


Screening of an Alcohol Tolerant and High-yield L-lactic Acid Strain and Optimization of Culture Medium
-
摘要: 为提高L-乳酸产量,降低L-乳酸的生产成本,该研究经过筛选、驯化获得一株耐酒精且高产L-乳酸的菌株鼠李糖乳杆菌AK-0779。使用玉米酒糟代替部分酵母粉作为菌株AK-0779发酵培养基的氮源。在单因素实验基础上,对葡萄糖添加量、酵母粉添加量和玉米酒糟添加量进行三因素三水平响应面优化试验。结果表明,最适发酵培养基为:葡萄糖添加量9.80%,玉米酒糟添加量0.98%,酵母粉添加量1.72%,L-乳酸产量为78.91 g/L,糖酸转换率为80.52%。与酵母粉完全充当氮源产L-乳酸82.36 g/L相比,产量无显著差异,说明玉米酒糟能有效代替部分酵母粉作为发酵培养基的氮源,降低L-乳酸生产成本。Abstract: In order to increase the yield of L-lactic acid and reduce the production cost of L-lactic acid, a strain of Lactobacillus rhamnosus AK-0779 with alcohol tolerance and high yield of L-lactic acid was screened and domesticated. Corn distiller's grains were used as the nitrogen source for the fermentation medium of strain AK-0779 instead of part of yeast powder. On the basis of single factor experiments, three factors and three levels response surface optimization experiments were carried out on the addition of glucose, yeast powder and corn distiller's grains. The results showed that the optimal fermentation medium was glucose 9.80%, corn distiller's grains 0.98%, yeast powder 1.72%, L-lactic acid yield 78.91 g/L and sugar acid conversion rate 80.52%. There was no significant difference in the yield of L-lactic acid compared with yeast powder, which was 82.36 g/L, indicating that corn distiller's grains could effectively replace part of yeast powder as the nitrogen source of fermentation medium and reduce the production cost of L-lactic acid.
-
L-乳酸是一种重要的有机酸,广泛应用于食品、皮革、医药、化工和农业等行业[1-2],需求量巨大。特别是近年来,聚乳酸是一种基于L-乳酸的聚合物,作为一种无毒、可生物降解的高分子材料,被广泛用于生产可生物降解的塑料,以取代传统的化学塑料,备受国内外关注。随着聚乳酸产业的大力发展,急需高光学纯度的L-乳酸满足市场需求。目前,微生物发酵法是常用的一种生产L-乳酸的方法[3]。乳杆菌由于具有转化率高、产酸能力强等优点被广泛用于乳酸的发酵生产。鼠李糖乳杆菌是产L-乳酸的主要乳杆菌,也是近年来研究的热门菌种。吴晖等[4]通过单因素实验与正交试验确定了鼠李糖乳杆菌L-乳酸发酵培养基主要成分的最佳配方,发酵所得L-乳酸产量为143.60 g/L,糖酸转换率为89.80%,达到了国内先进水平。张为魏等[5]对筛选得到的野生型鼠李糖乳杆菌CLRID10进行响应面试验优化后的L-乳酸产量达104.40 g/L,糖酸转换率为75.22%。李海洋等[6]在鼠李糖乳杆菌利用碳源和氮源能力的研究中,发现菌株在最佳发酵条件下的L-乳酸产量为48.25 g/L,糖酸转换率为80.42%。虽然鼠李糖乳杆菌作为微生物具有良好的产酸特性,且具有原料来源广、生产工艺简单,副产物少等优点[7-9],但对发酵的营养条件要求却比较苛刻[10],导致L-乳酸生产成本较高[11-13]。因此,寻找廉价的农业或林业废弃物来替代生产L-乳酸生产的主要基质是降低L-乳酸生产成本的一个重要途径[14]。
玉米酒糟(Corn distillers)是生产燃料乙醇的副产物,富含粗纤维、脂肪、氨基酸、矿物质、酶类和多种维生素等多种营养物质[15-16]。据统计,每生产1 t燃料乙醇产出约10~12 t的玉米酒糟[17],产量巨大,但只是少部分玉米酒糟经简单干燥处理后被用作动物饲料或堆肥出售[18-21],大部分玉米酒糟在储藏堆放过程中发生腐败变质,造成了周围环境的严重污染,其中的有效成分没有得到充分利用[22],造成资源的严重浪费。因此,如果将玉米酒糟代替部分酵母粉作为微生物发酵生产L-乳酸的氮源,不仅能解决环境污染问题,还能使玉米酒糟得到资源化利用,提高玉米酒糟附加值[23]。但玉米酒糟中含有2.5%~3.5%酒精,会抑制发酵微生物的生长繁殖[24],从而影响L-乳酸的生产,因此本研究首先经过筛选、驯化获得一株耐酒精高产L-乳酸的菌株,然后以玉米酒糟为基质对其发酵培养基进行优化,确定最佳发酵培养基条件。研究用玉米酒糟代替部分氮源,以期降低L-乳酸的生产成本,解决玉米酒糟直接排放对环境的污染问题,推动我国玉米深加工产业的发展。
1. 材料与方法
1.1 材料与仪器
6株初始菌株(6133、6141、6166、6167、乳酸菌1、AK-0779) 均由“吉林省食品生物制造科技创新中心”从自然发酵食品中筛选获得;玉米酒糟 购于吉林长春玉米深加工有限公司;乳酸菌肉汤培养基(MRS)固体培养基 MRS液体培养基中添加2%的琼脂粉;酵母浸膏肉汤(YE)发酵培养基(g/L) 葡萄糖20 g,酵母粉25 g,碳酸钙10 g。配好后,进行121 ℃灭菌20 min,碳酸钙单独灭菌;MRS肉汤 青岛高科技工业园海博生物技术有限公司;葡萄糖 北京化工厂有限责任公司;酵母粉 英国Oxoid公司;重质碳酸钙 天津市科密欧化学试剂有限公司;试剂均为分析纯;L/D试剂盒 南京建成生物工程研究所。
高速台式冷冻离心机(FA1104A) 贝克曼库尔特国际贸易有限公司;多功能酶标仪(Infinile M200) TECAN;超净工作台、高压灭菌锅 上海博讯实业有限公司; DAN测序仪( ABI 3730) 上海土森视觉科技有限公司。
1.2 实验方法
1.2.1 初始菌株培养
将初始菌株以4%的接种量接入MRS液体培养基中进行活化传代,于37 ℃厌氧条件下培养24 h,将酒精耐受能力及产酸增量作为本研究的菌株筛选指标。
1.2.2 初始菌株酒精胁迫试验
将初始菌株以4%的接种量接种于MRS液体培养基,37 ℃下培养24 h。再以相同接种量转接到含有酒精体积分别为0%、5%、10%和15%的MRS液体培养基中培养24 h[25],并利用稀释涂布平板法进行菌落计数,计算存活率(%)。按公式(1)计算菌株存活率,取三次平行试验的平均值。
(1) 式中:A0为未加酒精处理的活菌数(个);A1为加酒精处理后的活菌数(个)。
1.2.3 初始菌株产酸增量测定
将各初始菌株接种于MRS液体培养基,37 ℃下培养24 h,测定不同菌株的产L-乳酸增量。
1.2.4 筛选菌株胁迫条件的驯化
将具有高的酒精耐受能力和高的产L-乳酸能力的菌株作为筛选菌株。将筛选菌株接种于含有5%酒精体积的MRS液体培养基中[26],于37 ℃厌氧条件下培养24 h,多次驯化,计算存活率(取三次平行试验的平均值),直到其存活率达到80%左右,驯化停止。
1.2.5 筛选菌株的鉴定
将经驯化后的筛选菌株送至上海生工公司进行16 S rDNA 测序鉴定。所用仪器:ABI 3730型DAN测序仪;PCR反应体系如下:a. 缓冲液标准的缓冲液含 1 pmol/mL Tris·HCl,其 pH 为 8.3~9.0(室温),而在延伸温度(72 ℃)下,pH接近 7.2 。缓冲液中含有一种二价阳离子,用于激活DNA聚合酶的活性中心,一般使用Mg2+;b. dNTP脱氧核糖核苷三磷酸的缩写,是包括 dATP, dGTP,dTTP,dCTP 等在内的统称,即合成新的 DNA 链的材料。4 种 dNTP 的浓度相等,总浓度一般为 200 pmol/mL (即饱和浓度)。dNTP 可与 Mg2+ 结合,使游离的 Mg2+ 浓度下降,影响 DNA 聚合酶的活性;c. 引物是一段短的单链 DNA 片段,可结合在核酸链上与之互补的区域,其功能是作为核苷酸聚合作用的起始点,DNA 聚合酶可由其 3′端开始合成新的核酸链。引物长度一般以 18~30 bp 为宜,过短则降低特异性,过长则会引起引物间的退火而影响有效扩增,同时也增加引物合成的成本。引物的碱基顺序不能与非扩增区有同源性;d. 模板是一段单链或者双链 DNA,提供用于进行 PCR 扩增的原始信息。对模板的纯度要求低,但不能混有蛋白酶、核酸酶、DNA 聚合酶抑制剂、DNA 结合蛋白等。模板 DNA 应该尽量保持低温保存;e. Taq DNA 聚合酶以 DNA 为复制模板,从将 DNA 由 5' 端点开始复制到 3' 端的酶。DNA 聚合酶的主要活性是催化 DNA 的合成(在具备模板、引物、dNTP 等的情况下)及其相辅的活性。必须一直保存于−20 ℃,内含甘油保存。最适反应温度 72 ℃,即延伸温度,通常在配制PCR体系的最后加入。所用引物:27F:AGAGTTTGATCMTGGCTCAG;1492R:GGTTACCTTGTTACGACTT;PCR条件:预变性94 ℃ 4 min,循环94 ℃ 20 s,55 ℃ 20 s,72 1 min,35个循环,延伸10 min。系统发育树构建所用软件:Editplus和MEGA7。
1.2.6 筛选菌株发酵特性研究
将驯化后的筛选菌株接种到MRS液体培养基中,在37 ℃恒温培养箱中发酵0、12、24、36、48、60 h。用生理盐水进行10倍系列稀释后涂布于MRS固体培养基中,记录菌落数(取三次平行试验的平均值),同时利用L-乳酸试剂盒测定以上不同发酵时间的L-乳酸产量(详细测定方法见试剂盒说明书)。L-乳酸计算公式:L-乳酸含量(mmol/L)=(测定OD值-空白OD值)/(标准OD值-空白OD值)×标准品浓度×样本测试前稀释倍数。
1.2.7 筛选菌株产酸稳定性试验
将驯化后的筛选菌株接种到MRS液体培养基中,于37 ℃厌氧条件下培养24 h,进行重复传代,分别测定第1代、第3代、第5代、第7代、第9代和第10代时产L-乳酸能力(三次平行试验),对比传代不同次数对产L-乳酸能力的影响。
1.2.8 鼠李糖乳杆菌AK-0779的发酵培养基单因素试验
筛选菌株AK-0779在发酵过程中被自身产物所抑制,因此,在单因素实验时,向YE培养基中添加中和剂碳酸钙。在此条件下,发酵周期延长至72 h达到稳定状态,因此下述单因素实验发酵周期选择72 h。
1.2.8.1 不同碳源添加量对菌株AK-0779产L-乳酸的影响
在接种量为4%的YE培养基基础上,将葡萄糖作为发酵的碳源,以浓度分别为2%、5%、10%和15%添加量加入到容量为30 mL的三角瓶中,于37 ℃、90 r/min的恒温摇床中培养,发酵周期为72 h,考察不同葡萄糖添加量对L-乳酸产量及残糖含量的影响。
1.2.8.2 不同碳酸钙添加量对菌株AK-0779菌株产L-乳酸的影响
在接种量为4%的YE培养基基础上,葡萄糖添加量为10%,将碳酸钙作为发酵产酸过程中的中和剂,分别以1%、2%、3%、4%、5%的添加量添加到容量为30 mL的三角瓶中,于37 ℃、90 r/min的恒温摇床中培养,发酵周期为72 h,考察不同碳酸钙添加量对L-乳酸产量的影响。
1.2.8.3 不同玉米酒糟与酵母粉复配比对菌株AK-0779产L-乳酸的影响
根据前期预实验结果,酵母粉作为发酵的单一氮源时,最佳添加量为2.5%;当玉米酒糟充当发酵的单一氮源时,其L-乳酸产量极低,没有实际意义。为降低发酵的氮源成本,同时能将玉米酒糟中的营养成分资源化利用,本研究将玉米酒糟和酵母粉进行复配,以不同复配比添加进一步对培养条件优化。在接种量为4%的YE培养基基础上,总的氮源添加量为2.5%,玉米酒糟与酵母粉复配比设为3:1、2:1、1:1、1:2、1:3,分别添加到容量为30 mL的三角瓶中,于37 ℃、90 r/min的恒温摇床中培养,发酵周期为72 h,考察不同氮源复配比对L-乳酸产量的影响。
1.2.9 Box-Behnken响应面法优化试验
根据单因素实验结果,以最大程度降低发酵产L-乳酸的成本为出发点,特别是发酵氮源成本。在YE培养基基础上,对葡萄糖添加量、玉米酒糟添加量及酵母粉添加量进行三因素三水平的Box-Behnken试验设计,以L-乳酸产量为响应值,优化鼠李糖乳杆菌AK-0779最佳发酵培养条件。各因素水平及编码如表1所示。
表 1 响应面试验因素水平设计Table 1. Response surface test factor level design因素 水平 −1 0 1 葡萄糖(%) 5 10 15 酵母粉(%) 1.25 1.67 1.88 玉米酒糟(%) 0.63 0.83 1.25 1.2.10 L-乳酸的测定
L-乳酸测定:L/D试剂盒。
(2) 1.2.11 残糖含量测定
3,5-二硝基水杨酸法[27]。
1.3 数据处理
全文试验数据通过Excel预处理后,用统计分析软件GraphPad prism 8.0.1进行显著性分析。P<0.05表示有显著差异,P<0.0001表示有极显著差异。
2. 结果与分析
2.1 初始菌株的酒精胁迫试验及发酵产酸能力
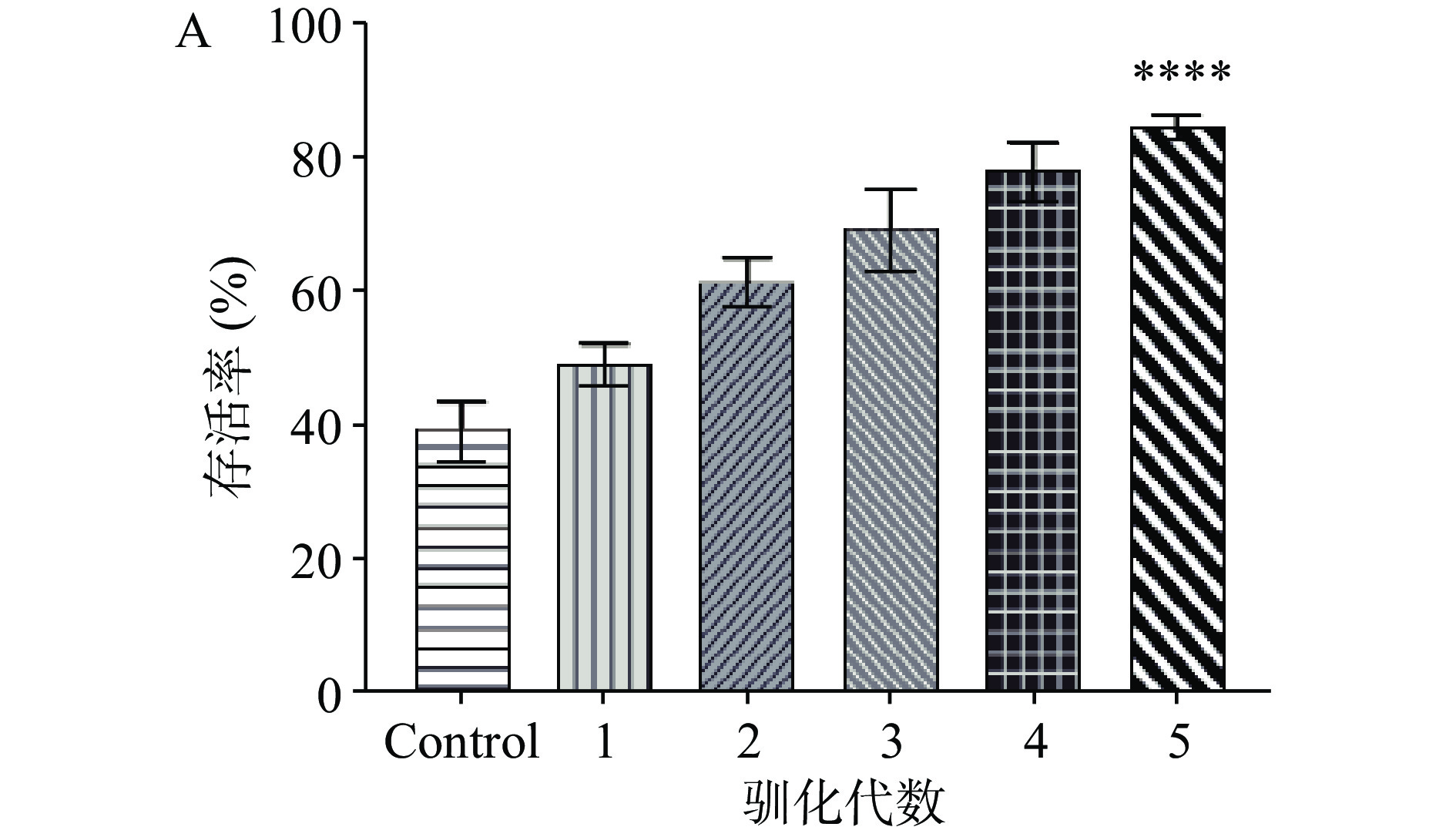
初始菌株在不同酒精体积分数下存活情况如图1A所示,在含5%酒精胁迫条件下,各菌株存活率均在40%~60%之间;当酒精体积分数增至10%时,所有菌株生长均受到明显抑制,菌株6133、6166、AK-0779及乳杆菌1受酒精抑制相对较小,菌株6141存活率明显降低。如图1B所示,初始菌株在初始葡萄糖浓度为20 g/L的条件下,相比于其他菌株初始乳酸量,菌株AK-0779发酵后乳酸增量最高,增量达到约11.02 g/L。在本研究中,各初始菌株在酒精胁迫下的存活率作为筛选的一个辅助条件,主要的筛选指标为乳酸产量。因此,选择菌株AK-0779作为筛选菌株,进行下一步驯化试验,以此来提高菌株AK-0779在精胁迫下的存活率。
![]() 图 1 初始菌株酒精胁迫及发酵产酸试验注:A.酒精胁迫;B.发酵乳酸增量;同一指标不同字母代表差异显著,P<0.05;相同字母代表差异不显著,P>0.05。Figure 1. Experiment on alcohol stress and fermentation acid production of the initial strain
图 1 初始菌株酒精胁迫及发酵产酸试验注:A.酒精胁迫;B.发酵乳酸增量;同一指标不同字母代表差异显著,P<0.05;相同字母代表差异不显著,P>0.05。Figure 1. Experiment on alcohol stress and fermentation acid production of the initial strain2.2 筛选菌株的驯化试验
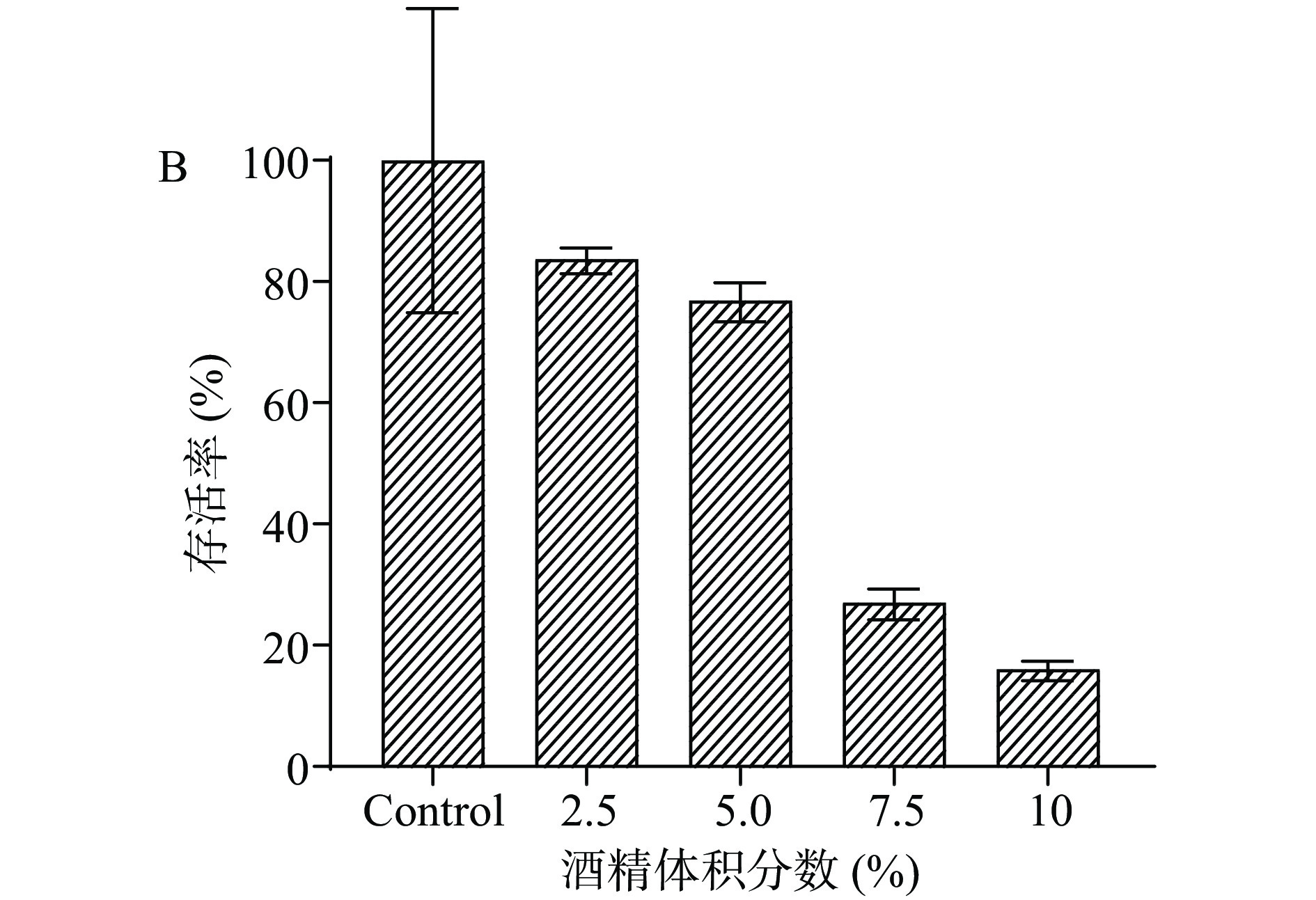
对筛选菌株AK-0779进行5%的酒精胁迫驯化。如图2A所示,当菌株驯化到第5代时存活率较未驯化时显著提高。将驯化5代后的筛选菌株在不同酒精体积分数下生长时,菌株存活结果如图2B所示,5%酒精条件下,其存活率为76.50%,与未驯化时的存活率42%相比,提高了34.50%,说明其在酒精胁迫条件下敏感性降低,能更好的在胁迫条件下生长。
![]() 图 2 菌株AK-0779的驯化及驯化后的酒精胁迫试验注:A.驯化情况;B.驯化后的酒精耐受情况;****表示处理组(驯化5代)与对照组(Control)差异极显著P<0.0001。Figure 2. Domestication of strain AK-0779 and alcohol stress test after training
图 2 菌株AK-0779的驯化及驯化后的酒精胁迫试验注:A.驯化情况;B.驯化后的酒精耐受情况;****表示处理组(驯化5代)与对照组(Control)差异极显著P<0.0001。Figure 2. Domestication of strain AK-0779 and alcohol stress test after training2.3 筛选菌株的鉴定
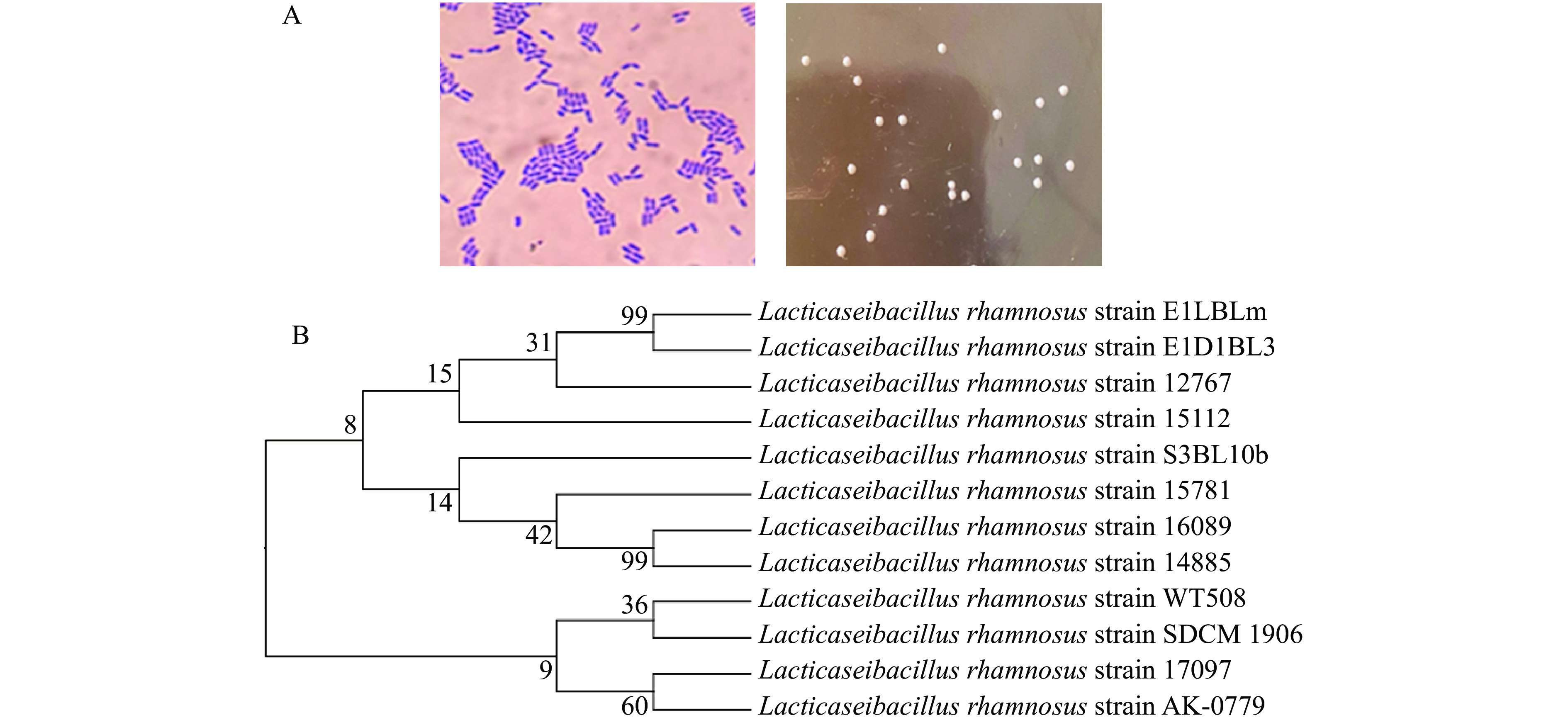
对驯化后的菌株AK-0779进行稀释涂布,在MRS固体培养基上培养48 h,经革兰氏染色,呈革兰氏阳性,形态呈短杆状(图3A),观察到菌株在平板上呈乳白色,圆形,表面光滑细腻的菌落形态(图3B)。经DNA序列测定,测定结果在NCBI官网进行序列比对,结果发现该菌株与鼠李糖乳杆菌显示出最高的分子系统学上的亲缘关系,同源性高达99.65%,将其命名为鼠李糖乳杆菌AK-0779,构建的系统发育树如(图3C)。
![]() 图 3 鼠李糖乳杆菌AK-0779的形态及系统发育树注:(A) 菌体形态和菌落形态;(B) 系统发育树。Figure 3. 16S rDNA sequence identification of strain AK-0779
图 3 鼠李糖乳杆菌AK-0779的形态及系统发育树注:(A) 菌体形态和菌落形态;(B) 系统发育树。Figure 3. 16S rDNA sequence identification of strain AK-07792.4 鼠李糖乳杆菌AK-0779发酵特性及产酸稳定性
在本实验室前期的研究中,发现菌株AK-0779主要产L-乳酸。如图4A和图4B所示,鼠李糖乳杆菌AK-0779在发酵12~48 h内能较好的维持活性,产L-乳酸13.46~13.75 g/L;当发酵60 h时,L-乳酸的产量有所下降,这可能是菌株在产酸过程会被其本身产物抑制。传代10次,结果如图4C所示,对比不同传代次数,鼠李糖乳杆菌AK-0779产L-乳酸量无明显差异,产酸稳定。结果说明鼠李糖乳杆菌AK-0779在发酵和产酸过程中具有良好的稳定性。
![]() 图 4 鼠李糖乳杆菌AK-0779的发酵性及其发酵产L-乳酸稳定性注:(A)活菌存活数;(B)发酵产酸性;(C)发酵产酸稳定性。Figure 4. Fermentability of L. rhamnosus AK-0779 and stability of L-lactic acid
图 4 鼠李糖乳杆菌AK-0779的发酵性及其发酵产L-乳酸稳定性注:(A)活菌存活数;(B)发酵产酸性;(C)发酵产酸稳定性。Figure 4. Fermentability of L. rhamnosus AK-0779 and stability of L-lactic acid2.5 鼠李糖乳杆菌AK-0779的发酵培养基单因素实验
2.5.1 葡萄糖添加量对鼠李糖乳杆菌AK-0779产L-乳酸影响
碳源构成了微生物生长和代谢的物质基础[28],同时也是发酵产品的起始原料,本研究以葡萄糖作为微生物发酵的碳源,考察不同葡萄糖添加量对菌株发酵产L-乳酸的影响。如图5所示,在YE发酵培养基其他条件不变的情况下,随着葡萄糖添加量的增加,鼠李糖乳杆菌AK-0779产L-乳酸量也逐渐增加,残糖含量也相应的增加。当葡萄糖添加量为10%时,L-乳酸产量最高,达到62.35 g/L。当添加量达到15%时,L-乳酸产量略有下降,这有可能在高糖浓度培养基中,形成了高渗透压环境,抑制了菌体的生长和代谢[29],或部分乳酸发生了转化,形成了乳酸盐,所以L-乳酸的产量降低。综合考虑,选择初始葡萄糖添加量为10%为宜。
![]() 图 5 葡萄糖添加量的L-乳酸产量及残糖含量注:*表示与对照组(葡萄糖添加量2%)相比,同一指标处理组数据差异显著,P<0.05。Figure 5. L-lactic acid yield and residual sugar content with different glucose addition
图 5 葡萄糖添加量的L-乳酸产量及残糖含量注:*表示与对照组(葡萄糖添加量2%)相比,同一指标处理组数据差异显著,P<0.05。Figure 5. L-lactic acid yield and residual sugar content with different glucose addition2.5.2 碳酸钙添加量对鼠李糖乳杆菌AK-0779产L-乳酸影响
乳杆菌的产酸能力特别强,其生长会受到代谢产物的抑制——乳酸[30]。因此,加入碳酸钙来中和发酵产生的乳酸,有助于保持菌株生长的最佳pH,以便它们能够充分利用营养底物,最大限度地将葡萄糖转化为乳酸。但碳酸钙添加量也会对菌株的生长有影响。添加量太少,中和不完全,达不到解除产物抑制问题,产酸能力依旧被限制;若添加量过多,会形成沉淀,菌株会被碳酸钙包埋,抑制发酵[31]。因此,确定合适的碳酸钙添加量也十分重要。由图6可以看出,L-乳酸的产量随着碳酸钙添加量增大而增加,3%时产量最高为82.36 g/L。当碳酸钙的添加量超过3%时,培养基在发酵过程中会逐渐凝固,产酸量会减少。因此,碳酸钙的最佳添加量为3%。
![]() 图 6 碳酸钙添加量对L-乳酸产量的影响注:*表示处理组与对照组(碳酸钙添加量1%)差异显著P<0.05;**表示差异极显著P<0.01。Figure 6. Effects of different CaCO3 addition on L-lactic acid production by L. rhamnosus AK-0779
图 6 碳酸钙添加量对L-乳酸产量的影响注:*表示处理组与对照组(碳酸钙添加量1%)差异显著P<0.05;**表示差异极显著P<0.01。Figure 6. Effects of different CaCO3 addition on L-lactic acid production by L. rhamnosus AK-07792.5.3 氮源复配比对鼠李糖乳杆菌AK-0779产L-乳酸影响
氮源是微生物生长必不可少的生长因子[32-33]。酵母粉是当前微生物发酵产L-乳酸最常用的氮源[34],然而较高的价格增加了L-乳酸生产成本。因此,采用廉价的培养基原料获得高产量L-乳酸是市场需求[35]。本研究采取玉米酒糟代替部分酵母粉作为氮源发酵生产L-乳酸,结果如图7所示,当玉米酒糟与酵母粉的复配比为1:2时,L-乳酸产量最高,为78.89 g/L,说明玉米酒糟代替部分酵母粉作为生产L-乳酸的氮源是可行的。所以选择玉米酒糟与酵母粉的复配比为1:2较为适宜。
![]() 图 7 氮源复配比对L-乳酸产量的影响注:**表示处理组与对照组(玉米酒糟:酵母粉=3:1)差异显著,P<0.05;****表示差异极显著,P<0.0001。Figure 7. Effects of different nitrogen source and compound ratio on L-lactic acid production by L. rhamnosus AK-0779
图 7 氮源复配比对L-乳酸产量的影响注:**表示处理组与对照组(玉米酒糟:酵母粉=3:1)差异显著,P<0.05;****表示差异极显著,P<0.0001。Figure 7. Effects of different nitrogen source and compound ratio on L-lactic acid production by L. rhamnosus AK-07792.6 响应面设计优化发酵培养基结果
2.6.1 响应面试验结果及方差分析
将影响L-乳酸产量的葡萄糖、玉米酒糟和酵母粉作为响应面试验设计的三个因素。响应面优化结果见表2,方差分析结果见表3。
表 2 响应面试验结果Table 2. Response surface test results实验号 因素 L-乳酸(g/L) A葡萄糖 B玉米酒糟 C酵母粉 1 0 0 0 79.22 2 1 0 −1 75.14 3 1 0 1 69.16 4 0 0 0 79.63 5 1 1 0 73.14 6 0 1 −1 74.17 7 0 −1 −1 72.74 8 −1 1 0 68.52 9 0 1 1 73.56 10 −1 0 1 71.46 11 0 −1 1 72.60 12 1 −1 0 71.20 13 −1 −1 0 66.68 14 0 0 0 79.37 15 0 0 0 79.59 16 0 0 0 78.95 17 −1 0 −1 65.52 表 3 响应面模型方差分析Table 3. Anova of response surface model来源 平方和 自由度 均方 F值 P值 显著性 模型 386.98 9 43.00 297.08 <0.0001 *** A 33.90 1 33.90 234.22 <0.0001 *** B 4.74 1 4.74 32.77 0.0007 ** C 12.34 1 12.34 85.27 <0.0001 *** AB 0.0025 1 0.0025 0.0172 0.8993 / AC 35.49 1 35.49 245.19 <0.0001 *** BC 0.0575 1 0.0575 0.3971 0.5486 / A2 183.86 1 183.86 1270.35 <0.0001 *** B2 56.40 1 56.40 389.69 <0.0001 *** C2 43.81 1 43.81 302.66 <0.0001 *** 残差 1.01 7 0.1447 失拟项 0.7205 3 0.2402 3.29 0.1403 / 纯误差 0.2925 4 0.0731 总和 387.99 16 R2 0.9974 R2Adj 0.9940 R2 Pre 0.9691 C.V.% 0.5155 1 注:P<0.05,差异显著,标记为*;P<0.01,差异非常显著,标记为**;P<0.0001,差异极显著,标记为***;P>0.05,差异不显著,标记为/。 利用Design-Expert 8.0软件对表2数据进行多元回归分析,得出多元回归方程:Y=80.15+2.06A+12.34B−0.0985C+0.025AB−2.98AC−0.1199BC−6.61A2−3.66B²−3.23C2。从表3中可以看出,回归模型极显著(P<0.0001),失拟项P值为0.1403>0.05,而模型的决定系数R2=0.9974,表明模型被成功拟合,试验值与预测值一致;而模型的拟合调整决定系数R2Adj=0.9940,表明99.40%的变异性可由模型解释,因此认为该模型可以用于预测不同水平的L-乳酸产量。3个因素都显著影响L-乳酸的产量,且对L-乳酸产量的影响大小依次为葡萄糖添加量>酵母粉添加量>玉米酒糟添加量。
2.6.2 各因素交互作用结果分析
不同因素交互作用对L-乳酸产量影响的响应面如图8所示。在图8中的响应曲面都有最高的峰值,表明所选因素的范围是可行的[36]。响应曲面越陡峭,说明该因素对L-乳酸的影响越大[37]。图8A可以看出,当酵母粉添加量不变时,随着葡萄糖添加量和玉米酒糟添加量的增大,L-乳酸产量先增大后减小。同样,由图8B、图8C可以看出,当玉米酒糟和葡萄糖添加量不变时,L-乳酸的产量趋于先增加后减少。葡萄糖添加量对产L-乳酸的响应曲面最陡峭,其次为酵母粉添加量,玉米酒糟添加量响应曲面最平缓,这与方差分析中的结果一致。
![]() 图 8 各因素交互影响的响应面图注:(A)葡萄糖和玉米酒糟;(B)葡萄糖和酵母粉;(C)玉米酒糟和酵母粉。Figure 8. Response surface diagram of interaction of various factors
图 8 各因素交互影响的响应面图注:(A)葡萄糖和玉米酒糟;(B)葡萄糖和酵母粉;(C)玉米酒糟和酵母粉。Figure 8. Response surface diagram of interaction of various factors2.6.3 验证试验结果
由回归方程可知,以玉米酒糟为基质鼠李糖乳杆菌AK-0779发酵培养基最佳工艺配方为葡萄糖添加量9.80%、玉米酒糟添加量0.98%、酵母粉添加量1.72%,在该条件下,L-乳酸的预期产量为79.25 g/L。进行3次验证重复试验,得到L-乳酸的产量分别78.88、78.89、78.97 g/L,平均值是78.91 g/L,相对误差为0.43%。
2.7 优化后氮源与酵母粉作为氮源产L-乳酸比较
通常情况下,选用酵母粉充当氮源会增加L-乳酸生产成本,低廉的农业或工业生产废弃物越来越受到L-乳酸生产企业的欢迎。本研究选用燃料酒精生产的废弃物玉米酒糟作为L-乳酸发酵生产的氮源,优化后氮源与酵母粉作为氮源产L-乳酸比较,结果如图9所示,酵母粉完成充当氮源时,L-乳酸产量为82.36 g/L,糖酸转换率为81.81%;玉米酒糟代替部分酵母粉充当氮源优化后L-乳酸产量78.91 g/L,糖酸转换率为80.52%,二者产量无明显差异。据市场调查,酵母粉的价格一般为每吨7万元,生产1吨L-乳酸如果完全用酵母粉作为氮源则需要2.12万元的氮源成本,而玉米酒糟的价格大约为每吨0.15万元,按照本研究的优化结果用玉米酒糟替代部分氮源生产1吨L-乳酸则需要1.54万元氮源成本,所以较完全用酵母粉作为氮源节省了0.58万元成本。说明玉米酒糟可以有效替代部分酵母粉作为微生物发酵氮源,达到了节约L-乳酸生产成本的目的。
![]() 图 9 不同条件优化后的鼠李糖乳杆菌AK-0779产酸情况及糖酸转换率Figure 9. Acid production and sugar acid conversion rate of L. rhamnosus AK-0779 optimized under different conditions
图 9 不同条件优化后的鼠李糖乳杆菌AK-0779产酸情况及糖酸转换率Figure 9. Acid production and sugar acid conversion rate of L. rhamnosus AK-0779 optimized under different conditions3. 讨论与结论
利用微生物发酵产L-乳酸的研究已成为热点,近几年,科研人员开始注重L-乳酸发酵基质的研究,多用廉价的农业废弃物替代生产L-乳酸的主要基质,如李慧芬等[38]用植物乳杆菌发酵豆粕产L-乳酸的研究;孙丽慧等[39]在高产L-乳酸的发酵培养基优化研究中,采用15 g/L棉籽饼粉和10 g/L的酵母粉为复合氮源,结果表明,廉价的棉籽饼粉可以替代部分酵母粉,还能达到高产L-乳酸的效果。王玉华等[14]用玉米浆替代部分酵母粉作为发酵乳酸的氮源,大大降低了L-乳酸的生产成本。使用玉米酒糟代替部分酵母粉为氮源的乳杆菌发酵尚未见到报道。本研究将玉米酒糟替代部分酵母粉,发现玉米酒糟能有效替代部分酵母粉。Kwon等[40]在优化产L-乳酸的发酵培养基中用19.3 g/L大豆粉替代15 g/L的酵母粉,同样获得了理想的结果,但需要在培养基中添加额外的维生素。Nancib等[41]也发现利用廉价氮源替代部分酵母粉,可以提高L-乳酸的产量,同样需要添加额外的维生素,这样反而没有达到降低发酵成本的目的。我国一直享有农业大国的称号,玉米资源十分丰富,特别是东三省,将玉米酒糟替代部分酵母粉作为微生物发酵氮源,有效节约L-乳酸生产成本。
本研究以燃料酒精生产产生的副产物玉米酒糟为基质,发酵生产L-乳酸,响应面法优化L-乳酸发酵培养基,得到如下结论: 经过筛选、驯化获得一株耐酒精高产L-乳酸的菌株鼠李糖乳杆菌AK-0779;利用筛选、驯化后的鼠李糖乳杆菌AK-0779发酵生产L-乳酸,通过响应面法确定最佳发酵培养基配方为:葡萄糖添加量为9.80%、玉米酒糟添加量为0.98%、酵母粉添加量为1.72%,在该条件下L-乳酸产量预测值为79.25 g/L。对比以玉米酒糟充当部分氮源和酵母粉完全充当氮源产L-乳酸的量,结果表明:玉米酒糟充当部分氮源L-乳酸产量为78.91 g/L,酵母粉完全充当氮源L-乳酸产量为82.36 g/L,产量无显著差异,说明玉米酒糟能有效代替部分酵母粉作为发酵培养基的氮源,降低L-乳酸生产成本。同时,也推动了我国玉米深加工企业的发展。
-
![]()
图 1 初始菌株酒精胁迫及发酵产酸试验
注:A.酒精胁迫;B.发酵乳酸增量;同一指标不同字母代表差异显著,P<0.05;相同字母代表差异不显著,P>0.05。
Figure 1. Experiment on alcohol stress and fermentation acid production of the initial strain
![]()
图 2 菌株AK-0779的驯化及驯化后的酒精胁迫试验
注:A.驯化情况;B.驯化后的酒精耐受情况;****表示处理组(驯化5代)与对照组(Control)差异极显著P<0.0001。
Figure 2. Domestication of strain AK-0779 and alcohol stress test after training
![]()
图 3 鼠李糖乳杆菌AK-0779的形态及系统发育树
注:(A) 菌体形态和菌落形态;(B) 系统发育树。
Figure 3. 16S rDNA sequence identification of strain AK-0779
![]()
图 4 鼠李糖乳杆菌AK-0779的发酵性及其发酵产L-乳酸稳定性
注:(A)活菌存活数;(B)发酵产酸性;(C)发酵产酸稳定性。
Figure 4. Fermentability of L. rhamnosus AK-0779 and stability of L-lactic acid
![]()
图 5 葡萄糖添加量的L-乳酸产量及残糖含量
注:*表示与对照组(葡萄糖添加量2%)相比,同一指标处理组数据差异显著,P<0.05。
Figure 5. L-lactic acid yield and residual sugar content with different glucose addition
![]()
图 6 碳酸钙添加量对L-乳酸产量的影响
注:*表示处理组与对照组(碳酸钙添加量1%)差异显著P<0.05;**表示差异极显著P<0.01。
Figure 6. Effects of different CaCO3 addition on L-lactic acid production by L. rhamnosus AK-0779
![]()
图 7 氮源复配比对L-乳酸产量的影响
注:**表示处理组与对照组(玉米酒糟:酵母粉=3:1)差异显著,P<0.05;****表示差异极显著,P<0.0001。
Figure 7. Effects of different nitrogen source and compound ratio on L-lactic acid production by L. rhamnosus AK-0779
![]()
图 8 各因素交互影响的响应面图
注:(A)葡萄糖和玉米酒糟;(B)葡萄糖和酵母粉;(C)玉米酒糟和酵母粉。
Figure 8. Response surface diagram of interaction of various factors
![]()
图 9 不同条件优化后的鼠李糖乳杆菌AK-0779产酸情况及糖酸转换率
Figure 9. Acid production and sugar acid conversion rate of L. rhamnosus AK-0779 optimized under different conditions
表 1 响应面试验因素水平设计
Table 1 Response surface test factor level design
因素 水平 −1 0 1 葡萄糖(%) 5 10 15 酵母粉(%) 1.25 1.67 1.88 玉米酒糟(%) 0.63 0.83 1.25  下载: 导出CSV
下载: 导出CSV
表 2 响应面试验结果
Table 2 Response surface test results
实验号 因素 L-乳酸(g/L) A葡萄糖 B玉米酒糟 C酵母粉 1 0 0 0 79.22 2 1 0 −1 75.14 3 1 0 1 69.16 4 0 0 0 79.63 5 1 1 0 73.14 6 0 1 −1 74.17 7 0 −1 −1 72.74 8 −1 1 0 68.52 9 0 1 1 73.56 10 −1 0 1 71.46 11 0 −1 1 72.60 12 1 −1 0 71.20 13 −1 −1 0 66.68 14 0 0 0 79.37 15 0 0 0 79.59 16 0 0 0 78.95 17 −1 0 −1 65.52
下载: 导出CSV
表 3 响应面模型方差分析
Table 3 Anova of response surface model
来源 平方和 自由度 均方 F值 P值 显著性 模型 386.98 9 43.00 297.08 <0.0001 *** A 33.90 1 33.90 234.22 <0.0001 *** B 4.74 1 4.74 32.77 0.0007 ** C 12.34 1 12.34 85.27 <0.0001 *** AB 0.0025 1 0.0025 0.0172 0.8993 / AC 35.49 1 35.49 245.19 <0.0001 *** BC 0.0575 1 0.0575 0.3971 0.5486 / A2 183.86 1 183.86 1270.35 <0.0001 *** B2 56.40 1 56.40 389.69 <0.0001 *** C2 43.81 1 43.81 302.66 <0.0001 *** 残差 1.01 7 0.1447 失拟项 0.7205 3 0.2402 3.29 0.1403 / 纯误差 0.2925 4 0.0731 总和 387.99 16 R2 0.9974 R2Adj 0.9940 R2 Pre 0.9691 C.V.% 0.5155 1 注:P<0.05,差异显著,标记为*;P<0.01,差异非常显著,标记为**;P<0.0001,差异极显著,标记为***;P>0.05,差异不显著,标记为/。
下载: 导出CSV
-
[1] 孙华. 以玉米为原料发酵生产L-乳酸工艺条件优化[J]. 中国新技术新产品,2019(22):51−52. [SUN Hua. Optimization of fermentation conditions for L-lactic acid production from maize[J]. New Technology and New Products in China,2019(22):51−52. doi: 10.3969/j.issn.1673-9957.2019.22.030 [2] WANG Y, CHEN C, CAI D, et al. The optimization of L-lactic acid production from sweet sorghum juice by mixed fermentation of Bacillus coagulans and Lactobacillus rhamnosus under unsterile conditions[J]. Bioresource Technology,2016,218:1098−1105. doi: 10.1016/j.biortech.2016.07.069
[3] KARIN H, BRBEL H H. Factors affecting the fermentative lactic acid production from renewable resources 1[J]. Enzyme and Microbial Technology,2000,26(2-4):87−107. doi: 10.1016/S0141-0229(99)00155-6
[4] 吴晖, 李明阳, 刘冬梅, 等. 鼠李糖乳杆菌发酵生产L-乳酸培养基的优化[J]. 食品工业科技,2008(4):81−84. [WU Hui, LI Mingyang, LIU Dongmei, et al. Optimization of L-lactic acid fermentation medium by Lactobacillus rhamnosus[J]. Science and Technology of Food Industry,2008(4):81−84. doi: 10.13386/j.issn1002-0306.2008.04.011 [5] 张为巍, 李元媛, 周铭锋, 等. 产L-乳酸的鼠李糖乳杆菌发酵培养基的优化[J]. 广州化工,2012,40(7):79−82. [ZHANG Weiwei, LI Yuanyuan, ZHOU Mingfeng, et al. Optimization of fermentation medium for L-lactic acid producing Lactobacillus rhamnosus[J]. Guangzhou Chemical Industry Co., Ltd.,2012,40(7):79−82. doi: 10.3969/j.issn.1001-9677.2012.07.028 [6] 李海洋, 韩军岐, 徐长亮. 鼠李糖乳杆菌乳酸发酵特性及碳氮源研究[J]. 乳业科学与技术,2012,35(2):17−20. [LI Haiyang, HAN Junqi, XU Changliang. Study on lactic acid fermentation characteristics and carbon and nitrogen sources of Lactobacillus rhamnosus[J]. Dairy Science and Technology,2012,35(2):17−20. doi: 10.3969/j.issn.1671-5187.2012.02.005 [7] 胡嘉欢, 张谷涵, 付永前, 等. 米根霉一步发酵法高效积累L-乳酸的策略[J]. 过程工程学报,2017,17(2):375−381. [HU Jiahuan, ZHANG Guhan, FU Yongqian, et al. The strategy of efficient accumulation of L-lactic acid by Rhizopus oryzae in one-step fermentation[J]. Journal of Process Engineering,2017,17(2):375−381. doi: 10.12034/j.issn.1009-606X.216303 [8] BERNARDO M P, COELHO L F, SASS D C, et al. L-(+)-lactic acid production by Lactobacillus rhamnosus B103 from dairy industry waste[J]. Brazilian Journal of Microbiology,2016,47(3):640−646. doi: 10.1016/j.bjm.2015.12.001
[9] ZHENG Jin, GAO Ming, WANG Qunhui, et al. Enhancement of l-lactic acid production via synergism in open co-fermentation of sophora flavescens residues and food waste[J]. Bioresource Technology,2017,225:159−164. doi: 10.1016/j.biortech.2016.11.055
[10] 赵鹏, 黄霞. 利用可再生资源及有机废物发酵生产乳酸的研究进展[J]. 食品与发酵工业,2001(4):60−65. [ZHAO Peng, HUANG Xia. Research progress in production of lactic acid by fermentation from renewable resources and organic wastes[J]. Food and Fermentation Industries,2001(4):60−65. doi: 10.3321/j.issn:0253-990X.2001.04.013 [11] 王勇. 以廉价生物质生产L-乳酸新方法研究[D]. 北京: 北京化工大学, 2017 WANG Yong. Study on a new method of producing L-lactic acid from cheap biomass [D]. Beijing: Beijing University of Chemical Technology, 2017.
[12] 于雷, 雷霆, 裴晓林, 等. L-乳酸发酵培养基中氮源的优化[J]. 食品科技,2007(6):49−53. [YU Lei, LEI Ting, PEI Xiaolin, et al. Optimization of nitrogen source in L-lactic acid fermentation medium[J]. Food Technology,2007(6):49−53. doi: 10.3969/j.issn.1005-9989.2007.06.014 [13] NGUYEN C M, KIM J S, HWANG H J, et al. Production of l-lactic acid from a green microalga, Hydrodictyon reticulum, by Lactobacillus paracasei LA104 isolated from the traditional Korean food, makgeolli.[J]. Bioresource Technology,2012,110:1302−1305.
[14] 王玉华, 陈萍, 朴春红, 等. 基因组改组鼠李糖乳杆菌生产L-乳酸发酵培养基的优化[J]. 食品科学,2009,30(21):316−319. [WANG Yuhua, CHEN Ping, PIAO Chunhong, et al. Optimization of L-lactic acid fermentation medium produced by genome shuffled Lactobacillus rhamnosus[J]. Food Science,2009,30(21):316−319. [15] 梁建光. 综合评价玉米酒糟及其可溶物[J]. 中国奶牛,2007(S1):64−66. [LIANG Jianguang. Comprehensive evaluation of corn distiller's grains and their soluble substances[J]. Chinese Dairy Cows,2007(S1):64−66. [16] SUPARNA S, NARAYAN B S, HEMEN S, et al. Utilization of distillers dried grains with solubles as a cheaper substrate for sophorolipid production by Rhodotorula babjevae YS3[J]. Journal of Environmental Chemical Engineering,2021,9(4):105494. doi: 10.1016/j.jece.2021.105494
[17] 石孔泉. 基于玉米浓醪酒糟基质的L-乳酸菌株的选育与发酵研究[D]. 无锡: 江南大学, 2006 SHI Kongquan. Breeding and fermentation of L-lactic acid strains based on the substrate of corn high ferment [D]. Wuxi: Jiangnan University, 2006.
[18] 李凯年, 逯德山. 在猪饲料中利用干玉米酒糟残液研究进展[J]. 猪业科学,2009,26(4):62−65. [LI Kainian, LU Deshan. Research progress on utilization of dry corn distiller's grains residue in pig feed[J]. Pig Science,2009,26(4):62−65. doi: 10.3969/j.issn.1673-5358.2009.04.022 [19] 杨一芃. 玉米酒糟的营养价值及猪饲料配比方法[J]. 山西农业科学,2010,38(4):95−96. [YANG Yipeng. Nutritional value of corn distiller's grains and proportioning method of pig feed[J]. Shanxi Agricultural Science,2010,38(4):95−96. doi: 10.3969/j.issn.1002-2481.2010.04.33 [20] 吴天祥, 杨海龙, 石贵阳, 等. 玉米酒糟乳酸菌发酵饲料培养基优化研究[J]. 饲料研究,2003(12):6−8. [WU Tianxiang, YANG Hailong, SHI Guiyang, et al. Study on optimization of feed medium for lactic acid bacteria fermentation of corn distiller's grains[J]. Feed Research,2003(12):6−8. doi: 10.3969/j.issn.1002-2813.2003.12.003 [21] JAE-CHEOL J, ZHIKAI Z, E U P, et al. Effects of feeding corn distillers dried grains with solubles diets without or with supplemental enzymes on growth performance of pigs: A meta-analysis[J]. Translational Animal Science,2021,5(2):1−15.
[22] 杨嘉伟. 玉米酒糟浸出脱脂研究[D]. 无锡: 江南大学, 2013 YANG Jiawei. Study on degreasing of corn distiller's grains [D]. Wuxi: Jiangnan University. 2013.
[23] 张拴力, 扈士海, 崔云, 等. 玉米酒糟为主要氮源的Nisin生产菌株的诱变选育[J]. 河北大学学报(自然科学版),2016,36(4):417−423. [ZHANG Shuanli, HU Shihai, CUI Yun, et al. Mutation breeding of nisin producing strain with corn distiller's grains as the main nitrogen source[J]. Journal of Hebei University (Natural Science Edition),2016,36(4):417−423. doi: 10.3969/j.issn.1000-1565.2016.04.014 [24] OTARI N, ARSLAN A, CARVALHO B, et al. Integrated Multi-Omics analysis of mechanisms underlying yeast ethanol tolerance[J]. Journal of Proteome Research,2021,20(8):3840−3852. doi: 10.1021/acs.jproteome.1c00139
[25] LI R, MIAO Y, YUAN S, et al. Integrated transcriptomic and proteomic analysis of the ethanol stress response in Saccharomyces cerevisiae Sc131[J]. Journal of Proteomics,2019,203:103377. doi: 10.1016/j.jprot.2019.103377
[26] 赵戎蓉, 贺娟. 耐60℃高温乳酸菌的驯化及鉴定[J]. 中国微生态学杂志,2011,23(4):4. [ZHAO Rongrong, HE Juan. Domestication and identification of 60 ℃ high temperature resistant lactic acid bacteria[J]. Chinese Journal of Microbiology,2011,23(4):4. [27] 赵凯, 许鹏举, 谷广烨. 3,5-二硝基水杨酸比色法测定还原糖含量的研究[J]. 食品科学,2008,29(8):3. [ZHAO Kai, XU Pengju, GU Guangye. Study on the determination of reducing sugar content by 3,5-Dinitrosalicylic acid colorimetry[J]. Food Science,2008,29(8):3. [28] 刘毓锋, 曾嘉锐, 黄文琪, 等. 外源碳源对葡萄酵素微生物生长代谢及生物活性的调节作用[J]. 食品工业科技,2020,41(8):104−110,116. [LIU Yufeng, ZENG Jiarui, HUANG Wenqi, et al. Regulation of exogenous carbon source on growth metabolism and biological activity of grape ferment microorganisms[J]. Science and Technology of Food Industry,2020,41(8):104−110,116. [29] 汤凤霞, 乔长晟, 蔡慧农, 等. L-乳酸米根霉发酵条件优化研究[J]. 集美大学学报(自然科学版),2007,12(2):118−123. [TANG Fengxia, QIAO Changsheng, CAI Huinong, et al. Optimization of fermentation conditions of Rhizopus oryzae L-lactic acid[J]. Journal of Jimei University (Natural Science Edition),2007,12(2):118−123. doi: 10.3969/j.issn.1007-7405.2007.02.005 [30] 周剑. L-乳酸生产菌株的诱变选育及发酵条件初步优化[D]. 南京: 南京工业大学, 2005 ZHOU Jian. Mutation breeding of L-lactic acid producing strain and preliminary optimization of fermentation conditions [D]. Nanjing: Nanjing University of Technology, 2005.
[31] 雷霆. 高产L-乳酸菌种的选育及培养基的优化[D]. 长春: 吉林大学, 2006 LEI Ting. Breeding of high-yield L-lactic acid bacteria and optimization of culture medium [D]. Changchun: Jilin University. 2006.
[32] 郭瑞, 李亚楠, 李怡阳, 等. 不同食品级氮源对发酵肉用乳酸菌生长的影响[J]. 肉类工业,2022(7):22−27. [GUO Rui, LI Yanan, LI Yiyang, et al. Effects of different food-grade nitrogen sources on the growth of lactic acid bacteria for meat fermentation[J]. Meat Industry,2022(7):22−27. doi: 10.3969/j.issn.1008-5467.2022.07.004 [33] LI H, LIN J, LIN J, et al. Effects of nitrogen sources and vitamins on production of L-lactic acid with high efficiency using Lactobacillus rhamnosus[J]. Food and Fermentation Industries,2007,33(2):22−26.
[34] ZHAO N, YUAN X L, CHEN J, et al. Effect of different carbon and nitrogen sources on mycelia growth of Antrodia cinnamomea[J]. Journal of West China Forestry Science, 2016.
[35] JUTURU V, JIN C W. Microbial production of lactic acid: The latest development[J]. Critical Reviews in Biotechnology,2015,36(6):967−977.
[36] LONG X, YAN Q, CAI L, et al. Box-Behnken design-based optimization for deproteinization of crude polysaccharides in Lycium barbarum berry residue using the Sevag method[J]. Heliyon,2020,6(5):e3888.
[37] 时菲菲, 陈毓, 周晨, 等. 响应面法优化党参多糖提取工艺及抗氧化活性研究[J]. 畜牧与兽医,2021,53(1):54−60. [SHI Feifei, CHEN Yu, ZHOU Chen, et al. Optimization of extraction process and antioxidant activity of Codonopsis pilosula polysaccharides by response surface methodology[J]. Animal Husbandry and Veterinary Medicine,2021,53(1):54−60. [38] 李慧芬, 马成, 张克顺. 植物乳杆菌发酵豆粕产L-乳酸的最佳条件优化[J]. 饲料工业,2015,36(5):32−36. [LI Huifen, MA Cheng, ZHANG Keshun. Optimization of optimal conditions for L-lactic acid production from soybean meal by Lactobacillus plantarum[J]. Feed Industry,2015,36(5):32−36. [39] 孙丽慧, 王云晓, 吕诗文, 等. 1株高产L-乳酸菌株的分离鉴定及其发酵培养基优化[J]. 食品科学,2018,39(6):135−140. [SUN Lihui, WANG Yunxiao, LÜ Shiwen, et al. Isolation and identification of a high-yield L-lactic acid strain and optimization of its fermentation medium[J]. Food Science,2018,39(6):135−140. doi: 10.7506/spkx1002-6630-201806022 [40] KWON, LEE, KEUN C Y, et al. Production of lactic acid by Lactobacillus rhamnosus with vitamin-supplemented soybean hydrolysate[J]. Enzyme & Microbial Technology,2000,26(2):209−215.
[41] NANCIB A, NANCIB N, MEZIANE-CHERIF D, et al. Joint effect of nitrogen sources and B vitamin supplementation of date juice on lactic acid production by Lactobacillus casei subsp. rhamnosus[J]. Bioresource Technology,2005,96(1):63−67. doi: 10.1016/j.biortech.2003.09.018
-
期刊类型引用(7)
1. 云丹卓玛,蔺辉星. 西藏牦牛肉制品中致病菌的检测与风险评估. 食品安全导刊. 2025(03): 137-139 .  百度学术
百度学术
2. 张二豪,落桑央吉,高潭,罗章. 西藏传统风干牦牛肉中微生物群落组成及安全评价. 食品与发酵工业. 2024(08): 182-188 . 百度学术
3. 王福清,张春颖,姚雪晴,许荔新. 发酵牦牛肉中的菌株分离鉴定及其应用研究. 中国调味品. 2024(11): 79-83 . 百度学术
4. 车丽娜,赵良忠,周晓洁. 基于高通量测序分析湘派卤牛肉冷藏过程中真菌多样性. 食品安全质量检测学报. 2023(04): 127-136 . 百度学术
5. 李彦,符慧靖,秦建鹏,路玉倩,梁丽雅,吴子健,马俪珍. 天然复合添加剂对卤牛肉贮藏品质及风味的影响. 中国食品学报. 2023(11): 231-245 . 百度学术
6. 李思雨,杜贺超,包佳亮,姚宏亮,蒋加进. 贵州腌鱼微生物多样性和益生特性分析. 农产品加工. 2023(24): 66-71 . 百度学术
7. 杨爱馥,姜红旭,孙赟,梁冰,崔博采,郑秋月,姚丽锋,姜丽. 基于高通量测序分析进口大麦真菌多样性. 食品科技. 2022(11): 297-302 . 百度学术
其他类型引用(3)

 下载:
下载:

















计量
- 文章访问数: 145
- HTML全文浏览量: 33
- PDF下载量: 13
- 被引次数: 10



