


Effects of Theabrownin on Brain Metabolites in D-galactose-induced Aging in Mice
-
摘要: 本研究通过D-半乳糖皮下注射建立小鼠衰老模型,利用免疫组化和UHPLC-QE-MS代谢组学技术检测正常对照小鼠、D-半乳糖模型小鼠、茶褐素(预防和治疗)干预小鼠的脑组织,探究茶褐素对D-半乳糖致衰老小鼠脑组织免疫组化的影响,筛选茶褐素对衰老小鼠作用的差异代谢物并分析其涉及的代谢通路。结果表明:茶褐素干预后,衰老小鼠脑组织中的乙酰胆碱转移酶阳性表达面密度显著升高(P<0.05);预防性给予茶褐素可以显著调节衰老小鼠脑组织中的17种代谢物,主要涉及甘油磷脂代谢、组氨酸代谢、TCA循环、乙醛酸和二羧酸代谢、β-丙氨酸代谢5条代谢通路;茶褐素治疗可以显著调节衰老小鼠脑组织中的32种代谢物,主要涉及甘油磷脂代谢,组氨酸代谢,牛磺酸和亚牛磺酸代谢,缬氨酸、亮氨酸和异亮氨酸的降解,苯丙氨酸、酪氨酸和色氨酸的生物合成5条通路。其中,肌肽、LysoPC(P-18:1(9Z))、PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z))、PC(15:0/15:0)、D-阿拉伯糖-5-磷酸盐、硫酸胆固醇和二磷酸腺苷核糖7个代谢物以及甘油磷脂代谢和组氨酸代谢2条代谢通路为茶褐素预防组和治疗组共有。茶褐素可以通过调节小鼠体内脂类代谢,能量代谢,氨基酸代谢等过程,从而起到抗衰老作用。Abstract: The objective of the study is to investigate the effect of theabrownin on the immunohistochemistry of brain tissue in aging mice, screen the differential metabolites and analyze the metabolic pathways involved. A mouse aging model was established by subcutaneous injection of D-galactose in mice. Immunohistochemical and UHPLC-QE-MS metabolomics techniques were used to detect brain samples of normal control mice, D-galactose model mice and theabrownin-treated (pretreatment and treatment) mice. The results showed that the areal density of choline acetyltransferase positive expression was significantly increased (P<0.05) after theabrownin intervention. Prophylactic administration of theabrownin could significantly regulate 17 metabolites in brain tissue of aging mice, mainly involving glycerophospholipid metabolism, histidine metabolism, TCA cycle, glyoxylate and dicarboxylate metabolism and beta-alanine metabolism. Theabrownin treatment could significantly regulate 32 metabolites in brain tissue of aging mice, mainly involving glycerophospholipid metabolism, histidine metabolism, taurine and hypotaurine metabolism, valine, leucine and isoleucine degradation, phenylalanine, tyrosine and tryptophan biosynthesis. Carnosine, LysoPC(P-18:1(9Z)), PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z)), PC(15:0/15:0), D-arabinose 5-phosphate, cholesterol sulfate, adenosine diphosphate ribose, glycerophospholipid metabolism and histidine metabolism were common in the theabrownin pretreatment and theabrownin treatment groups. Theabrownin can play an anti-aging role by regulating lipid metabolism, energy metabolism, amino acid metabolism and other processes in mice.
-
Keywords:
- theabrownin /
- immunocytochemistry /
- brain metabonomics /
- anti-aging
-
普洱茶(Camellia sinensis var. assamica)是中国云南传统的历史名茶,由大叶种晒青毛茶经微生物固态发酵而成[1-2]。其风味独特、陈香浓郁、汤色鲜红、滋味醇厚。茶褐素(Theabrownin,TB)是普洱茶的重要活性成分之一,在普洱茶中的含量可达16.86%,是普洱茶最主要的呈色物质,对普洱茶的品质与功能有决定性作用[3]。饮食调节已经被证明是干预机体衰老的有效方法之一。氧化应激被认为是导致衰老的一个重要因素,是体内自由基产生的一种负面作用[4-5]。本课题组前期研究发现茶褐素可以有效提高D-半乳糖(D-galactose,D-gal)致衰老小鼠肝脏中超氧化物歧化酶及谷胱甘肽过氧化物酶的活性,降低丙二醛含量,表明茶褐素能够对抗自由基损害,有效预防由氧化应激导致的氧化损伤,具有抗衰老作用[6]。
机体的衰老通常伴随着系统性的病变,研究这些病变过程比较复杂。近年来,代谢组学技术已被广泛应用在多种复杂疾病研究中。代谢组学是对生物体内源性物质反应生成的所有代谢产物进行研究,是机体整体功能或状态的最终体现,也是多因素作用的综合效应[7]。由于衰老的复杂性,脑组织中可能存在大量代谢产物,因此对其进行代谢组学分析并鉴定代谢物的变化,可反映衰老的发生和发展过程中的变化。本研究以D-gal皮下注射建立小鼠衰老模型,利用UHPLC-QE-MS代谢组学技术,并采用偏最小二乘判别分析、正交偏最小二乘判别分析方法结合变量投影重要度与非参数检验共同分析正常对照组、D-gal模型组、茶褐素预防组、茶褐素治疗组小鼠脑组织中代谢物的变化规律,探寻茶褐素延缓衰老的潜在代谢物,为茶褐素的作用机制研究提供新的思路。
1. 材料与方法
1.1 材料与仪器
SPF级雄性C57BL/6小鼠 5~6周龄,体质量为20±2 g,购自常州卡文斯实验动物有限公司(许可证号:SCXK(苏)2016-0010);动物饲料 江苏省协同医药生物工程有限责任公司。本实验方案开展前已获得云南农业大学实验动物管理与动物福利委员会(IACUC)的审查许可;大益牌普洱熟茶(唛号7262,批次1801,2018年7月5日生产) 勐海茶厂;D-gal >99%,美国西格玛奥德里奇公司;0.9%生理盐水 山东齐都药业有限公司;4%多聚甲醛通用型组织固定液 广州赛国生物科技有限公司;无水乙醇、二甲苯、中性树胶 分析纯,国药集团化学试剂有限公司;HE染液套装、1%牛血清白蛋白溶液(含0.3% Triton X-100)、乙酰胆碱转移酶(Choline acetyltransferase,ChAT)抗体(稀释比1:500)、HRP标记的山羊抗兔IgG抗体、抗体孵育盒 Servicebio有限公司;组化试剂盒DAB显色剂 中杉金桥公司;甲醇、乙腈、乙酸铵、氨水 分析纯,上海佰晔生物科技中心;L-2-氯苯丙氨酸 色谱纯,上海恒柏生物科技有限公司。
RE-52AA旋转蒸发仪 上海亚荣生化仪器厂;Vanquish超高效液相、Q Exactive HFX高分辨质谱、Heraeus Fresco17离心机 Thermo Fisher Scientific公司;BSA124S-CW分析天平 Sartorius公司;明澈D24 UV纯水仪 Merck Millipore公司;YM-080S超声仪 深圳市方奥微电子有限公司;JJ-12J脱水机、JB-P5包埋机 武汉俊杰电子有限公司;RM2016病理切片机 上海徕卡仪器有限公司;GFL-230烤箱 天津市莱玻瑞仪器设备有限公司;XSP-C204显微镜 北京荣兴光恒科技有限公司。
1.2 实验方法
1.2.1 普洱熟茶茶褐素的制备
普洱熟茶磨碎,过80目筛,将茶样按1:10茶水比于沸水浸提30 min,趁热过滤,重复2次,茶汤减压低温65±1 ℃浓缩,乙醇(乙醇:滤液=4:1,V/V)醇沉24 h,离心(4000 r/min,20 min)收集沉淀,蒸馏水溶解后真空冷冻干燥,得粗茶褐素(供试样,茶褐素含量:68.5%)[8]。
1.2.2 动物分组、建模与取样
将40只小鼠饲养在SPF级动物房,室温20±2 ℃,湿度45%~60%,适应性饲养2周后,随机分成4组,每组各10只,分为正常对照组(Control)、D-gal模型组(Aging)、TB预防组(TB1)和TB治疗组(TB2)。TB预防组小鼠于D-gal建模前预防性给予1.215 g/kg·bw[8]TB灌胃8周,其他组不做任何处理,随后D-gal模型组、TB预防组、TB治疗组给予皮下注射500 mg/kg·bw D-gal,正常对照组给予皮下注射等量生理盐水,连续给药16周。在D-gal建模8周后,TB治疗组同时给予1.215 g/kg·bw TB灌胃8周,D-gal模型组和正常对照组同时给予等量蒸馏水灌胃8周。试验结束后,将小鼠脊椎脱臼牺牲,冰上取出脑组织。4只小鼠的脑组织放入4%多聚甲醛组织固定液固定24 h;6只小鼠的脑组织用4 ℃生理盐水冲洗表面残血,滤纸吸干水分,液氮速冷,样品保存于−80 ℃冰箱中待测。
1.2.3 小鼠脑组织切片ChAT免疫组化
将固定液中的脑组织取出,石蜡包埋切片,用ABC法[9]进行ChAT免疫组化。切片先放入0.3%过氧化氢,用抗体孵育盒室温孵育30 min;1%牛血清白蛋白溶液(含0.3% TritonX-100)37 ℃孵育30 min;一抗ChAT 37 ℃孵育2 h后置于4 ℃冰箱过夜;二抗HRP标记的山羊抗兔IgG和链霉亲和素-生物素复合物分别在37 ℃温箱孵育2 h;0.05% DAB(含0.01%过氧化氢)室温显色2 min。以上除1%牛血清白蛋白溶液封闭后直接放入一抗外,其余各步骤之间均用0.01 mol/L磷酸盐缓冲液(pH7.4)漂洗3次,5 min/次。贴片后晾干,梯度酒精脱水,二甲苯透明,中性树脂封片。显微镜镜检,图像采集分析。用CaseViewer 2.4软件打开后,选取组织的目的区域进行400倍成像,成像完成后使用Image-Pro Plus 6.0分析软件,统一以像素面积pixel作为标准单位,分别测量每张切片中三个视野阳性的累积光密度值(Integrated optical density,IOD)以及对应的组织像素面积(Area);并计算出面密度(AREAL DENSITY)=IOD/Area。
1.2.4 UHPLC-QE-MS代谢组学分析
代谢物提取:称取50 mg脑组织样品,加入1000 μL提取液(甲醇:乙腈:水 = 2:2:1(V/V),含内标L-2-氯苯丙氨酸1 μg/mL),涡旋混匀30 s,35 Hz研磨处理4 min,超声5 min(冰水浴),重复步骤3次,−40 ℃静置1 h,将样品4 ℃,12000 r/min离心15 min,取上清于进样瓶中上机检测。所有样品另取等量上清混合成QC样品上机检测。
色谱条件:使用Vanquish超高效液相色谱仪,通过Waters ACQUITY UPLC BEH Amide (2.1 mm×100 mm,1.7 μm)液相色谱柱对目标化合物进行色谱分离。液相色谱A相为水相,含25 mmol/L乙酸铵和25 mmol/L氨水,B相为乙腈。采用梯度洗脱:0~0.5 min,95% B;0.5~7 min,95%~65% B;7~8 min,65%~40% B;8~9 min,40% B;9~9.1 min,40%~95% B;9.1~12 min,95% B。流动相流速:0.5 mL/min,柱温:25 ℃,样品盘温度:4 ℃,进样体积3 μL。
质谱条件:Thermo Q Exactive HFX质谱仪在控制软件(Xcalibur,Thermo)控制下进行一级、二级质谱数据采集。详细参数如下:鞘层气体流量:30 Arb,辅助气体流量:10 Arb,毛细管温度:350 ℃,全ms分辨率:60000,ms/ms分辨率:7500,NCE模式下碰撞能量:10/30/60,喷雾电压:4 kV(正离子模式)、3.8 kV(负离子模式)。
1.3 数据处理
原始数据经ProteoWizard软件转成mzXML格式后,使用自主编写的R程序包(内核为XCMS)进行峰识别、峰提取、峰对齐和积分等处理,再与BiotreeDB(V2.1)自建二级质谱数据库匹配进行物质注释,算法打分的Cutoff值设为0.3。对XCMS提取得到的数据运用SIMCA 14.1程序进行偏最小二乘判别分析(Partial least squares discriminant analysis,PLS-DA)和正交偏最小二乘判别分析(Orthogonal projection to latent structures,OPLS-DA)。同时结合S-plot图中的变量投影重要度(Variable importance for the projection,VIP)大于1为筛选标准,初步筛选出差异代谢物。再进一步采用单变量统计分析,得到P值。选择VIP>1且P<0.05的代谢物作为差异代谢物,将得到的差异代谢物数据运用Metabo Analyst数据库进行代谢通路富集分析。
2. 结果与分析
2.1 各组小鼠脑组织海马区ChAT免疫组化染色结果
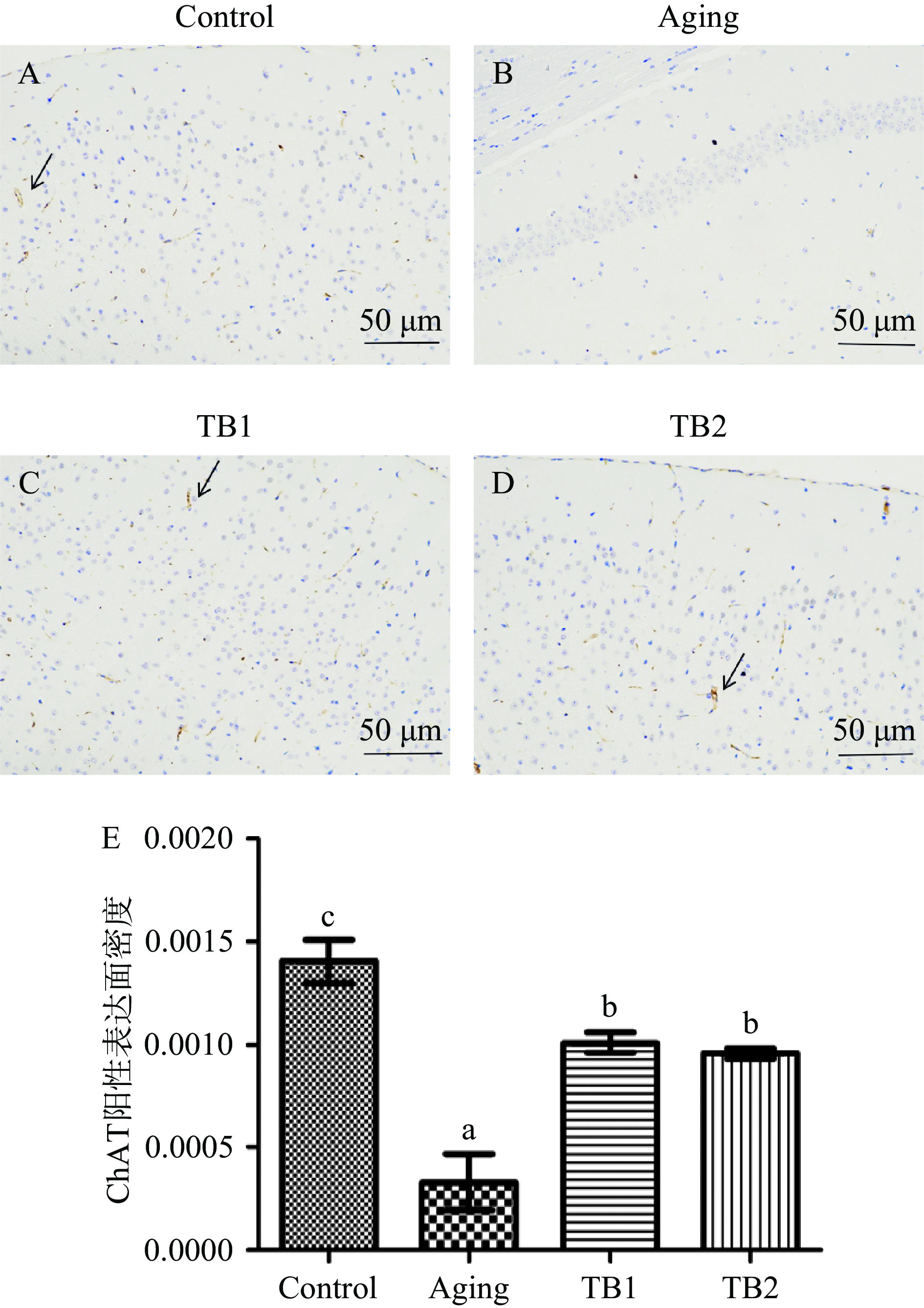
显微镜下观察小鼠脑组织海马区ChAT的阳性表达(苏木素染细胞核为蓝色;DAB显出的阳性表达为棕黄色,黑色箭头)。由图1可知,D-gal模型组ChAT阳性细胞染色较其他组减少,且面密度显著降低(P<0.05);茶褐素干预后,衰老小鼠ChAT阳性表达面密度显著升高(P<0.05),但TB1和TB2两组之间无显著性差异(P>0.05)。
![]() 图 1 各组小鼠脑组织ChAT免疫组化结果注:放大倍数:400×;不同字母表示差异显著(P<0.05)。Figure 1. ChAT immunohistochemical results of brain tissues in each group
图 1 各组小鼠脑组织ChAT免疫组化结果注:放大倍数:400×;不同字母表示差异显著(P<0.05)。Figure 1. ChAT immunohistochemical results of brain tissues in each group2.2 寻找脑组织中衰老相关的代谢物及代谢通路
通过QC样本监测UHPLC-QE-MS系统稳定性,结果如表1所示,表明本次实验建立的UHPLC-QE-MS分析方法的重复性和稳定性符合大批量脑组织样本代谢组学的分析要求。
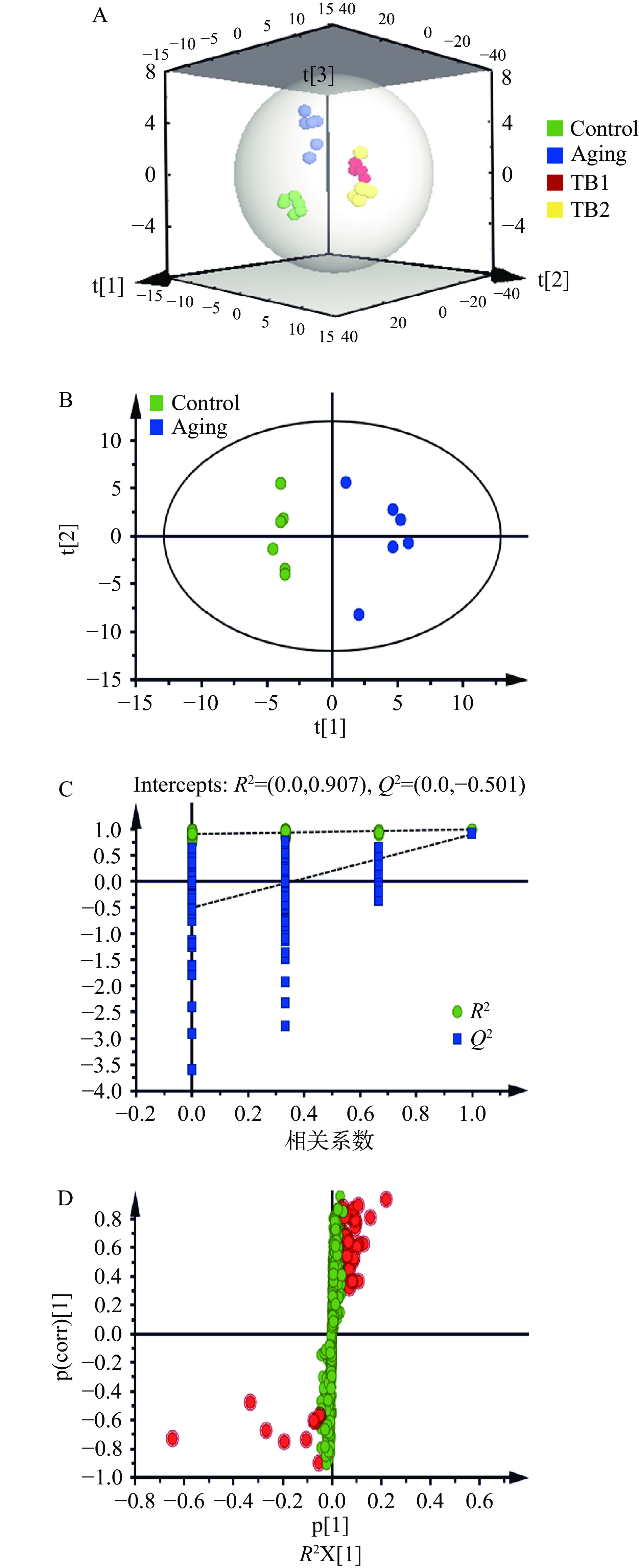
表 1 QC样品中内标响应稳定性Table 1. Stability of internal standard response in QC samples内标种类 正离子模式 负离子模式 m/z R.T. RSD m/z R.T. RSD L-2-氯苯丙氨酸 200.0473 273.324 5.8% 198.0325 264.603 4.6% 对对照组、D-gal模型组、TB预防组和TB治疗组小鼠脑组织样品进行检测,得到各组小鼠的脑组织代谢数据。运用PLS-DA对各组小鼠脑组织数据进行代谢轮廓分析(图2A),结果表明衰老使小鼠脑组织代谢轮廓发生了改变。通过对D-gal模型组与对照组小鼠脑组织数据进行OPLS-DA分析,可见D-gal模型组和对照组明显分离(图2B)并得到相应的S-plot图(图2C)以便寻找D-gal模型组与对照组的组间内源性差异代谢物。进一步对模型数据进行多次(次数n=200)随机排列实验以进行模型验证。结果表明,R2参数接近1,说明建立的模型符合样本数据的真实情况;置换检验随机模型的Q2值小于原模型的参数模型的Q2值且Q2的回归线与纵轴的截距为负值,表明模型未产生过分拟合,模型可靠(图2D)。通过对D-gal模型组和对照组在正、负离子模式下代谢物数据进行分析,找到与衰老相关的42个代谢物(表2)。衰老主要涉及乙醛酸和二羧酸代谢(Glyoxylate and dicarboxylate metabolism)、苯丙氨酸、酪氨酸和色氨酸的生物合成(Phenylalanine,tyrosine and tryptophan biosynthesis)、甘油磷脂代谢(Glycerophospholipid metabolism)、TCA循环(TCA cycle)、组氨酸代谢(Histidine metabolism)、维生素B6代谢(Vitamin B6 metabolism)、牛磺酸和亚牛磺酸代谢(Taurine and hypotaurine metabolism)、缬氨酸、亮氨酸和异亮氨酸的降解(Valine,leucine and isoleucine degradation)、嘧啶代谢(Pyrimidine metabolism)、β-丙氨酸代谢(beta-Alanine metabolism)10条代谢通路(图3A和图3B)。
![]() 图 2 不同组小鼠脑组织的比较分析注:A为不同组小鼠脑组织的PLS-DA散点图;B为对照组和D-gal模型组小鼠脑组织OPLS-DA得分图;C为置换检验图;D为S-plot图;图D中红色的点表明这些代谢物的VIP值大于等于1,绿色的点表示这些代谢物的VIP值小于1。Figure 2. Comparative analysis of mouse brain samples in different groups表 2
图 2 不同组小鼠脑组织的比较分析注:A为不同组小鼠脑组织的PLS-DA散点图;B为对照组和D-gal模型组小鼠脑组织OPLS-DA得分图;C为置换检验图;D为S-plot图;图D中红色的点表明这些代谢物的VIP值大于等于1,绿色的点表示这些代谢物的VIP值小于1。Figure 2. Comparative analysis of mouse brain samples in different groups表 2D-gal诱导衰老小鼠脑组织的差异代谢产物 Table 2. Differential metabolites in brain of aging mice induced by D-gal编号 正/负离子模式 代谢物 代谢物
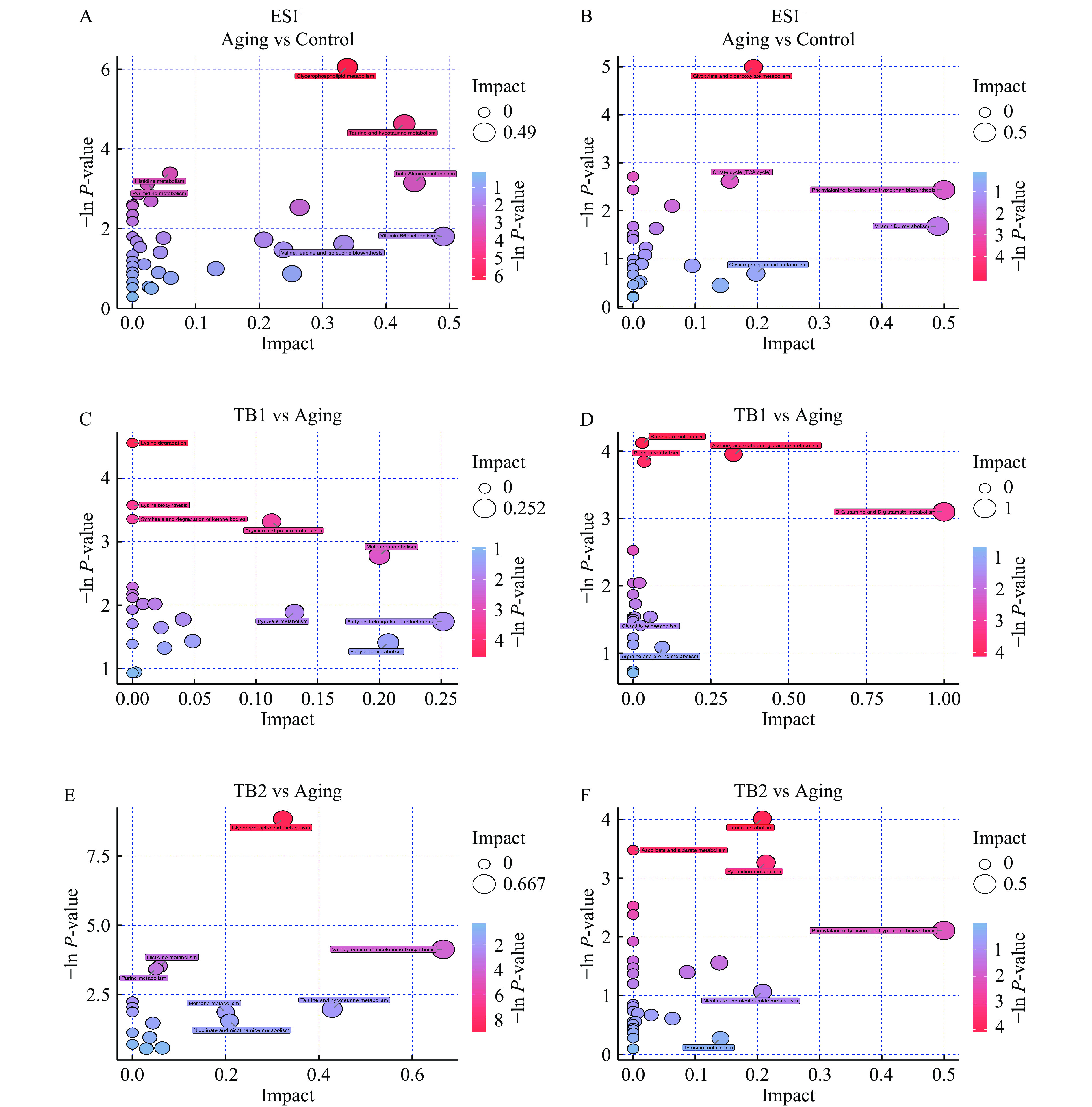
中文名称质荷比 保留时间(min) 1 ESI+ Cytosine 胞嘧啶 112.0507 478.6850 2 ESI+ Carnosine 肌肽 226.8889 439.9890 3 ESI+ 3-Methylglutarylcarnitine 3-甲基戊二酰肉碱 290.9379 442.9370 4 ESI+ Aminoadipic acid 氨基己二酸 161.1285 460.1125 5 ESI+ PE(22:5(7Z,10Z,13Z,16Z,19Z)/22:4(7Z,10Z,13Z,16Z)) PE(22:5/22:4) 842.0741 148.3530 6 ESI+ Acetyl-CoA 乙酰辅酶A 810.1326 451.3795 7 ESI+ N-Acetylhistidine N-乙酰组氨酸 197.1030 457.3510 8 ESI+ LysoPC(P-18:1(9Z)) LysoPC(P-18:1) 506.0144 189.8050 9 ESI+ dTDP-4-acetamido-4,6-dideoxy-D-galactose dTDP-4-乙酰胺基-4,6-双脱氧-D-半乳糖 589.1477 475.9890 10 ESI+ LysoPC(18:2(9Z,12Z)) LysoPC(18:2) 520.3412 215.6990 11 ESI+ PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z)) PS(20:5/18:1) 808.5132 216.6680 12 ESI+ alpha-Tocopherolquinone α-托可醌 447.3832 30.8130 13 ESI+ PC(15:0/15:0) PC(15:0/15:0) 705.5821 204.1455 14 ESI+ 1-Methyladenosine 1-甲基腺苷 282.0133 277.1165 15 ESI+ Taurine 牛磺酸 126.0218 184.7230 16 ESI+ L-Isoleucine L-异亮氨酸 132.1020 295.9510 17 ESI+ Cohibin B Cohibin B 577.5199 58.2276 18 ESI+ PE(20:5(5Z,8Z,11Z,14Z,17Z)/P-18:0) PE(20:5/P-18:0) 750.5435 146.6850 19 ESI+ SM(d18:1/24:1(15Z)) SM(d18:1/24:1) 813.6853 202.0390 20 ESI+ PE(16:1(9Z)/P-18:0) PE(16:1/P-18:0) 702.5428 157.1005 21 ESI+ Methylimidazole acetaldehyde 甲基咪唑乙醛 124.0870 47.1570 22 ESI+ PE(20:4(8Z,11Z,14Z,17Z)/P-16:0) PE(20:4/P-16:0) 724.5272 38.9930 23 ESI+ SM(d18:1/16:0) SM(d18:1/16:0) 703.5467 157.1725 24 ESI+ PC(16:0/P-16:0) PC(16:0/P-16:0) 718.5384 164.0870 25 ESI+ PS(22:4(7Z,10Z,13Z,16Z)/18:2(9Z,12Z)) PS(22:4/18:2) 836.5097 172.6190 26 ESI+ 4-Acetamido-2-amino-6-nitrotoluene 4-乙酰氨基-2-氨基-6-硝基甲苯 209.1032 439.989 27 ESI− Galactitol 半乳糖醇 180.8123 356.2440 28 ESI− D-Arabinose 5-phosphate D-阿拉伯糖5-磷酸 228.0274 477.5370 29 ESI− Adenosine 腺苷 265.7674 230.6680 30 ESI− L-Tyrosine L-酪氨酸 180.0657 321.6580 31 ESI− 15-Methylpalmitate 15-甲基棕榈酸酯 269.2485 10.9386 32 ESI− beta-D-Glucosamine β-D-葡萄糖胺 213.0490 391.9780 33 ESI− 16-Methylheptadecanoic acid 16-甲基庚烷酸 283.2638 38.0120 34 ESI− Pantothenic acid 泛酸 217.1188 281.8835 35 ESI− 2-Ketobutyric acid 2-酮基丁酸 101.0233 212.6130 36 ESI− Malonate 丙二酸 103.0582 391.1115 37 ESI− Cholesterol sulfate 硫酸胆固醇 465.2992 58.3416 38 ESI− LysoPA(16:0/0:0) LysoPA(16:0/0:0) 408.2598 58.6062 39 ESI− Adenosine diphosphate ribose 二磷酸腺苷核糖 557.0812 452.6930 40 ESI− L-Gulonolactone L-古洛内酯 176.9801 172.2120 41 ESI− 12-Hydroxydodecanoic acid 12-羟基十二烷酸 214.8324 69.1386 42 ESI− Gamma-Linolenic acid γ-亚麻酸 277.2172 38.5270 ![]() 图 3 小鼠脑组织中差异代谢物的代谢通路分析注:A,C,E为正离子模式下的代谢通路分析图;B,D,F为负离子模式下的代谢通路分析图。Figure 3. Metabolic pathway analysis of differential metabolites in brain samples of mice
图 3 小鼠脑组织中差异代谢物的代谢通路分析注:A,C,E为正离子模式下的代谢通路分析图;B,D,F为负离子模式下的代谢通路分析图。Figure 3. Metabolic pathway analysis of differential metabolites in brain samples of mice2.3 TB对D-半乳糖致衰老小鼠脑组织代谢轮廓的调节作用及代谢途径的分析
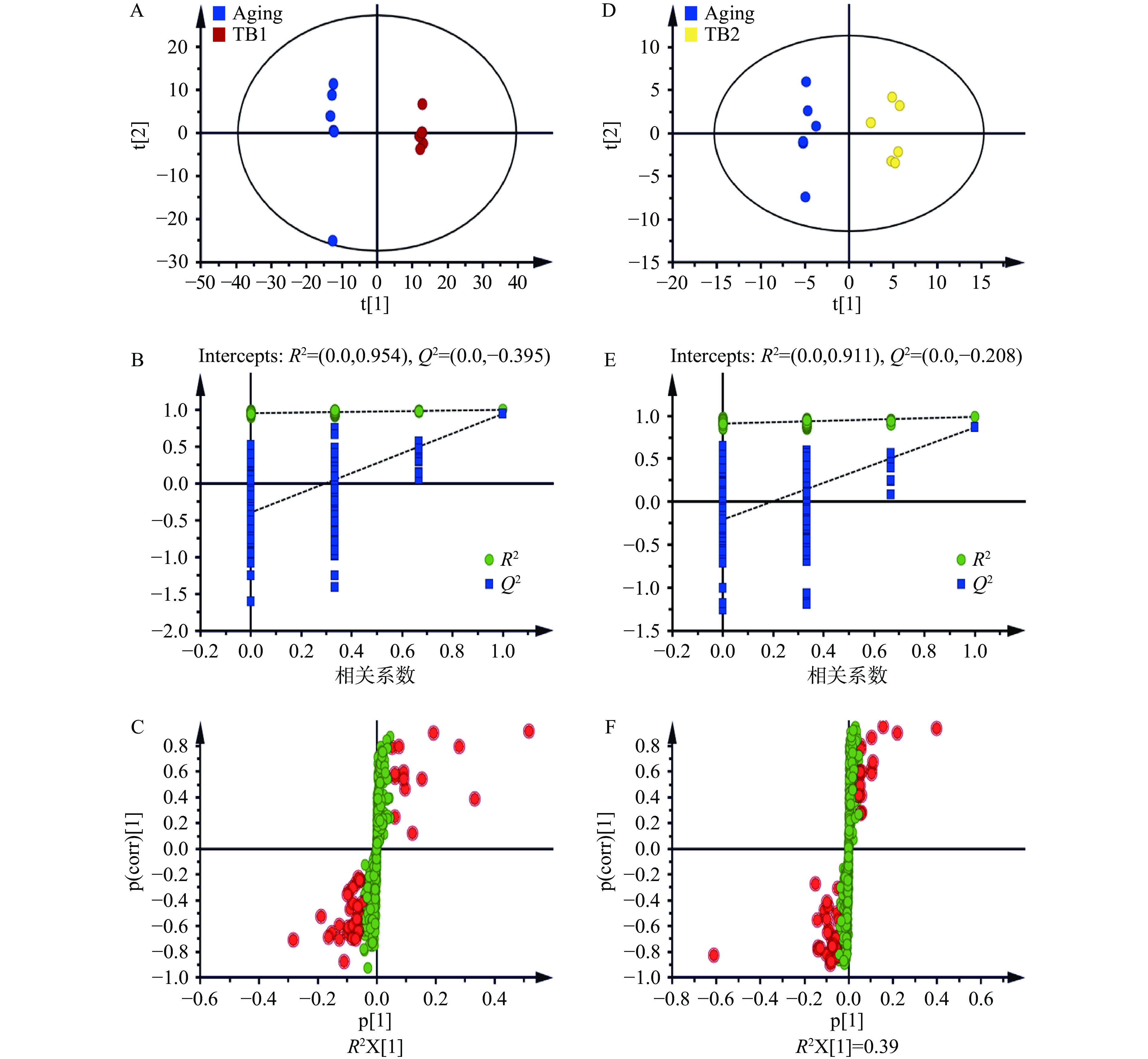
通过OPLS-DA分析对TB1组与D-gal模型组、TB2组与D-gal模型组的脑组织样本数据进行代谢轮廓分析(图4),表明与D-gal模型组相比,给予茶褐素后使小鼠脑组织代谢轮廓发生了改变。由表3可知,TB1组对衰老相关代谢物中的17个物质具有调节作用且具有统计学意义。预防性给予TB1后,D-gal模型组中丙二酸、4-乙酰氨基-2-氨基-6-硝基甲苯、N-乙酰组氨酸等代谢物的含量显著降低(P<0.05);肌肽、硫酸胆固醇、LysoPC(P-18:1(9Z))、PC(15:0/15:0)、PE(22:5(7Z,10Z,13Z,16Z,19Z)/22:4(7Z,10Z,13Z,16Z))、PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z))等代谢物的含量显著升高(P<0.05)。预防性给予TB1主要涉及甘油磷脂代谢(Glycerophospholipid metabolism)、组氨酸代谢(Histidine metabolism)、TCA循环(TCA cycle)、乙醛酸和二羧酸代谢(Glyoxylate and dicarboxylate metabolism)、β-丙氨酸代谢(beta-Alanine metabolism)5条代谢通路(图3C和3D)。TB2组对衰老相关代谢物中的32个物质具有调节作用且具有统计学意义。TB2治疗后,D-gal模型组中dTDP-4-乙酰胺基-4,6-双脱氧-D-半乳糖、α-托可醌、半乳糖醇等代谢物的含量显著降低(P<0.05);胞嘧啶、肌肽、LysoPC(P-18:1(9Z))、(18:2(9Z,12Z))、PE(20:5(5Z,8Z,11Z,14Z,17Z)/P-18:0)、PE(16:1(9Z)/P-18:0)、PC(15:0/15:0)、PC(16:0/P-16:0)、PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z))、硫酸胆固醇等代谢物的含量显著升高(P<0.05)。TB2治疗主要涉及甘油磷脂代谢(Glycerophospholipid metabolism)、组氨酸代谢(Histidine metabolism)、牛磺酸和亚牛磺酸代谢(Taurine and hypotaurine metabolism)、缬氨酸、亮氨酸和异亮氨酸的降解(Valine, leucine and isoleucine degradation)、苯丙氨酸、酪氨酸和色氨酸的生物合成(Phenylalanine, tyrosine and tryptophan biosynthesis)5条代谢通路(图3E和3F)。
![]() 图 4 D-gal模型组和TB组小鼠脑组织的比较分析注:A、D为OPLS-DA图;B、E为置换检验图;C、F为S-plot图。Figure 4. Comparative analysis of mouse brain samples in D-galactose model and TB groups表 3 茶褐素干预衰老小鼠脑组织的差异代谢产物Table 3. Differential metabolites in brain of aging mice interfered by TB
图 4 D-gal模型组和TB组小鼠脑组织的比较分析注:A、D为OPLS-DA图;B、E为置换检验图;C、F为S-plot图。Figure 4. Comparative analysis of mouse brain samples in D-galactose model and TB groups表 3 茶褐素干预衰老小鼠脑组织的差异代谢产物Table 3. Differential metabolites in brain of aging mice interfered by TB编号 代谢物 代谢物中文名称 Aging vs Control TB1 vs Aging TB2 vs Aging 变化
趋势差异
倍数变化
趋势差异
倍数变化
趋势差异
倍数1 Cytosine 胞嘧啶 ↓ 0.8029 ↑ 1.5325 2 Carnosine 肌肽 ↓ 0.4533 ↑ 5.4159 ↑ 5.0120 3 3-Methylglutarylcarnitine 3-甲基戊二酰肉碱 ↓ 0.3732 ↑ 2.9011 4 Aminoadipic acid 氨基己二酸 ↓ 0.7159 ↑ 1.3386 5 PE(22:5(7Z,10Z,13Z,16Z,19Z)/22:4(7Z,10Z,13Z,16Z)) PE(22:5/22:4) ↓ 0.6348 ↑ 1.4000 6 Acetyl-CoA 乙酰辅酶A ↓ 0.5719 ↑ 1.4852 7 N-Acetylhistidine N-乙酰组氨酸 ↑ 1.9021 ↓ 0.5132 8 LysoPC(P-18:1(9Z)) LysoPC(P-18:1) ↓ 0.5983 ↑ 1.4671 ↑ 1.5938 9 dTDP-4-acetamido-4,6-dideoxy-D-galactose dTDP-4-乙酰胺基-4,6-双脱氧-D-半乳糖 ↑ 2.0146 ↓ 0.6199 10 LysoPC(18:2(9Z,12Z)) LysoPC(18:2) ↓ 0.8280 ↑ 1.0170 11 PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z)) PS(20:5/18:1) ↓ 0.4437 ↑ 1.2359 ↑ 4.0043 12 alpha-Tocopherolquinone α-托可醌 ↑ 1.7640 ↓ 0.7212 13 PC(15:0/15:0) PC(15:0/15:0) ↓ 0.6307 ↑ 1.8148 ↑ 1.5431 14 1-Methyladenosine 1-甲基腺苷 ↓ 0.7053 ↑ 1.5229 15 Taurine 牛磺酸 ↓ 0.7418 ↑ 1.7621 16 L-Isoleucine L-异亮氨酸 ↓ 0.7858 ↑ 1.2969 17 Cohibin B Cohibin B ↓ 0.8474 ↓ 0.7069 18 PE(20:5(5Z,8Z,11Z,14Z,17Z)/P-18:0) PE(20:5/P-18:0) ↓ 0.2859 ↑ 1.3156 19 SM(d18:1/24:1(15Z)) SM(d18:1/24:1) ↑ 1.6743 ↑ 1.5704 20 PE(16:1(9Z)/P-18:0) PE(16:1/P-18:0) ↓ 0.5368 ↑ 1.8409 21 Methylimidazole acetaldehyde 甲基咪唑乙醛 ↓ 0.7202 ↑ 1.4419 22 PE(20:4(8Z,11Z,14Z,17Z)/P-16:0) PE(20:4/P-16:0) ↓ 0.3147 ↑ 1.2648 23 SM(d18:1/16:0) SM(d18:1/16:0) ↑ 1.5238 ↑ 2.1357 24 PC(16:0/P-16:0) PC(16:0/P-16:0) ↓ 0.5032 ↑ 1.4610 25 PS(22:4(7Z,10Z,13Z,16Z)/18:2(9Z,12Z)) PS(22:4/18:2) ↓ 0.7539 ↑ 1.3393 26 4-Acetamido-2-amino-6-nitrotoluene 4-乙酰氨基-2-氨基-6-硝基甲苯 ↑ 4.2653 ↓ 0.2134 27 Galactitol 半乳糖醇 ↓ 0.7715 ↓ 0.8221 28 D-Arabinose 5-phosphate D-阿拉伯糖5-磷酸 ↓ 0.6743 ↑ 1.5383 ↑ 1.4582 29 Adenosine 腺苷 ↓ 0.1937 ↓ 0.6391 30 L-Tyrosine L-酪氨酸 ↓ 0.3581 ↑ 1.8098 31 15-Methylpalmitate 15-甲基棕榈酸酯 ↓ 0.8433 ↓ 0.5385 32 beta-D-Glucosamine β-D-葡萄糖胺 ↑ 1.3058 ↓ 0.7841 33 16-Methylheptadecanoic acid 16-甲基庚烷酸 ↓ 0.5698 ↓ 0.7048 34 Pantothenic acid 泛酸 ↑ 1.4896 ↑ 1.5618 35 2-Ketobutyric acid 2-酮基丁酸 ↓ 0.8280 ↑ 1.2947 36 Malonate 丙二酸 ↑ 1.4562 ↓ 0.6996 37 Cholesterol sulfate 硫酸胆固醇 ↓ 0.1937 ↑ 1.3156 ↑ 1.4716 38 LysoPA(16:0/0:0) LysoPA(16:0/0:0) ↑ 0.6971 ↑ 1.6510 39 Adenosine diphosphate ribose 二磷酸腺苷核糖 ↓ 0.3581 ↑ 4.9166 ↑ 4.8489 40 L-Gulonolactone L-古洛内酯 ↓ 0.8433 ↑ 1.2209 41 12-Hydroxydodecanoic acid 12-羟基十二烷酸 ↓ 0.6902 ↑ 1.4131 42 Gamma-Linolenic acid γ-亚麻酸 ↓ 0.8724 ↓ 0.7663 注:“↑”和“↓”表示上调和下调。 整合茶褐素(TB1和TB2组)干预后衰老小鼠脑组织中与抗衰老相关的代谢物。TB1与D-gal模型组相比,包括PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z))、PC(15:0/15:0)、LysoPC(P-18:1(9Z))等5种甘油磷脂类,氨基己二酸和N-乙酰组氨酸2种氨基酸,乙酰辅酶A和γ-亚麻酸2种脂肪酰类以及少量甾体(硫酸胆固醇)、脂肪酸类(3-甲基戊二酰肉碱)、醇类(泛酸)、脂类(二磷酸腺苷核糖)和其他类。TB2与D-gal模型组相比,包括PE(20:5(5Z,8Z,11Z,14Z,17Z)/P-18:0)、(16:1(9Z)/P-18:0)、LysoPC(P-18:1(9Z))、(18:2(9Z,12Z))、PC(16:0/P-16:0)、(15:0/15:0)等9种甘油磷脂类,胞嘧啶和腺苷2种核苷、SM(d18:1/24:1(15Z))、(d18:1/16:0)2种鞘脂类、D-阿拉伯糖5-磷酸和β-D-葡萄糖胺2种糖类、牛磺酸和2-酮基丁酸2种有机酸以及少量脂肪酸类(16-甲基庚烷酸)、甾体(硫酸胆固醇)、氨基酸(L-异亮氨酸)和其他类。其中有部分代谢物及代谢通路为TB1和TB2组共有:代谢物为肌肽、LysoPC(P-18:1(9Z))、PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z))、PC(15:0/15:0)、D-阿拉伯糖-5-磷酸盐、硫酸胆固醇和二磷酸腺苷核糖,代谢通路为甘油磷脂代谢和组氨酸代谢。
3. 讨论
脑衰老表现为认知功能低下和中枢胆碱能系统功能下降。ChAT是一种重要的神经递质乙酰胆碱的合成酶,也是胆碱能神经元的标志物,在一定程度上可以反映胆碱能系统功能状况[10-11]。本研究发现茶褐素组的胆碱能神经元的数目明显增加,结合前期研究提示茶褐素能够改善D-gal致衰老小鼠的空间学习记忆能力[6],因此,推断茶褐素可能是通过增强海马胆碱能系统的功能,从而保护胆碱能神经元,改善衰老小鼠的学习记忆能力。
磷脂酰胆碱(Phosphatidylcholine,PC)、磷脂酰乙醇胺(Phosphatidyl ethanlamine,PE)和磷脂酰丝氨酸(Phosphatidylserine,PS)脂类物质都是甘油磷脂代谢重要的相关物质,与体内自由基的产生、氧化应激、老化速度等密切相关[12-13]。PC在脑神经细胞中约占17%~20%[14-16]。LysoPCs类物质的含量随年龄增加而减少,这一变化一定程度上意味着磷脂代谢发生紊乱[17-18]。PS在神经系统和代谢过程中发挥至关重要的作用,其通过影响神经传导物质的代谢,激活细胞膜上的多种酶,从而改善大脑机能、提高大脑认知[19-20]、修复大脑损伤[21-23]。本研究中,灌胃茶褐素后,显著提高D-gal致衰老小鼠体内LysoPC(P-18:1(9Z))、PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z))及PC(15:0/15:0)的含量(差异倍数TB1组为1.4671、1.2359、1.8148,TB2组为1.5938、4.0043、1.5431且P<0.05),揭示茶褐素可升高脂类物质含量,改善衰老小鼠脑部功能,通过调节甘油磷酸代谢起到抗衰老的作用。
肌肽(Carnosine)具有许多有益的抗氧化特性,能延缓衰老[24]。本研究发现灌胃茶褐素后,显著提高D-gal致衰老小鼠体内肌肽的含量(差异倍数TB1组为5.4159,TB2组为5.0120且P<0.05),表明茶褐素可通过升高肌肽含量,从而使紊乱的组氨酸代谢得到恢复。
硫酸胆固醇(Cholesterol sulfate)可通过减少氧化应激来调节脑代谢和发挥神经保护效应[25]。本研究发现灌胃茶褐素后,显著提高D-gal致衰老小鼠体内硫酸胆固醇的含量(差异倍数TB1组为1.3156,TB2组为1.4716且P<0.05)。因此,茶褐素对硫酸胆固醇的作用可能有助于降低脑内氧化应激水平,进而延缓衰老。
乙酰辅酶A(Acetyl-CoA)是预防性给予茶褐素后的特征代谢物。乙酰辅酶A是能源物质经TCA循环代谢的重要中间产物,提高其水平有望逆转大脑衰老[26-27]。本研究预防性给予茶褐素后,乙酰辅酶A含量升高,推测茶褐素可能对TCA循环代谢有调控作用。
L-异亮氨酸(L-Isoleucine)、牛磺酸(Taurine)和L-酪氨酸(L-Tyrosine)是茶褐素治疗后的特征代谢物。支链氨基酸可以明显延长大鼠的寿命,并减轻多种组织器官的炎症[28]。本研究发现由D-gal致衰老小鼠脑组织中L-异亮氨酸含量显著降低;给予茶褐素治疗后,L-异亮氨酸含量增加,从而调节缬氨酸、亮氨酸和异亮氨酸的降解通路。牛磺酸是一种游离氨基酸,具有较强抗氧化活性,其也是一种重要的神经递质,参与大脑的发育,与认知能力相关[29-30]。本研究发现茶褐素治疗后,牛磺酸含量升高,有效改善D-gal致衰老小鼠机体内的牛磺酸和亚牛磺酸代谢紊乱。L-酪氨酸可来源于苯丙氨酸,参与能量代谢和清除自由基。苯丙氨酸经羟化作用生成酪氨酸,酪氨酸进一步经酪氨酸羟化酶作用生成神经递质[31-32]。神经递质是连接脑内神经元突触之间的信使,主要发挥信息传递功能[33]。本研究结果表明茶褐素有效抑制D-gal引起的苯丙氨酸、酪氨酸和色氨酸的生物合成代谢紊乱,恢复L-酪氨酸水平。
4. 结论
基于UHPLC-QE-MS代谢组学方法,并结合多元统计学比较研究了D-gal致衰老小鼠在茶褐素干预下的脑组织代谢变化,预防性给予茶褐素筛选出17个显著差异代谢物及其参与的5条代谢通路,茶褐素治疗筛选出32个显著差异代谢物及其参与的5条代谢通路。预防和治疗两种干预方式对衰老小鼠脑组织代谢都有一定调节作用,二者存在共同和差异的代谢物、作用代谢途径,主要涉及甘油磷脂代谢和组氨酸代谢2条代谢通路。总之,长期饮用茶褐素具有抗衰老作用。
-
![]()
图 1 各组小鼠脑组织ChAT免疫组化结果
注:放大倍数:400×;不同字母表示差异显著(P<0.05)。
Figure 1. ChAT immunohistochemical results of brain tissues in each group
![]()
图 2 不同组小鼠脑组织的比较分析
注:A为不同组小鼠脑组织的PLS-DA散点图;B为对照组和D-gal模型组小鼠脑组织OPLS-DA得分图;C为置换检验图;D为S-plot图;图D中红色的点表明这些代谢物的VIP值大于等于1,绿色的点表示这些代谢物的VIP值小于1。
Figure 2. Comparative analysis of mouse brain samples in different groups
![]()
图 3 小鼠脑组织中差异代谢物的代谢通路分析
注:A,C,E为正离子模式下的代谢通路分析图;B,D,F为负离子模式下的代谢通路分析图。
Figure 3. Metabolic pathway analysis of differential metabolites in brain samples of mice
![]()
图 4 D-gal模型组和TB组小鼠脑组织的比较分析
注:A、D为OPLS-DA图;B、E为置换检验图;C、F为S-plot图。
Figure 4. Comparative analysis of mouse brain samples in D-galactose model and TB groups
表 1 QC样品中内标响应稳定性
Table 1 Stability of internal standard response in QC samples
内标种类 正离子模式 负离子模式 m/z R.T. RSD m/z R.T. RSD L-2-氯苯丙氨酸 200.0473 273.324 5.8% 198.0325 264.603 4.6%  下载: 导出CSV
下载: 导出CSV
表 2
D-gal诱导衰老小鼠脑组织的差异代谢产物 Table 2 Differential metabolites in brain of aging mice induced by D-gal
编号 正/负离子模式 代谢物 代谢物
中文名称质荷比 保留时间(min) 1 ESI+ Cytosine 胞嘧啶 112.0507 478.6850 2 ESI+ Carnosine 肌肽 226.8889 439.9890 3 ESI+ 3-Methylglutarylcarnitine 3-甲基戊二酰肉碱 290.9379 442.9370 4 ESI+ Aminoadipic acid 氨基己二酸 161.1285 460.1125 5 ESI+ PE(22:5(7Z,10Z,13Z,16Z,19Z)/22:4(7Z,10Z,13Z,16Z)) PE(22:5/22:4) 842.0741 148.3530 6 ESI+ Acetyl-CoA 乙酰辅酶A 810.1326 451.3795 7 ESI+ N-Acetylhistidine N-乙酰组氨酸 197.1030 457.3510 8 ESI+ LysoPC(P-18:1(9Z)) LysoPC(P-18:1) 506.0144 189.8050 9 ESI+ dTDP-4-acetamido-4,6-dideoxy-D-galactose dTDP-4-乙酰胺基-4,6-双脱氧-D-半乳糖 589.1477 475.9890 10 ESI+ LysoPC(18:2(9Z,12Z)) LysoPC(18:2) 520.3412 215.6990 11 ESI+ PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z)) PS(20:5/18:1) 808.5132 216.6680 12 ESI+ alpha-Tocopherolquinone α-托可醌 447.3832 30.8130 13 ESI+ PC(15:0/15:0) PC(15:0/15:0) 705.5821 204.1455 14 ESI+ 1-Methyladenosine 1-甲基腺苷 282.0133 277.1165 15 ESI+ Taurine 牛磺酸 126.0218 184.7230 16 ESI+ L-Isoleucine L-异亮氨酸 132.1020 295.9510 17 ESI+ Cohibin B Cohibin B 577.5199 58.2276 18 ESI+ PE(20:5(5Z,8Z,11Z,14Z,17Z)/P-18:0) PE(20:5/P-18:0) 750.5435 146.6850 19 ESI+ SM(d18:1/24:1(15Z)) SM(d18:1/24:1) 813.6853 202.0390 20 ESI+ PE(16:1(9Z)/P-18:0) PE(16:1/P-18:0) 702.5428 157.1005 21 ESI+ Methylimidazole acetaldehyde 甲基咪唑乙醛 124.0870 47.1570 22 ESI+ PE(20:4(8Z,11Z,14Z,17Z)/P-16:0) PE(20:4/P-16:0) 724.5272 38.9930 23 ESI+ SM(d18:1/16:0) SM(d18:1/16:0) 703.5467 157.1725 24 ESI+ PC(16:0/P-16:0) PC(16:0/P-16:0) 718.5384 164.0870 25 ESI+ PS(22:4(7Z,10Z,13Z,16Z)/18:2(9Z,12Z)) PS(22:4/18:2) 836.5097 172.6190 26 ESI+ 4-Acetamido-2-amino-6-nitrotoluene 4-乙酰氨基-2-氨基-6-硝基甲苯 209.1032 439.989 27 ESI− Galactitol 半乳糖醇 180.8123 356.2440 28 ESI− D-Arabinose 5-phosphate D-阿拉伯糖5-磷酸 228.0274 477.5370 29 ESI− Adenosine 腺苷 265.7674 230.6680 30 ESI− L-Tyrosine L-酪氨酸 180.0657 321.6580 31 ESI− 15-Methylpalmitate 15-甲基棕榈酸酯 269.2485 10.9386 32 ESI− beta-D-Glucosamine β-D-葡萄糖胺 213.0490 391.9780 33 ESI− 16-Methylheptadecanoic acid 16-甲基庚烷酸 283.2638 38.0120 34 ESI− Pantothenic acid 泛酸 217.1188 281.8835 35 ESI− 2-Ketobutyric acid 2-酮基丁酸 101.0233 212.6130 36 ESI− Malonate 丙二酸 103.0582 391.1115 37 ESI− Cholesterol sulfate 硫酸胆固醇 465.2992 58.3416 38 ESI− LysoPA(16:0/0:0) LysoPA(16:0/0:0) 408.2598 58.6062 39 ESI− Adenosine diphosphate ribose 二磷酸腺苷核糖 557.0812 452.6930 40 ESI− L-Gulonolactone L-古洛内酯 176.9801 172.2120 41 ESI− 12-Hydroxydodecanoic acid 12-羟基十二烷酸 214.8324 69.1386 42 ESI− Gamma-Linolenic acid γ-亚麻酸 277.2172 38.5270
下载: 导出CSV
表 3 茶褐素干预衰老小鼠脑组织的差异代谢产物
Table 3 Differential metabolites in brain of aging mice interfered by TB
编号 代谢物 代谢物中文名称 Aging vs Control TB1 vs Aging TB2 vs Aging 变化
趋势差异
倍数变化
趋势差异
倍数变化
趋势差异
倍数1 Cytosine 胞嘧啶 ↓ 0.8029 ↑ 1.5325 2 Carnosine 肌肽 ↓ 0.4533 ↑ 5.4159 ↑ 5.0120 3 3-Methylglutarylcarnitine 3-甲基戊二酰肉碱 ↓ 0.3732 ↑ 2.9011 4 Aminoadipic acid 氨基己二酸 ↓ 0.7159 ↑ 1.3386 5 PE(22:5(7Z,10Z,13Z,16Z,19Z)/22:4(7Z,10Z,13Z,16Z)) PE(22:5/22:4) ↓ 0.6348 ↑ 1.4000 6 Acetyl-CoA 乙酰辅酶A ↓ 0.5719 ↑ 1.4852 7 N-Acetylhistidine N-乙酰组氨酸 ↑ 1.9021 ↓ 0.5132 8 LysoPC(P-18:1(9Z)) LysoPC(P-18:1) ↓ 0.5983 ↑ 1.4671 ↑ 1.5938 9 dTDP-4-acetamido-4,6-dideoxy-D-galactose dTDP-4-乙酰胺基-4,6-双脱氧-D-半乳糖 ↑ 2.0146 ↓ 0.6199 10 LysoPC(18:2(9Z,12Z)) LysoPC(18:2) ↓ 0.8280 ↑ 1.0170 11 PS(20:5(5Z,8Z,11Z,14Z,17Z)/18:1(9Z)) PS(20:5/18:1) ↓ 0.4437 ↑ 1.2359 ↑ 4.0043 12 alpha-Tocopherolquinone α-托可醌 ↑ 1.7640 ↓ 0.7212 13 PC(15:0/15:0) PC(15:0/15:0) ↓ 0.6307 ↑ 1.8148 ↑ 1.5431 14 1-Methyladenosine 1-甲基腺苷 ↓ 0.7053 ↑ 1.5229 15 Taurine 牛磺酸 ↓ 0.7418 ↑ 1.7621 16 L-Isoleucine L-异亮氨酸 ↓ 0.7858 ↑ 1.2969 17 Cohibin B Cohibin B ↓ 0.8474 ↓ 0.7069 18 PE(20:5(5Z,8Z,11Z,14Z,17Z)/P-18:0) PE(20:5/P-18:0) ↓ 0.2859 ↑ 1.3156 19 SM(d18:1/24:1(15Z)) SM(d18:1/24:1) ↑ 1.6743 ↑ 1.5704 20 PE(16:1(9Z)/P-18:0) PE(16:1/P-18:0) ↓ 0.5368 ↑ 1.8409 21 Methylimidazole acetaldehyde 甲基咪唑乙醛 ↓ 0.7202 ↑ 1.4419 22 PE(20:4(8Z,11Z,14Z,17Z)/P-16:0) PE(20:4/P-16:0) ↓ 0.3147 ↑ 1.2648 23 SM(d18:1/16:0) SM(d18:1/16:0) ↑ 1.5238 ↑ 2.1357 24 PC(16:0/P-16:0) PC(16:0/P-16:0) ↓ 0.5032 ↑ 1.4610 25 PS(22:4(7Z,10Z,13Z,16Z)/18:2(9Z,12Z)) PS(22:4/18:2) ↓ 0.7539 ↑ 1.3393 26 4-Acetamido-2-amino-6-nitrotoluene 4-乙酰氨基-2-氨基-6-硝基甲苯 ↑ 4.2653 ↓ 0.2134 27 Galactitol 半乳糖醇 ↓ 0.7715 ↓ 0.8221 28 D-Arabinose 5-phosphate D-阿拉伯糖5-磷酸 ↓ 0.6743 ↑ 1.5383 ↑ 1.4582 29 Adenosine 腺苷 ↓ 0.1937 ↓ 0.6391 30 L-Tyrosine L-酪氨酸 ↓ 0.3581 ↑ 1.8098 31 15-Methylpalmitate 15-甲基棕榈酸酯 ↓ 0.8433 ↓ 0.5385 32 beta-D-Glucosamine β-D-葡萄糖胺 ↑ 1.3058 ↓ 0.7841 33 16-Methylheptadecanoic acid 16-甲基庚烷酸 ↓ 0.5698 ↓ 0.7048 34 Pantothenic acid 泛酸 ↑ 1.4896 ↑ 1.5618 35 2-Ketobutyric acid 2-酮基丁酸 ↓ 0.8280 ↑ 1.2947 36 Malonate 丙二酸 ↑ 1.4562 ↓ 0.6996 37 Cholesterol sulfate 硫酸胆固醇 ↓ 0.1937 ↑ 1.3156 ↑ 1.4716 38 LysoPA(16:0/0:0) LysoPA(16:0/0:0) ↑ 0.6971 ↑ 1.6510 39 Adenosine diphosphate ribose 二磷酸腺苷核糖 ↓ 0.3581 ↑ 4.9166 ↑ 4.8489 40 L-Gulonolactone L-古洛内酯 ↓ 0.8433 ↑ 1.2209 41 12-Hydroxydodecanoic acid 12-羟基十二烷酸 ↓ 0.6902 ↑ 1.4131 42 Gamma-Linolenic acid γ-亚麻酸 ↓ 0.8724 ↓ 0.7663 注:“↑”和“↓”表示上调和下调。
下载: 导出CSV
-
[1] LIANG Y, ZHANG L, LU J. A study on chemical estimation of Pu-erh tea quality[J]. Journal of the Science of Food & Agriculture,2005,85(3):381−390.
[2] ZHAO Z J, TONG H R, ZHOU L, et al. Fungal colonization of Pu-erh tea in Yunnan[J]. Journal of Food Safety,2010,30(4):769−784. doi: 10.1111/j.1745-4565.2010.00240.x
[3] QIN Y, GONG J S, ZHANG H F, et al. Extraction technology of theabrownine from Pu-erh tea and its physico-chemical properties[J]. Chemistry & Industry of Forest Products,2009,29(5):95−98.
[4] MARGALEF M, IGLESISS-CARRES L, PONS Z, et al. Age related differences in the plasma kinetics of flavanols in rats[J]. Journal of Nutritional Biochemistry,2016,29:90−96. doi: 10.1016/j.jnutbio.2015.11.007
[5] JADOON S, KARIM S, ASAD M H H B, et al. Anti-aging potential of phytoextract loaded-pharmaceutical creams for human skin cell longetivity[J]. Oxida Med Cell Longe,2015(1):709628.
[6] LEI S W, ZHANG Z F, XIE G H, et al. Theabrownin modulates the gut microbiome and serum metabolome in aging mice induced by D-galactose[J]. Journal of Functional Foods,2022,89:104941. doi: 10.1016/j.jff.2022.104941
[7] 周曼丽, 俞赟丰, 罗晓欣, 等. 基于代谢组学技术的冠心病血瘀证不同进展阶段大鼠潜在生物标志物及代谢通路研究[J]. 中国中医药信息杂志,2022,29(1):73−79. [ZHOU M L, YU Y F, LUO X X, et al. Study on potential biomarkers and metabolic pathways of rats in different progressive stages of coronary heart disease with blood stasis syndrome based on metabonomics technology[J]. Chinese Journal of Information on TCM,2022,29(1):73−79. [8] 岳随娟, 刘建, 龚加顺. 普洱茶茶褐素对大鼠肠道菌群的影响[J]. 茶叶科学,2016,36(3):261−267. [YUE S J, LIU J, GONG J S. Effects of theabrownin extracted from Pu-erh tea on the intestinal flora[J]. Journal of Tea Science,2016,36(3):261−267. doi: 10.3969/j.issn.1000-369X.2016.03.006 [9] 王丽英, 杨丽, 朱建华. 康复新对去卵巢大鼠学习记忆能力和基底前脑胆碱能神经元的影响[J]. 解剖学研究,2021,43(5):479−483. [WANG L Y, YANG L, ZHU J H. Effects of Kangfuxin on learning and memory ability and basal forebrain cholinergic neurons of ovariectomized rats[J]. Anatomy Research,2021,43(5):479−483. doi: 10.3969/j.issn.1671-0770.2021.05.001 [10] 陈庆林, 陈勤, 金蓓蓓. 远志皂苷对AD小鼠学习记忆能力及中枢胆碱能系统标志酶活性的影响[J]. 中药药理与临床,2011,27(3):33−36. [CHEN Q L, CHEN Q, JIN B B. The influence of tenuigenin on learning-memory and the activity of marker enzymes of central cholinergic system in AD mice[J]. Chinese Remedies & Clinics,2011,27(3):33−36. doi: 10.13412/j.cnki.zyyl.2011.03.015 [11] 许勇, 李金莲, 王桂华, 等. 不同类型茶叶水提取液对衰老小鼠学习记忆和胆碱能神经功能的影响[J]. 滨州医学院学报,2017,40(5):340−342. [XU Y, LI J L, WANG G H, et al. The effects of tea leaves on learing, memory and cholinergic system in aged mice[J]. Journal of Binzhou Medical University,2017,40(5):340−342. doi: 10.3969/j.issn.1001-9510.2017.05.006 [12] OAKMAN C, TENORI L, BIGANZOLI L, et al. Uncovering the metabolomic fingerprint of breast cancer[J]. International Journal of Biochemistry & Cell Biology,2011,43(7):1010−1020.
[13] WAI M, WONG K, BRAIDY N, et al. Dysregulation of lipids in Alzheimer's disease and their role as potential biomarkers[J]. Alzheimers & Dementia the Journal of the Alzheimer’s Association,2017,13(7):810−827.
[14] KIM S T, CHUNG Y H, LEE H S, et al. Protective effects of phosphatidylcholine on oxaliplatin-induced neuropathy in rats[J]. Life Sciences,2015,130:81−87. doi: 10.1016/j.lfs.2015.03.013
[15] SI X, STRAPPE P, BLANCHARD C, et al. Enhanced anti-obesity effects of complex of resistant starch and chitosan in high fat diet fed rats[J]. Carbohydrate Polymers,2017,157:834−841. doi: 10.1016/j.carbpol.2016.10.042
[16] ZHANG J, WEI S Y, YUAN L, et al. Davunetide improves spatial learning and memory in Alzheimer’s disease-associated rats[J]. Physiology & Behavior,2017,174:67−73.
[17] ZHANG L X, HAN X F, LI Z, et al. Metabolomics research on time-selected combination of Liuwei Dihuang and Jinkui Shenqi pills in treating kidney deficiency and aging by chemometric methods[J]. Chemometrics and Intelligent Laboratory Systems,2014,130(1):50−57.
[18] SCHNEIDER M, LEVANT B, REICHEL M, et al. Lipids in psychiatric disorders and preventive medicine[J]. Neuroscience & Biobehavioral Reviews,2017,76:336−362.
[19] LIU S H, CHANG C D, CHEN P H, et al. Docosahexaenoic acid and phosphatidylserine supplementations improve antioxidant activities and cognitive functions of the developing brain on pentylenetetrazol-induced seizure model[J]. Brain Research,2012,1451:19−26. doi: 10.1016/j.brainres.2012.02.060
[20] ZHANG Y Y, YANG L Q, GUO L M. Effect of phosphatidylserine on memory in patients and rats with Alzheimer’s disease[J]. Genetics and Molecular Research,2015,14(3):9325−9333. doi: 10.4238/2015.August.10.13
[21] 王东升, 刘英姿, 刘莹莹, 等. 脑梗死与磷脂酰丝氨酸相关性的研究进展[J]. 中华老年心脑血管病杂志, 2017, 19(8): 887-889 WANG D S, LIU Y Z, LIU Y Y, et al. Research progress of correlation between cerebral infarction and phosphatidylserine[J]. Chinese Journal of Geriatric Heart Brain and Vessel Diseases, 2017, 19(8): 887-889.
[22] 刘慧燕, 张少敏, 沈会平, 等. 大豆磷脂酰丝氨酸对力竭运动小鼠心肌抗氧化的影响[J]. 中国食品添加剂,2016(3):103−106. [LIU H Y, ZHANG S M, SHEN H P, et al. The anti-oxidant effects of soybean phosphatidylserine on myocardium in exhaustive swimming mice[J]. China Food Additives,2016(3):103−106. doi: 10.3969/j.issn.1006-2513.2016.03.008 [23] 张赟彬, 高雅, 魏蒙月, 等. 磷脂酰丝氨酸在乳粉中应用的研究[J]. 食品工业,2017,38(8):173−176. [ZHANG Y B, GAO Y, WEI M Y, et al. An experiment on the formula powder containing phosphatidylserine[J]. The Food Industry,2017,38(8):173−176. [24] JUKIC I, K N, STUPIN A, et al. Carnosine, small but mighty-prospect of use as functional ingredient for functional food formulation[J]. Antioxidants (Basel, Switzerland),2021,10(7):1037.
[25] 蒋悦庭, 倪佳英, 郭沈睿, 等. 硫酸胆固醇的生理功能及其在相关疾病中的作用[J]. 上海交通大学学报:医学版,2021,41(3):5. [JIANG Y T, NI J Y, GUO S R, et al. Physiological function of cholesterol sulfate and its role in related diseases[J]. Journal of Shanghai Jiao Tong University (Medical Science),2021,41(3):5. [26] CHEN W P, YU X L, WU Y S, et al. The SESAME complex regulates cell senescence through the generation of acetyl-CoA[J]. Nature Metabolism,2021,3:983−1000. doi: 10.1038/s42255-021-00412-9
[27] CURRAIS A, HUANG L, GOLDBERG J, et al. Elevating acetyl-CoA levels reduces aspects of brain aging[J]. Elife,2019,19(8):47866.
[28] 姚赫. 支链氨基酸对机体胰岛素抵抗和炎性衰老的影响及其作用机制[D]. 北京: 卫生部北京老年医学研究所, 2021 YAO H. Effect and mechanism of branched chain amino acids on insulin resistance and inflammatory aging[D]. Beijing: Beijing Institute of Geriatrics, Ministry of Health, 2021.
[29] 赵凡凡. 甘草对D-gal诱导的大鼠抗衰老作用及基于牛磺酸通路的靶标代谢组学研究[D]. 太原: 山西大学, 2018 ZHAO F F. The effect of Glycyrrhiza uralensis Fisch on D-gal induced aging rats and the analysis of the target metabolomics based on taurine pathway[D]. Taiyuan: Shanxi University, 2018.
[30] 徐得莱, 苏存锦, 施爱明, 等. 部分抗衰老药物研究进展[J]. 医药导报,2022,41(3):361−366. [XU D L, SU C J, SHI A M, et al. Research progress of some anti-aging drugs[J]. Herald of Medicine,2022,41(3):361−366. [31] 李永彪, 吴兰兰, 樊玉青, 等. 蜘蛛香环烯醚萜部位干预CUMS致抑郁小鼠脑组织的1H-NMR代谢组学分析[J]. 中国实验方剂学杂志,2020,26(19):195−203. [LI Y B, WU L L, FAN Y Q, et al. 1H-NMR metabolomic analysis of brain tissue of CUMS-induced depressed mice intervened by iridoid part of valerianae jatamansi rhizome et radix[J]. Chinese Journal of Experimental Traditional Medical Formulae,2020,26(19):195−203. [32] HE W, WU G. Metabolism of amino acids in the brain and their roles in regulating food intake[J]. Advances in Experimental Medicine and Biology,2020,1265:167−185.
[33] 王丹, 赵瑞珍, 李小黎, 等. 益肾调气法对血管性痴呆模型大鼠脑单胺神经递质的干预作用[J]. 世界中医药,2020,15(10):1395−1399. [WANG D, ZHAO R Z, LI X L, et al. Intervention effects of tonifying kidney and regulating qi method on cerebral monoamine neurotransmitters in model rats with vascular dementia[J]. World Chinese Medicine,2020,15(10):1395−1399. doi: 10.3969/j.issn.1673-7202.2020.10.005 -
期刊类型引用(9)
1. 李杰,马丹丹. 以正磷酸盐为酯化剂制备磷酸酯玉米淀粉及其对Ni~(2+)的吸附性能. 印染助剂. 2025(02): 36-41 .  百度学术
百度学术
2. 安鸿雁,刘馨远,赵国兴,李义. 蜡质玉米淀粉乙酰化的工艺研究. 当代化工. 2023(02): 477-480 . 百度学术
3. 孔刘娟,段元良,栾庆民,王欣彤,袁世英,李新华,刘开昌,李克文. 食用变性淀粉研究进展. 精细与专用化学品. 2023(03): 24-29 . 百度学术
4. 西芳,刘馨远,曹国强. 蜡质玉米淀粉制备乙酰化二淀粉磷酸酯的工艺优化. 广州化工. 2023(08): 80-82+104 . 百度学术
5. 肖志刚,邵晨,杨柳,杨庆余,朱旻鹏,杨宏黎. 淀粉改性方法的研究现状及进展. 农产品加工. 2020(03): 81-84+88 . 百度学术
6. 爨珊珊,石海信,王爱荣,欧春兰,耿昊天,王小丽. 两性淀粉制备与应用的机理及研究进展. 化工技术与开发. 2019(02): 11-16+33 . 百度学术
7. 刘丹,邓利玲,郑连姬,钟耕. 高取代度磷酸魔芋葡甘聚糖酯的制备. 食品与发酵工业. 2019(08): 149-155 . 百度学术
8. 沈艳琴,杨树,武海良,王忠梁. 浆纱用中低温水溶性淀粉浆料的研究进展. 纺织学报. 2019(06): 143-152 . 百度学术
9. 冯琳,黄祖强,孙宁钊,胡华宇,张燕娟,玉琼广. 三聚磷酸钠对醋酸酯淀粉反应效率的影响及工艺优化. 中国食品添加剂. 2018(12): 149-155 . 百度学术
其他类型引用(6)
 下载:
下载:




计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 15



