿素的提取方法及药理作用进展[J]. 科技创新与应用,2016(24):14. [GE J K, SUN L N, ZHAO H, et al. Advances in extraction methods and pharmacological effects of chlorophyll[J]. Technology Innovation and Application,2016(24):14.] GE J K, SUN L N, ZHAO H, et al . Advances in extraction methods and pharmacological effects of chlorophyll[J]. Technology Innovation and Application,2016 (24 ):14 .[31] MARONEZE M M, ZEPKA L Q, LOPES E J, et al. Chlorophyll oxidative metabolism during the phototrophic and heterotrophic growth of scenedesmus obliquus[J]. Antioxidants,2019,8(12):1−15.
[32] NINOMIYA R, MATSUOKA K, MOROI Y. Micelle formation of sodium chenodeoxycholate and solubilization into the micelles:Comparison with other unconjugated bile salts[J]. Biochimica Et Biophysica Acta-Molecular and Cell Biology of Lipids,2003,1634(3):116−125. doi: 10.1016/j.bbalip.2003.09.003
[33] 陈书博. 酶解桃胶多糖的结构表征、乳化性与乳液体外消化特性研究[D]. 杭州:浙江工商大学, 2022. [CHEN S B. Study on structure characterization emulsification and in vitro digestibility of enzymatic hydrolyzed prach gum polysaccharide[D]. Hangzhou:Zhejiang Gongshang University, 2022.] CHEN S B. Study on structure characterization emulsification and in vitro digestibility of enzymatic hydrolyzed prach gum polysaccharide[D]. Hangzhou: Zhejiang Gongshang University, 2022.
[34] LAZZARO F, SAINT J A, VIOLLEAU F, et al. Gradual disaggregation of the casein micelle improves its emulsifying capacity and decreases the stability of dairy emulsions[J]. Food Hydrocolloids,2017,63:189−200. doi: 10.1016/j.foodhyd.2016.08.037
[35] YUE M, HUANG M, ZHU Z, et al. Effect of ultrasound assisted emulsification in the production of pickering emulsion formulated with chitosan self-assembled particles:Stability, macro, and micro rheological properties[J]. LWT-Food Science and Technology,2022,154:112595. doi: 10.1016/j.lwt.2021.112595
-
期刊类型引用(7)
1. 刘晓蕊,王静,吴静,李茹一. 多糖对红葡萄酒颜色稳定性的影响研究进展. 食品与发酵工业. 2025(03): 413-420 .  百度学术
百度学术
2. 陈鑫龙,张媛媛,徐令斌,白雪冰,李爱华,陶永胜. 用多酚类指标表征干红葡萄酒味感质量的模型构建. 农业工程学报. 2024(08): 281-289 . 百度学术
3. 聂旭元,张军强,况刚,王强. 澄清处理对干红葡萄原酒中矿质元素的影响. 食品安全质量检测学报. 2024(15): 175-181 . 百度学术
4. 赵现方,浦宁,杨雪芹,李发珠,缪博,王丽,南立军,王振吉. 基于CIELab色空间法分析光质对葡萄酒颜色的影响. 中国果菜. 2024(09): 19-22 . 百度学术
5. 朱袁正鸿,石方舟,张众,杨慧,王征,孙文静,张军翔. 微氧处理下赤霞珠葡萄酒香气的外源多酚基质效应. 食品与发酵工业. 2024(17): 111-119 . 百度学术
6. 黄越,黄传书,杨碧文,赵珮,王梅,周鸿媛,刘艳,吴均,戴宏杰. 不同品种桑椹发酵酒的体外抗氧化、降血糖及降血脂活性研究. 食品与发酵工业. 2024(22): 150-158 . 百度学术
7. 董书甲,任辉,刘露,赵铖,丁之恩. 葡萄酒酿造中常用辅料及应用研究. 安徽农业科学. 2024(22): 1-7+19 . 百度学术
其他类型引用(2)
 下载:
下载:





计量
- 文章访问数: 108
- HTML全文浏览量: 8
- PDF下载量: 14
- 被引次数: 9



