


Isolation, Identification and Enzyme Activity Analysis of Dominant Fermentation Strains from Traditional Pickled and Dried Mustard in Ningbo
-
摘要: 以宁波地区梅干菜半成品和成品为试验材料,采用十倍稀释法分离其中优势发酵菌株,从半成品中得到7株细菌,分别命名为MGCB1~MGCB7,从成品中得到3株细菌,分别命名为MGC1~MGC3。通过形态学观察、生理生化试验及16S rRNA序列分析对菌种进行鉴定,结果表明,MGCB1~MGCB7分别为枯草芽孢杆菌属(Bacillus subtilis)、空气芽孢杆菌属(Bacillus aerius)、高山芽孢杆菌属(Bacillus altitudinis)、短小芽孢杆菌属(Bacillus pumilus)、琥珀葡萄球菌属(Staphylococcus succinus)、琥珀葡萄球菌属 (Staphylococcus succinus)和解淀粉芽孢杆菌属(Bacillus amyloliquefaciens),MGC1~MGC3分别为芽孢杆菌属(Bacillus sp.)、枝芽孢杆菌属(Virgibacillus halodenitrificans)、哥特氏芽孢杆菌属(Cytobacillus gottheilii)。MGCB1和MGCB7产酶能力相对较高,在37 ℃培养96 h后,蛋白酶活分别为16.39±0.79和13.45±0.46 U/mL,纤维素酶活分别为3.27±0.13和1.60±0.02 U/mL。研究结果可为梅干菜纯种发酵剂的开发及工业化生产提供有益参考。Abstract: With the semi-finished and finished products of pickled and dried mustard in Ningbo as experimental materials, the dominant bacteria strains were isolated by ten-fold dilution method. The results showed that seven and three bacteria were obtained from the semi-finished and finished products, respectively, which were named as strain MGCB1~MGCB7 and strain MGC1~MGC3. The ten strains were identified by morphological observation, physiological and biochemical tests, and 16S rRNA sequence analysis. They were likely belong to Bacillus subtilis (MGCB1), Bacillus aerius (MGCB2), Bacillus altitudinis (MGCB3), Bacillus pumilus (MGCB4), Staphylococcus succinus (MGCB5), Staphylococcus succinus (MGCB6) and Bacillus amyloliquefaciens (MGCB7), Bacillus sp.
(MGC1), Virgibacillus halodenitrificans (MGC2), and Cytobacillus gottheilii (MGC3), respectively. The protease activities of MGCB1 and MGCB7 were 16.39±0.79 and 13.45±0.46 U/mL, and the cellulase activities of MGCB1 and MGCB7 were 3.27±0.13 and 1.60±0.02 U/mL, respectively, after cultured at 37 ℃ for 96 h. These findings provide a useful reference for developing appropriate starter cultures to improve the quality of traditional pickled and dried mustard. -
梅干菜亦称梅菜,是以雪里蕻、芥菜或油菜为主要原料,经挑选、去根、清洗、除水、腌制和干制等传统工艺制作而成的传统特色发酵蔬菜,在长江三角洲地区尤为常见,至今已有数百年的加工历史[1-2]。梅干菜色泽油光黄黑、滋味鲜美、香气浓郁、风味独特,富含多种维生素、膳食纤维及锌、镁、钾等7种人体所必需的矿物质元素[3-4]。同时梅干菜还具有消食健胃、降血脂和降血压等保健功能,获得了美国食品药品监督管理局的认可,被定为“天然健康食品”[5]。
由于传统梅干菜常采用自然发酵方式,多依赖自然状态下的优势菌株来实现产品发酵,因而其中的微生物种类繁多,菌落结构演替复杂。关于梅干菜的研究,目前大多集中在营养成分[6]、干燥技术[7]、挥发性风味物质分析[8-9]、提取物抗菌抗氧化活性[10-11]及加工工艺[12]等方面,但对梅干菜中菌种分离鉴定及酶活研究相对较少。尚雪娇等[13]采用传统培养方法从梅干菜分离出5种共12株乳酸菌,经分子生物学鉴定分别为植物乳杆菌(Lactobacillus plantarum)、短乳杆菌(Lactobacillus brevis),副干酪乳杆菌副干酪亚种(Lactobacillus paracasei subsp. paracasei)、食品乳杆菌(Lactobacillus alimentarius)和融合魏斯氏菌(Weissella confuse)。此外,蛋白酶能分解蛋白质生成多种氨基酸,而氨基酸是梅干菜的主要风味来源之一[14],纤维素酶可将纤维素降解为小分子糖类,在多种生理生化反应下形成梅干菜特有的香气成分[15-16]。因此,开展梅干菜微生物分离鉴定及酶活相关研究,对获得梅干菜发酵过程中相关微生物作用及其与特征风味的关系、提升其品质和附加值具有现实意义[17]。
本研究从宁波地区的梅干菜半成品和成品中分离纯化出10株细菌,通过菌落、菌体形态特征观察和生理生化试验分析初步判断其种属,之后根据16S rRNA序列同源性及进化树分析进行分类鉴定,最后通过对蛋白酶活和纤维素酶活的分析,筛选出适合梅干菜发酵的菌株,为梅干菜纯种发酵剂的研发及工业化生产提供理论基础。
1. 材料与方法
1.1 材料与仪器
梅干菜半成品:添加雪里蕻重量6%的食盐,常温下腌制2个月,经清洗、挑选、切段后,烘干至水分含量15%~20%,之后采用100 ℃蒸汽处理5 min,2021年5月下旬取样;梅干菜成品:梅干菜半成品装坛密封,常温发酵2个月,2021年7月下旬取样 由宁波市鄞州三丰可味食品有限公司提供;氢氧化钠、氯化钠、过氧化氢、无水磷酸氢二钠、磷酸二氢钠、琼脂粉 上海沪试实验室器材股份有限公司;三氯乙酸、乙二胺四乙酸二钠、3, 5-二硝基水杨酸、对二甲氨基苯甲醛、二苯胺、N, N-二甲基对苯二胺二盐酸盐、乙酸钙、羟甲基纤维素钠 上海阿拉丁生化科技股份有限公司;酪蛋白 北京索莱宝科技有限公司;其余试剂均为分析纯 上海源叶生物科技有限公司;胰蛋白胨大豆琼脂培养基(TSA)、胰蛋白胨大豆肉汤培养基(TSB)、MRS肉汤培养基、酵母浸粉 北京陆桥技术股份有限公司;TSINGKE TSP701-50 Trelief Bacteria Genomic DNA Kit试剂盒、TE缓冲液、PCR扩增引物、Taq PCR Master Mix 南京擎科生物科技有限公司。
DA2204B电子天平 上海天美天平仪器有限公司;HH-S4数显恒温水浴锅 江苏金怡仪器科技有限公司;PTC-200TM PCR仪 美国Bio-Rad公司;SW-CJ-2D超净工作台 苏州净化设备有限公司;YXQ-100S11蒸汽灭菌锅 上海博讯实业有限公司医疗设备厂;XS-3000-200三目生物显微镜 北京京宁国大科技发展有限责任公司;DHG-9245A电热恒温鼓风干燥箱 上海一恒科学仪器有限公司;BCD-218STPS冰箱 青岛海尔股份有限公司;TGLL-18G型高速冷冻离心机 江苏省太仓市仪器厂;pHS-3C型精密pH计 上海三信仪表厂;GL-88B型漩涡混合器 海门市其林贝尔仪器制造有限公司;HX-4型拍打式无菌均质器 上海沪析实业有限公司;UV-1100型紫外可见分光光度计 上海美谱达仪器有限公司;JY-ECP 3000电泳仪 北京君意东方电泳设备有限公司。
1.2 实验方法
1.2.1 菌株分离纯化
无菌操作下称取5 g梅干菜样品放入规格20×22 cm的无菌均质袋,加入45 mL 0.9%无菌生理盐水,于均质器中以8.0/s的速度正反各拍打2 min,制成1:10的样品匀液[18],按照10倍稀释法稀释至107倍,充分混匀后取各个稀释梯度溶液0.1 mL分别加入TSA固体培养基和MRS固体培养基中,用涂布棒将稀释液均匀涂布于培养基表面,放入37 ℃恒温培养箱于50%湿度条件下培养18~24 h。挑取生长旺盛的不同形态特征的单菌落,经多次划线分离,直至菌落的颜色、大小、形态一致,对其进行编号并保存备用。
1.2.2 菌株形态学观察
观察平板中菌落的生长情况,包括大小、外观、颜色、边缘、黏着度等菌落形态特征。在生物显微镜油镜下观察经革兰氏染色后的菌株,并记录菌体形态特征。
1.2.3 生理生化鉴定
通过氧化酶、过氧化氢酶、糖发酵、甲基红、V-P、苯丙氨基酸脱氨酶、吲哚、硝酸盐还原、纤维素分解、3-酮基乳糖、脲酶、色氨酸脱氨酶、明胶液化、精氨酸双水解酶、乙酸氧化和七叶灵水解试验,以及腌制蔬菜常用浓度确定的不同NaCl含量(4%、8%、12%、16%)、不同蔗糖含量(2%、10%、20%、40%)和不同pH(3.5、5.5、7.5、9.5、11.5)条件下菌株生长情况的一系列试验对菌株进行生理生化特性分析。具体培养基的配制和鉴定参照《常见细菌系统鉴定手册》[19]和《伯杰氏细菌鉴定手册》[20]。
1.2.4 菌株分子生物学鉴定
1.2.4.1 基因组DNA提取
参照TSINGKE TSP701-50 Trelief Bacteria Genomic DNA Kit试剂盒中的方法提取供试菌株基因组DNA。
1.2.4.2 16S rRNA的PCR扩增
以微生物基因组DNA为模板进行PCR扩增。PCR扩增体系:取200 µL PCR管,加入PCR Master mix 12.5 µL,上下游引物各1 µL(细菌通用引物为27F和1492R),DNA模板2.5 µL,补ddH2O至25 µL,标记,盖紧,将PCR管放入PCR仪中进行扩增。PCR反应条件:95 ℃预变性5 min;35个循环×(94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸50 s);72 ℃延伸10 min。
1.2.4.3 测序与分析
PCR扩增结束后,进行琼脂糖凝胶电泳检测,观察PCR产物有无非特异性条带,以及产物的纯度和DNA片段大小,无误后,将PCR产物送至南京擎科生物科技有限公司进行测序。将测得的序列在NCBI上的GenBank序列数据库进行Blast比对分析,在基因数据库中进行同源性搜索比对,找到同源性较高的细菌后,挑出模式菌株,并下载这些菌株的基因序列,利用MEGA 6.0软件进行系统发育树的构建,进而确定此菌株的种属信息。
1.2.5 菌株的酶活测定
1.2.5.1 粗酶液的制备
从平板上挑取单个菌落转接至25 mL TSB液体培养基,在37 ℃条件下培养96 h。吸取10 mL培养好的菌液至EP管,4 ℃、10000 r/min离心10 min,上清液即为待测酶液。
1.2.5.2 蛋白酶活测定
吸取1.0 mL酶液于40±1 ℃水浴锅中保温2 min,样品管加入40±1 ℃预热5 min的酪蛋白溶液(10.0 g/L),在40±1 ℃下反应10 min,立即加入2.0 mL 65.4 g/L的三氯乙酸溶液终止反应(对照管先在酶液中加三氯乙酸溶液反应后再加酪蛋白溶液)。静置10 min后过滤,取1.0 mL滤液依次加5.0 mL 42.4 g/L的Na2CO3溶液和1.0 mL福林试剂,在40±1 ℃下显色20 min,于680 nm下测吸光值A680。采用福林法[21]测定生成的酪氨酸含量,标准曲线测得为y=0.0101x+0.0042,R2=0.9995,以每分钟水解酪蛋白产生1 µg酪氨酸所需的酶量定义为一个酶活力单位,结果表示为U/mL。
1.2.5.3 纤维素酶活测定
参照QB 2583-2003《纤维素酶制剂》中所规定的CMCA-DNS法[22]进行。吸取2.0 mL pH6.0的CMC-Na溶液于25 mL具塞刻度试管中,加入0.5 mL酶液(对照管不加),置于50±1 ℃水浴锅中反应30 min,迅速加入3.0 mL DNS试剂,接着向对照管中加0.5 mL酶液。将试验管于沸水浴加热10 min,冷却,用去离子水定容至25 mL,于540 nm下测吸光值A540。以葡萄糖作标准曲线计算还原糖含量,标准曲线测得为y=0.3288x−0.0661,R2=0.9978,以1 h水解羧甲基纤维素钠底物生成1 mg还原糖为1个酶活力单位,结果以U/mL表示。
1.3 数据处理
试验每个处理均重复三次,结果表示为平均值±标准差,采用MEGA 6.0软件绘制系统发育树,并用统计软件SPSS22.0进行显著性(LSD;P<0.05)分析,用Excel 2016软件作图。
2. 结果与分析
2.1 形态学观察
2.1.1 菌落形态特征观察
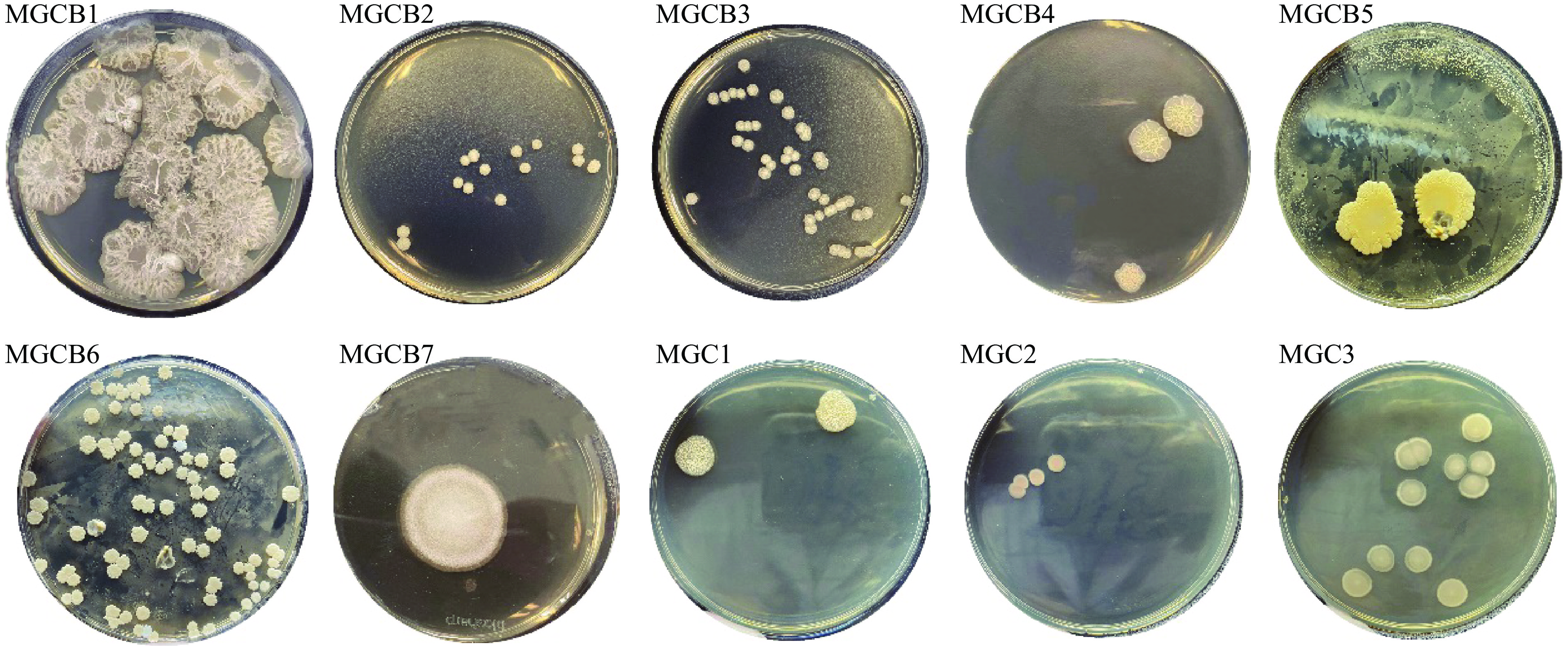
通过对梅干菜样品进行稀释、涂布和分离纯化,结果显示TSA平板上有菌落生长,其中从梅干菜半成品中分离出菌落形态不同的细菌共7株,分别命名为MGCB1、MGCB2、MGCB3、MGCB4、MGCB5、MGCB6、MGCB7,从梅干菜成品中分离出细菌3株,分别命名为MGC1、MGC2、MGC3,经分离纯化后得到的菌落形态特征如图1和表1所示,筛选出的菌株大都为白色或淡黄色、凸起的圆形菌落,表面呈褶皱状、细丝状或颗粒状,少数的菌株表面较光滑,菌株直径总体相差较大,多数有不规则的边缘,通过挑取时的状态判断MGCB1和MGCB7的黏着性相对较强。
表 1 菌落形态特征Table 1. Characteristics of colony morphological菌株 直径(mm) 外观 颜色 边缘 黏着性 MGCB1 20.0±1.6 圆形,表面有褶皱 灰白色 凹凸不平 很强 MGCB2 3.0±0.1 圆形,表面呈丝状凸起 乳白色 整齐 弱 MGCB3 3.1±0.1 圆形,表面呈丝状 乳白色 较整齐 弱 MGCB4 9.4±3.7 圆形,中心呈丝状 淡黄色 呈花边状 弱 MGCB5 17.6±0.5 不规则圆形,表面光滑湿润 黄色 不规则凸起 弱 MGCB6 3.4±0.1 圆形,表面光滑湿润 奶油色 呈锯齿状 弱 MGCB7 25.3±4.1 圆形,表面颗粒状 白色 整齐 较强 MGC1 10.0±0.8 圆形,表面呈细丝状 乳白色 较整齐 弱 MGC2 4.0±0.2 圆形,表面光滑 淡粉色 规则有晕边 弱 MGC3 7.6±1.2 圆形,表面光滑 淡黄色 规则有晕边 弱 2.1.2 菌体形态特征观察
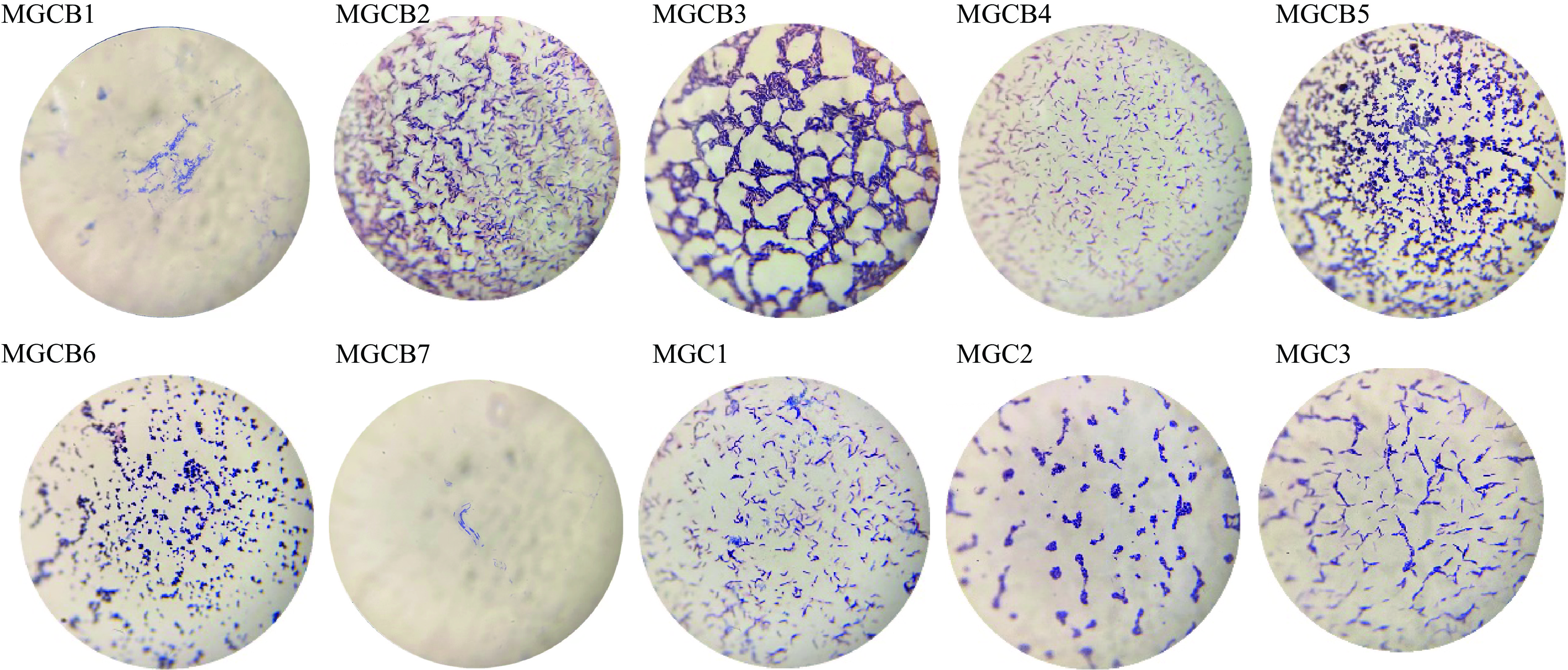
菌体形态特征是通过观察待测菌株在显微镜下的状态和革兰氏染色结果来判断,如图2所示,在光学显微镜的油镜下(总共放大倍数为100倍),分离出的大部分菌株在显微观察下都为杆状菌,只有菌株MGCB5和MGCB6为球状菌,革兰氏染色结果显示10株菌均为阳性(G+)。形态学仅仅是通过一些宏观和微观的观察,初步了解菌落菌体的形态特征,但不能确定其确切的种属,因此需要进一步利用生理生化特性分析及分子生物学方法进行鉴定。
2.2 生理生化鉴定
将分离得到的10株菌进行一系列生理生化试验,由表2可知,10株菌均不能合成吲哚,不能分解纤维素,不能发酵乳糖,不能生成苯丙氨基酸脱氨酶和色氨酸脱氨酶,但均能发酵葡萄糖和水解七叶灵;除MGC3外都能产生精氨酸双水解酶,菌株中只有MGCB5和MGCB6能够氧化乙酸;除MGCB1和MGCB7外,接触酶试验都为阳性;另外MGCB1、MGCB2、MGCB7、MGC2和MGC3能把硝酸盐还原成亚硝酸盐,为硝酸盐还原试验阳性,其余菌株都为阴性;明胶液化试验中MGC2和MGC3没有菌生长,另外MGCB3和MGCB4为明胶液化阴性,说明其不能产生明胶酶进而分解明胶,其余都为阳性。根据生理生化特性分析结果参考《常见细菌系统鉴定手册》[19]和《伯杰氏细菌鉴定手册》[20],初步判断MGCB1、MGCB2、MGCB3、MGCB4、MGCB7、MGC1、MGC2、MGC3都为芽孢杆菌属(Bacillus),MGCB5和MGCB6为葡萄球菌属(Staphylococcus)。
表 2 生理生化试验结果Table 2. Results of physiological and biochemical tests特征 菌株 MGCB1 MGCB2 MGCB3 MGCB4 MGCB5 MGCB6 MGCB7 MGC1 MGC2 MGC3 氧化酶 + + + + − − + + − − 接触酶 − + + + + + − + + + 糖发酵 + + + + + + + + + + 甲基红试验 − − + + − − + + − − V-P试验 + + + + − − + + − − 苯丙氨基酸脱氨酶 − − − − − − − − − − 吲哚试验 − − − − − − − − − − 硝酸盐还原 + + − − − − + − + + 纤维素分解 − − − − − − − − − − 3-酮基乳糖 − − − − − − − − − − 脲酶 + − − − + + + − − − 色氨酸脱氨酶 − − − − − − − − − − 明胶液化 + + − − + + + + − − 精氨酸双水解酶 + + + + + + + + + − 乙酸氧化 − − − − + + − − − − 七叶灵水解 + + + + + + + + + + 耐
渗
透
压氯化钠 4% + + + + + + + + + + 8% + + + + + + + + + + 12% − + − + + + + − − − 16% − − − − − − + − − − 蔗糖 2% + + + + + + + + + − 10% + + + + + + + + − − 20% + + + + + + + + − − 40% + − − − − − − − − − 耐
酸
耐
碱pH 3.5 − − − − − − − − − − 5.5 + + + + + + + + − − 7.5 + + + + + + + + − − 9.5 + + + + + + + + + + 11.5 + + + + + + + + − − 微生物分类 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 葡萄球菌属 葡萄球菌属 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 注:“+”表示反应呈阳性或生长;“−”表示反应呈阴性或不生长。 在耐盐实验中,菌株在4% NaCl和8% NaCl环境下长势基本较好,其中MGCB7菌株耐盐能力最强,但是MGCB1、MGCB3、MGC1、MGC2和MGC3在含有12%和16% NaCl的培养基中均不能正常生长。在耐糖实验中,半成品中的7种菌在2%、10%、20%蔗糖浓度下都能正常生长,其中MGCB1在40%蔗糖浓度下也能生长,耐糖性能最好;MGC2只在2%蔗糖浓度下生长,耐糖能力较差;菌株中MGC3的耐糖性能最差。在耐pH实验中,随着生长环境pH的升高,菌种生长状况总体上先变好再变差,半成品中的7种菌和MGC1在pH7.5时长势最好,在pH11.5时仍能生长则证明其耐碱能力较强,MGC2和MGC3只在pH9.5时生长,证明其耐酸能力较弱。
2.3 分子生物学鉴定
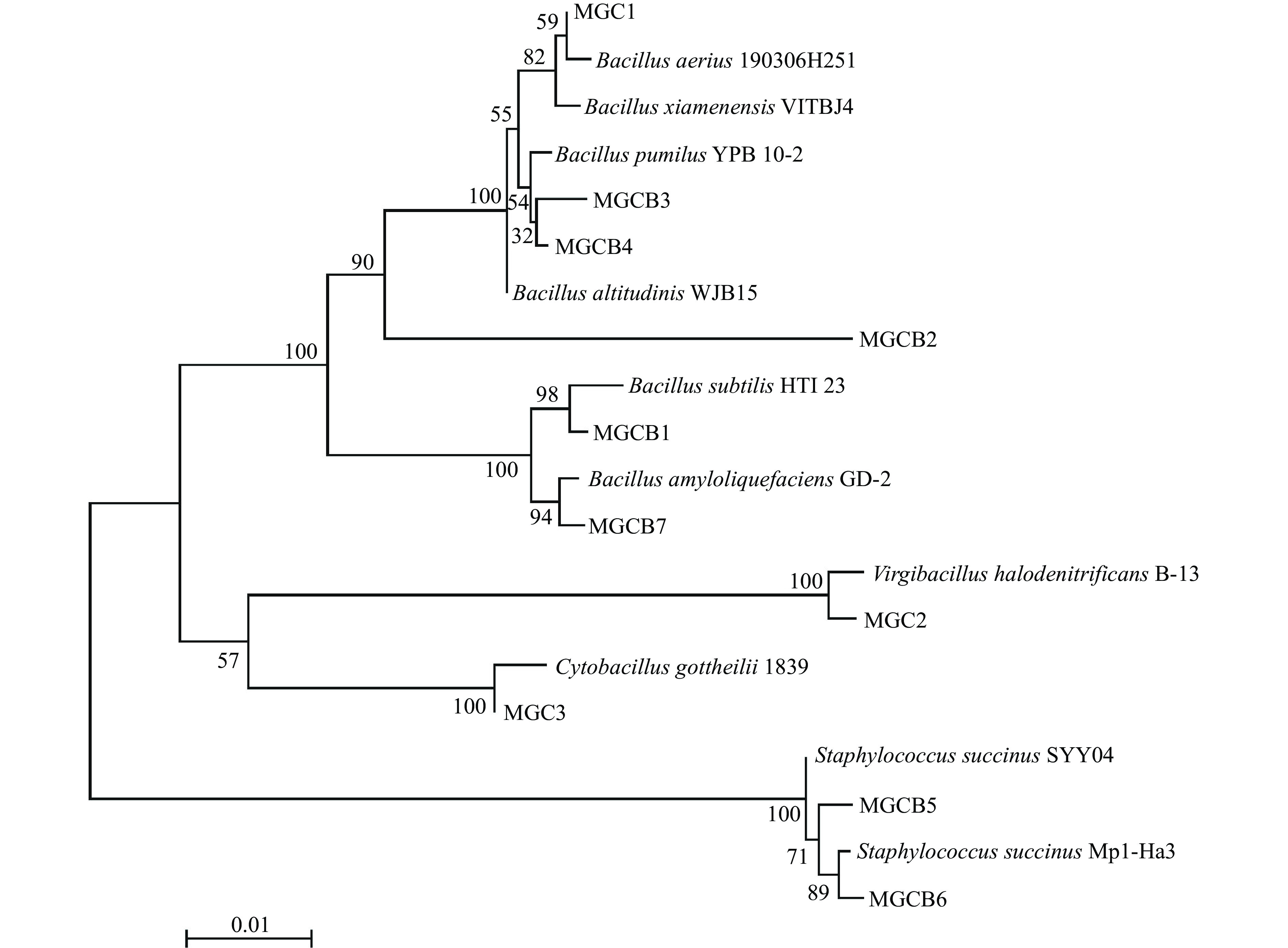
将待测菌株进行16S rRNA测序后,将序列结果登录NCBI数据库进行相似性比对,同时利用MEGA 6.0软件进行系统发育树的绘制,结果见表3和图3。发育树各分支的长短代表了各进化关系的远近,即距离越小亲缘关系越近[23]。由表3可知,10株菌株除MGCB2之外与NCBI数据库中的基因序列同源性均达到99%以上,和标准菌株序列覆盖率均达到98%以上,E值都为0.0。结合形态学观察和生理生化特性分析,MGCB1~MGCB7可能分别为枯草芽孢杆菌属(Bacillus subtilis)、空气芽孢杆菌属IP10菌株(Bacillus aerius)、高山芽孢杆菌属(Bacillus altitudinis)、短小芽孢杆菌属(Bacillus pumilus)、琥珀葡萄球菌属(Staphylococcus succinus)、琥珀葡萄球菌属(Staphylococcus succinus)和解淀粉芽孢杆菌属(Bacillus amyloliquefaciens);MGC1、MGC2和MGC3可能分别为芽孢杆菌属(Bacillus sp.
)、枝芽孢杆菌属(Virgibacillus halodenitrificans)和哥特氏芽孢杆菌属(Cytobacillus gottheilii)。 表 3 菌株的16S rRNA序列同源性比对结果Table 3. Homologous alignment analysis of 16S rRNA sequences of strains菌株
编号BLAST结果 序列大小(bp) 覆盖百分率(%) 同源性(%) 菌株登录号 MGCB1 枯草芽孢杆菌属(Bacillus subtilis) 1449 99 100.00 MK521066.1 MGCB2 空气芽孢杆菌属(Bacillus aerius) 797 86 98.70 MT225712.1 MGCB3 高山芽孢杆菌属(Bacillus altitudinis) 1450 99 99.93 KU877629.1 MGCB4 短小芽孢杆菌属(Bacillus pumilus) 1449 99 99.86 JQ308586.1 MGCB5 琥珀葡萄球菌属(Staphylococcus succinus) 1450 98 100.00 MZ674190.1 MGCB6 琥珀葡萄球菌属(Staphylococcus succinus) 1452 99 99.59 MN461567.1 MGCB7 解淀粉芽孢杆菌属(Bacillus amyloliquefaciens) 1451 99 99.86 MH373537.1 MGC1 芽孢杆菌属(Bacillus sp. ) 1433 100 99.93 MH475934.1 MGC2 枝芽孢杆菌属(Virgibacillus halodenitrificans) 1468 99 99.45 MN756670.1 MGC3 哥特氏芽孢杆菌属(Cytobacillus gottheilii) 1390 99 99.86 CP071709.1 注:以上菌株E值均为0。 ![]() 图 3 待测菌株基于16S rRNA的系统发育树Figure 3. The phylogenetic tress of the tested strains based on 16S rRNA
图 3 待测菌株基于16S rRNA的系统发育树Figure 3. The phylogenetic tress of the tested strains based on 16S rRNA2.4 酶活分析
蛋白酶能将大分子蛋白水解成小分子多肽和氨基酸,纤维素酶能将纤维素降解为多糖,这些都会对梅干菜风味产生作用,同时纤维素的降解对梅干菜的硬度、咀嚼、质构有一定影响[24],故产蛋白酶和纤维素酶的能力可以作为评价梅干菜发酵菌株发酵性能优劣的重要指标[15]。
由表4可知,在分离得到的10株菌中,MGCB1和MGCB7的产蛋白酶能力相对较强,分别为16.39±0.79和13.45±0.46 U/mL;MGC2没有产蛋白酶的能力;37 ℃培养96 h后蛋白酶活在7~10 U/mL之间的有MGCB2、MGCB3、MGCB4和MGC1;琥珀葡萄球菌属的两株菌(MGCB5、MGCB6)和MGC3产蛋白酶的能力很弱。
表 4 不同菌株的产酶能力Table 4. Enzyme production capacity of different bacterial strains菌株 蛋白酶活(U/mL) 纤维素酶活(U/mL) MGCB1枯草芽孢杆菌属 16.39±0.79a 3.27±0.13a MGCB2空气芽孢杆菌属 8.40±0.13cd 2.00±0.08b MGCB3高山芽孢杆菌属 9.20±0.71c 1.32±0.01de MGCB4短小芽孢杆菌属 8.76±0.42c 1.31±0.02e MGCB5琥珀葡萄球菌属 0.87±0.12e ND MGCB6琥珀葡萄球菌属 0.75±0.09e ND MGCB7解淀粉芽孢杆菌属 13.45±0.46b 1.60±0.02c MGC1芽孢杆菌属 7.82±0.36d 1.42±0.01 de MGC2枝芽孢杆菌属 ND 1.19±0.06f MGC3哥特氏芽孢杆菌属 0.29±0.05e 1.44±0.06d 注:ND表示未检测到酶活;同列不同小写字母表示差异显著(P<0.05)。 10株菌株中产纤维素酶能力最强的是MGCB1,为3.27±0.13 U/mL;琥珀葡萄球菌属的两株菌(MGCB5、MGCB6)没有产纤维素酶的能力;其余菌株在37 ℃培养96 h后的纤维素酶活均在1~2 U/mL之间,产纤维素酶能力相对较弱(表4)。
3. 讨论
传统梅干菜发酵过程中特殊风味的形成主要源于微生物的代谢活动及协同发酵作用,因此传统梅干菜中存在多种共生微生物[25]。尚雪娇等[13]研究表明,湖北恩施地区梅干菜中存在的菌属有乳酸杆菌属(Lactobacillus)、假单胞菌属(Pseudomonas)、葡萄球菌属(Staphylococcus)、魏斯氏菌属(Weissella)、鞘脂单胞菌属(Sphingomonas)、嗜冷杆菌属(Psychrobacter)等,其相对丰度分别为60.50%、8.69%、2.86%、1.73%、1.28%、1.07%。刘长根[15]对来自不同地区的25份梅干菜样品进行高通量测序分析,发现丰度较高的菌属主要有乳酸杆菌属(Lactobacillus)、葡萄球菌属(Staphylococcus)、硫胺素芽孢杆菌属(Aneurinibacillus)、芽孢杆菌属(Bacillus)、拉乌尔菌属(Raoultella)等。虞任莹等[26]采用Illumina Miseq高通量测序技术对宁波地区梅干菜成品进行分析,发现慢生芽孢杆菌属(Lentibacillus)、交替芽胞杆菌属(Alteribacillus)、嗜盐球菌属(Halococcus)、乳杆菌属(Lactobacillus)、海单胞菌属(Oceanimonas)、枝芽孢菌属(Virgibacillus)和丛毛单胞菌属(Comamonas)等为梅干菜后发酵阶段的优势菌属,相对丰度均在6%以上。
本研究从宁波地区梅干菜半成品和成品中分离出的10株菌中,经鉴定也发现有芽孢杆菌属(Bacillus sp.)、枝芽孢杆菌属(Virgibacillus halodenitrificans)和葡萄球菌属(Staphylococcus succinus)等,推测这些菌可能对宁波地区梅干菜风味和品质的形成有重要贡献。邓丽[27]从自然发酵芥菜中分离出9株对金黄色葡萄球菌有拮抗作用的菌,分别为1株中村芽孢杆菌(Bacillus nakamurai)、1株短小芽孢杆菌(Bacillus pumilus)、2株贝莱斯芽孢杆菌(Bacillus velezensis)、2株莫海威芽孢杆菌(Bacillus mojavensis)和3株解淀粉芽孢杆菌(Bacillus amyloliquefaciens),说明芽孢杆菌对食源性致病菌会产生拮抗作用,故芽孢杆菌属(Bacillus sp.)可能对梅干菜的品质存在有益影响。程艳薇等[28]从镇远道菜中筛选出1株贝莱斯芽孢杆菌(Bacillus velezensis)、2株芽孢杆菌(Bacillus sp.)、3株解淀粉芽孢杆菌(Bacillus amyloliquefaciens)和1株变形杆菌(Proteus sp.),发现芽孢杆菌(Bacillus sp.)产生物胺含量较高,因此芽孢杆菌属(Bacillus sp.)可能与梅干菜风味的形成有关。葡萄球菌属不仅能代谢过氧化氢酶防止产生异味,还可能代谢产生氨基酸、醇类、酮类和醛类等风味组分[14]。Xu等[29]研究发现,葡萄球菌属是发酵红辣椒中的优势细菌属,发酵结束时(32 d)占95%,另外证实了从中分离的曲霉、芽孢杆菌、Brachybacterium、Microbacterium和葡萄球菌与正丁醇、乙酸盐和苯酚等关键气味活性物质的形成高度相关,其中曲霉和芽孢杆菌与这些风味物质呈正相关。
此外有研究表明,不同菌种之间的协同作用可通过抑制一些酶和微生物减少具有不良滋味的含硫化合物的形成,在发酵中后期还能促进酯类、醇类、酮类、醛类和杂环化合物等风味组分的形成[30]。本研究也从宁波地区梅干菜中分离获得了8株芽孢杆菌和2株葡萄球菌,推测梅干菜特殊风味的形成过程依赖于多菌种共生发酵模式,是多种菌在发酵过程中相互协同作用的结果,其中MGCB1、MGCB6和MGCB7具有较好的生理耐受性能及产酶能力,可能对梅干菜风味形成产生重要影响。
4. 结论
从宁波地区的梅干菜半成品中分离出7种菌,从梅干菜成品中分离出3种菌,经鉴定MGCB1~MGCB7可能分别为枯草芽孢杆菌属、空气芽孢杆菌属、高山芽孢杆菌属、短小芽孢杆菌属、琥珀葡萄球菌属、琥珀葡萄球菌属和解淀粉芽孢杆菌属,MGC1~MGC3可能分别为芽孢杆菌属、枝芽孢杆菌属和哥特氏芽孢杆菌属。10菌株在8% NaCl培养基中均能生长,菌株MGCB2、MGCB4、MGCB5和MGCB6能在12% NaCl培养基中生长,MGCB7能在16% NaCl培养基中生长,表明其耐盐能力较强。MGCB1(枯草芽孢杆菌属)和MGCB7(解淀粉芽孢杆菌属)的产酶能力相对较高,37 ℃培养96 h后蛋白酶活分别为16.39±0.79和13.45±0.46 U/mL,纤维素酶活分别为3.27±0.13和1.60±0.02 U/mL。MGCB1、MGCB6和MGCB7在生理耐受性能及产酶能力方面具有较好的综合性能,其协同作用可能会对梅干菜的品质和风味形成有重要贡献,但具体生产性能还有待进一步研究。
-
![]()
图 3 待测菌株基于16S rRNA的系统发育树
Figure 3. The phylogenetic tress of the tested strains based on 16S rRNA
表 1 菌落形态特征
Table 1 Characteristics of colony morphological
菌株 直径(mm) 外观 颜色 边缘 黏着性 MGCB1 20.0±1.6 圆形,表面有褶皱 灰白色 凹凸不平 很强 MGCB2 3.0±0.1 圆形,表面呈丝状凸起 乳白色 整齐 弱 MGCB3 3.1±0.1 圆形,表面呈丝状 乳白色 较整齐 弱 MGCB4 9.4±3.7 圆形,中心呈丝状 淡黄色 呈花边状 弱 MGCB5 17.6±0.5 不规则圆形,表面光滑湿润 黄色 不规则凸起 弱 MGCB6 3.4±0.1 圆形,表面光滑湿润 奶油色 呈锯齿状 弱 MGCB7 25.3±4.1 圆形,表面颗粒状 白色 整齐 较强 MGC1 10.0±0.8 圆形,表面呈细丝状 乳白色 较整齐 弱 MGC2 4.0±0.2 圆形,表面光滑 淡粉色 规则有晕边 弱 MGC3 7.6±1.2 圆形,表面光滑 淡黄色 规则有晕边 弱  下载: 导出CSV
下载: 导出CSV
表 2 生理生化试验结果
Table 2 Results of physiological and biochemical tests
特征 菌株 MGCB1 MGCB2 MGCB3 MGCB4 MGCB5 MGCB6 MGCB7 MGC1 MGC2 MGC3 氧化酶 + + + + − − + + − − 接触酶 − + + + + + − + + + 糖发酵 + + + + + + + + + + 甲基红试验 − − + + − − + + − − V-P试验 + + + + − − + + − − 苯丙氨基酸脱氨酶 − − − − − − − − − − 吲哚试验 − − − − − − − − − − 硝酸盐还原 + + − − − − + − + + 纤维素分解 − − − − − − − − − − 3-酮基乳糖 − − − − − − − − − − 脲酶 + − − − + + + − − − 色氨酸脱氨酶 − − − − − − − − − − 明胶液化 + + − − + + + + − − 精氨酸双水解酶 + + + + + + + + + − 乙酸氧化 − − − − + + − − − − 七叶灵水解 + + + + + + + + + + 耐
渗
透
压氯化钠 4% + + + + + + + + + + 8% + + + + + + + + + + 12% − + − + + + + − − − 16% − − − − − − + − − − 蔗糖 2% + + + + + + + + + − 10% + + + + + + + + − − 20% + + + + + + + + − − 40% + − − − − − − − − − 耐
酸
耐
碱pH 3.5 − − − − − − − − − − 5.5 + + + + + + + + − − 7.5 + + + + + + + + − − 9.5 + + + + + + + + + + 11.5 + + + + + + + + − − 微生物分类 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 葡萄球菌属 葡萄球菌属 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 芽孢杆菌属 注:“+”表示反应呈阳性或生长;“−”表示反应呈阴性或不生长。
下载: 导出CSV
表 3 菌株的16S rRNA序列同源性比对结果
Table 3 Homologous alignment analysis of 16S rRNA sequences of strains
菌株
编号BLAST结果 序列大小(bp) 覆盖百分率(%) 同源性(%) 菌株登录号 MGCB1 枯草芽孢杆菌属(Bacillus subtilis) 1449 99 100.00 MK521066.1 MGCB2 空气芽孢杆菌属(Bacillus aerius) 797 86 98.70 MT225712.1 MGCB3 高山芽孢杆菌属(Bacillus altitudinis) 1450 99 99.93 KU877629.1 MGCB4 短小芽孢杆菌属(Bacillus pumilus) 1449 99 99.86 JQ308586.1 MGCB5 琥珀葡萄球菌属(Staphylococcus succinus) 1450 98 100.00 MZ674190.1 MGCB6 琥珀葡萄球菌属(Staphylococcus succinus) 1452 99 99.59 MN461567.1 MGCB7 解淀粉芽孢杆菌属(Bacillus amyloliquefaciens) 1451 99 99.86 MH373537.1 MGC1 芽孢杆菌属(Bacillus sp. ) 1433 100 99.93 MH475934.1 MGC2 枝芽孢杆菌属(Virgibacillus halodenitrificans) 1468 99 99.45 MN756670.1 MGC3 哥特氏芽孢杆菌属(Cytobacillus gottheilii) 1390 99 99.86 CP071709.1 注:以上菌株E值均为0。
下载: 导出CSV
表 4 不同菌株的产酶能力
Table 4 Enzyme production capacity of different bacterial strains
菌株 蛋白酶活(U/mL) 纤维素酶活(U/mL) MGCB1枯草芽孢杆菌属 16.39±0.79a 3.27±0.13a MGCB2空气芽孢杆菌属 8.40±0.13cd 2.00±0.08b MGCB3高山芽孢杆菌属 9.20±0.71c 1.32±0.01de MGCB4短小芽孢杆菌属 8.76±0.42c 1.31±0.02e MGCB5琥珀葡萄球菌属 0.87±0.12e ND MGCB6琥珀葡萄球菌属 0.75±0.09e ND MGCB7解淀粉芽孢杆菌属 13.45±0.46b 1.60±0.02c MGC1芽孢杆菌属 7.82±0.36d 1.42±0.01 de MGC2枝芽孢杆菌属 ND 1.19±0.06f MGC3哥特氏芽孢杆菌属 0.29±0.05e 1.44±0.06d 注:ND表示未检测到酶活;同列不同小写字母表示差异显著(P<0.05)。
下载: 导出CSV
-
[1] HUANG S, HUANG M, FENG B. Antioxidant activity of extracts produced from pickled and dried mustard (Brassica juncea Coss. Var. foliosa Bailey)[J]. International Journal of Food Properties,2012,15(2):374−384. doi: 10.1080/10942912.2010.487628
[2] LI C, TANG Z F, HUANG M, et al. Antioxidant efficacy of extracts produced from pickled and dried mustard in rapeseed and peanut oils[J]. Journal of Food Science,2012,77(4):394−400. doi: 10.1111/j.1750-3841.2011.02606.x
[3] LI M, HUANG M, ZHU Q, et al. Pickled and dried mustard foreign matter detection using multispectral imaging system based on single shot method[J]. Journal of Food Engineering,2020,285:110106. doi: 10.1016/j.jfoodeng.2020.110106
[4] 乔倩, 李远志, 庞宇辰, 等. 梅菜干燥技术的研究[C]. 广东省食品学会. “健康食品与功能性食品配料”学术研讨会暨2016年广东省食品学会年会论文集. 中国广东广州: 广东省食品学会, 2016: 133−135. QIAO Q, LI Y Z, PANG Y C, et al. Research on drying technology of pickled and dried mustard[C]. Guangdong Food Association. Academic Symposium on “Healthy Food and Functional Food Ingredients” and Proceedings of the Annual Conference of Guangdong Food Association in 2016. China Guangzhou, Guangdong: Guangdong Food Association, 2016: 133−135.
[5] 张书弦, 李远志, 黄苇, 等. 惠州梅菜的营养价值与加工研究进展[J]. 农产品加工(学刊),2013(16):74−76. [ZHANG S X, LI Y Z, HUANG W, et al. Research progress on nutritional value and processing of pickled and dried mustard in Huizhou[J]. Agricultural Products Processing (Journal),2013(16):74−76. [6] 茹巧美, 任国平, 张学兵. 15种不同产地梅干菜营养功能与有害成分分析[J]. 中国调味品,2020,45(9):150−155. [RU Q M, REN G P, ZHANG X B. Analysis of nutritional functions and harmful components of 15 kinds of pickled and dried mustard from different origins[J]. China Condiments,2020,45(9):150−155. [7] 罗伟, 余以刚, 胡双芳, 等. 不同干燥方式加工的梅干菜风味物质研究[J]. 广东农业科学,2016,43(8):131−137. [LUO W, YU Y G, HU S F, et al. Study on the flavor compounds of pickled and dried mustard processed by different drying methods[J]. Guangdong Agricultural Science,2016,43(8):131−137. doi: 10.16768/j.issn.1004-874X.2016.08.022 [8] SHEN Q, CHENG H, PU Y F, et al. Characterization of volatile compounds in pickled and dried mustard (Brassica juncea, Coss.) using optimal HS-SPME-GC-MS[J]. CyTA-Journal of Food,2018,16(1):331−339. doi: 10.1080/19476337.2017.1380705
[9] SHEN Q, JIANG J, WANG M, et al. Volatile compounds and antioxidant properties of pickled and dried mustard as influenced by different cooking methods[J]. Journal of Food Processing and Preservation,2019,43(4):23−26.
[10] 沈清, 楼乐燕, 尹培, 等. 5种梅干菜的酚类化合物及抗氧化能力比较分析[J]. 食品科学,2018,39(12):212−218. [SHEN Q, LOU L Y, YIN P, et al. Comparative analysis of phenolic compounds and antioxidant capacity of five kinds of pickled and dried mustard[J]. Food Science,2018,39(12):212−218. doi: 10.7506/spkx1002-6630-201812033 [11] 张新, 张有做. 绍兴梅干菜提取物抗氧化活性研究[J]. 食品与发酵科技,2017,53(6):75−80. [ZHANG X, ZHANG Y Z. Antioxidant activity of extracts from pickled and dried mustard in Shaoxing[J]. Food and Fermentation Science and Technology,2017,53(6):75−80. [12] 周美琪, 周其德, 田赛莺, 等. 低盐腌制对缙云梅干菜加工品质的影响[J]. 核农学报,2018,32(8):1562−1571. [ZHOU M Q, ZHOU Q D, TIAN S Y, et al. Effects of low-salt pickling on the processing quality of pickled and dried mustard in Jinyun[J]. Journal of Nuclear Agriculture,2018,32(8):1562−1571. doi: 10.11869/j.issn.100-8551.2018.08.1562 [13] 尚雪娇, 王玉荣, 杨江, 等. 应用Illumina MiSeq高通量测序技术解析梅干菜中细菌多样性[J]. 中国酿造,2019,38(1):140−144. [SHANG X J, WANG Y R, YANG J, et al. Analysis of bacterial diversity in pickled and dried mustard using Illumina MiSeq high-throughput sequencing technology[J]. China Brewing,2019,38(1):140−144. doi: 10.11882/j.issn.0254-5071.2019.01.028 [14] 陈倩, 李永杰, 扈莹莹, 等. 传统发酵食品中微生物多样性与风味形成之间关系及机制的研究进展[J]. 食品工业科技,2021,42(9):412−419. [CHEN Q, LI Y J, HU Y Y, et al. Research progress on the relationship and mechanism between microbial diversity and flavor formation in traditional fermented food[J]. Science and Technology of Food Industry,2021,42(9):412−419. doi: 10.13386/j.issn1002-0306.2020070365 [15] 刘长根. 我国传统发酵蔬菜微生物多样性比较[D]. 南昌: 南昌大学, 2019. LIU C G. Comparison of microbial diversity of traditional fermented vegetables in my country[D]. Nanchang: Nanchang University, 2019.
[16] 张俊杰, 郭晨, 尚益民, 等. 鹰嘴豆纳豆优良发酵菌株的筛选与初步鉴定[J]. 中国酿造,2018,37(7):88−92. [ZHANG J J, GUO C, SHANG Y M, et al. Screening and preliminary identification of excellent fermentation strains of chickpea and natto[J]. China Brewing,2018,37(7):88−92. doi: 10.11882/j.issn.0254-5071.2018.07.018 [17] PENG Q, JIANG S, CHEN J, et al. Unique microbial diversity and metabolic pathway features of fermented vegetables from Hainan, China[J]. Frontiers in Microbiology,2018,9:399. doi: 10.3389/fmicb.2018.00399
[18] 中华人民共和国卫生部. GB 4789.2-2016 食品安全国家标准 食品微生物学检验 菌落总数测定[S]. 北京: 中国标准出版社, 2016. Ministry of Health of the People's Republic of China. GB 4789.2-2016 National Standard for Food Safety Microbiological examination of food Determination of total bacterial count[S]. Beijing: Standards Press of China, 2016.
[19] 东秀珠, 蔡妙英. 常见细菌系统鉴定手册[M]. 北京: 科学出版社, 2001: 364–398. DONG X Z, CAI M Y. Manual for systematic identification of common bacteria[M]. Beijing: Science Press, 2001: 364–398.
[20] BUCHANAN R E, GIBBONS N E. Bergey’s manual of determinative bacteriology[M]. 9th ed. Beijing: Science Press, 1984.
[21] 中国国家标准化管理委员会. GB/T 23527-2009 蛋白酶制剂[S]. 北京: 中国标准出版社, 2009. Standardization Administration of China. GB/T 23527-2009 Protease preparation[S]. Beijing: China Standard Press, 2009.
[22] 中华人民共和国轻工行业标准. QB 2583-2003 纤维素酶制剂[S]. 北京: 中国标准出版社, 2003. People's Republic of China Light Industry Standard. QB 2583-2003 Cellulase preparation[S]. Beijing: China Standard Press, 2003.
[23] 曹佳璐. 传统四川泡菜盐水乳酸菌多样性的研究[D]. 北京: 中国农业大学, 2017. CAO J L. Study on the diversity of lactic acid bacteria in traditional Sichuan pickle brine[D]. Beijing: China Agricultural University, 2017.
[24] 何维, 安天星, 余玲, 等. 四川太和毛霉豆豉中优势发酵菌株的分离鉴定与酶活分析[J]. 安徽农业科学,2021,49(14):157−161. [HE W, AN T X, YU L, et al. Isolation, identification and enzyme activity analysis of dominant fermentation strain from Taihe mucor tempeh in Sichuan[J]. Anhui Agricultural Sciences,2021,49(14):157−161. doi: 10.3969/j.issn.0517-6611.2021.14.042 [25] YANG X Z, HU W Z, XIU Z L, et al. Microbial dynamics and volatilome profiles during the fermentation of Chinese northeast sauerkraut by Leuconostoc mesenteroides ORC 2 and Lactobacillus plantarum HBUAS 51041 under different salt concentrations[J]. Food Research International,2020,130:108926. doi: 10.1016/j.foodres.2019.108926
[26] 虞任莹, 彭思佳, 李艺, 等. 梅干菜和腌制雪菜细菌菌群多样性分析[J]. 食品工业科技,2021,42(18):134−141. [YU R Y, PENG S J, LI Y, et al. Analysis of bacterial diversity of the pickled and dried mustard and the fermented potherb mustard[J]. Food Industry Science and Technology,2021,42(18):134−141. doi: 10.13386/j.issn1002-0306.2021010065 [27] 邓丽. 金黄色葡萄球菌拮抗细菌的分离鉴定及机制研究[D]. 贵州: 贵州大学, 2021. DENG L. Isolation, identification and mechanism of antagonistic bacteria against Staphylococcus aureus[D]. Guizhou: Guizhou University, 2021.
[28] 程艳薇, 王雪郦, 俞露, 等. 贵州镇远道菜中产生物胺细菌的初步筛选及鉴定[J]. 中国酿造,2022,41(5):106−112. [CHENG Y W, WANG X L, YU L, et al. Preliminary screening and identification of biogenic amine bacteria in Zhenyuan dish of Guizhou Province[J]. China Brewing,2022,41(5):106−112. doi: 10.11882/j.issn.0254-5071.2022.05.019 [29] XU X, WU B, ZHAO W, et al. Correlation between autochthonous microbial communities and key odorants during the fermentation of red pepper (Capsicum annuum L.)[J]. Food Microbiology,2020,91:103510. doi: 10.1016/j.fm.2020.103510
[30] XIAO Y, HUANG T, HUANG C, et al. The microbial communities and flavour compounds of Jiangxi Yancai, Sichuan Paocai and Dongbei Suancai: Three major types of traditional Chinese fermented vegetables[J]. LWT-Food Science and Technology,2020,121:108865. doi: 10.1016/j.lwt.2019.108865
-
期刊类型引用(4)
1. 梁鑫富,董庆亮. 光谱法结合分子对接技术研究柠檬苦素与牛血清白蛋白的相互作用. 食品工业科技. 2024(21): 37-44 .  本站查看
本站查看
2. 王江涛,赵晓瑜,郭彩霞. 花青素协同酶水解法对翅果油粕蛋白功能性质和结构特性的影响. 食品工业科技. 2023(20): 18-26 . 本站查看
3. 王靖蕊,彭佳敏,代成,李雁,解新安. 基于光谱和分子模拟研究半乳甘露聚糖和人血清白蛋白的相互作用. 食品工业科技. 2022(02): 42-49 . 本站查看
4. 姚红柳,付金凤,尹广婷,周雪健,苏丽红. 芦丁与弹性蛋白酶相互作用的研究. 化学试剂. 2022(01): 46-51 . 百度学术
其他类型引用(7)
 下载:
下载:



计量
- 文章访问数: 201
- HTML全文浏览量: 47
- PDF下载量: 29
- 被引次数: 11



