


Protective Effect of 6′′′-Feruloylspinosin on Aβ1-42-Induced SH-SY5Y Cells Injury
-
摘要: 目的:探讨酸枣仁黄酮类成分6′′′-阿魏酰斯皮诺素预处理对Aβ1-42诱导损伤SH-SY5Y细胞的保护作用。方法:利用6′′′-阿魏酰斯皮诺素(1、5、10、20和40 μmol/L)预处理SH-SY5Y细胞2 h,随后加入5 μmol/L的Aβ1-42共同孵育24 h,利用CCK-8法检测细胞活力;Calcein-AM/PI双染法观察细胞形态;试剂盒检测细胞内活性氧、丙二醛、线粒体膜电位水平及谷胱甘肽过氧化物酶活力;蛋白免疫印迹法检测细胞凋亡相关蛋白:Bcl-2关联死亡启动子重组蛋白、B细胞淋巴瘤/白血病-2蛋白的表达。结果:6′′′-阿魏酰斯皮诺素可以抑制Aβ1-42诱导的细胞凋亡,提高细胞活力(P<0.001);改善Aβ1-42诱导损伤细胞的氧化应激情况,降低细胞内活性氧(P<0.001)、丙二醛水平(P<0.01),提高谷胱甘肽过氧化物酶活力(P<0.001);改善Aβ1-42诱导损伤的细胞线粒体功能,上调细胞线粒体膜电位水平(P<0.001);增加抗凋亡蛋白B细胞淋巴瘤/白血病-2蛋白的表达(P<0.01),并降低促凋亡蛋白Bcl-2关联死亡启动子重组蛋白的表达(P<0.01)。结论:6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤的SH-SY5Y细胞具有保护作用,其机制与6′′′-阿魏酰斯皮诺素抗氧化活性及调节凋亡相关蛋白表达有关。
-
关键词:
- 酸枣仁 /
- 6′′′-阿魏酰斯皮诺素 /
- β淀粉样蛋白 /
- 抗氧化活性 /
- 线粒体
Abstract: Objective: To investigate the protective effects of 6′′′-feruloylspinosinon Aβ1-42-induced SH-SY5Y cells injury. Methods: SH-SY5Y cells were pretreated with 6′′′-feruloylspinosin (1、5、10、20 and 40 μmol/L) for 2 h and then co-treated with 5 μmol/L of Aβ1-42 for 24 h. Cell viability was measured by CCK-8 assay; cell morphology was observed by Calcein-AM/PI assay; intracellular reactive oxygen species, malonic dialdehyde level, Glutathione peroxidaseactivity and mitochondrial membrane potential level were detected by the kit; expression of apoptosis-related proteins: Bcl-xL/Bcl-2 associated death promoter, B-cell lymphoma/leukemia-2 were detected by Western Blotting. Results: 6′′′-feruloylspinosin could inhibit Aβ1-42-induced apoptosis and improve cell viability (P<0.001); reduce intracellular reactive oxygen species (P<0.001) and malondialdehyde (P<0.01) levels; increase glutathione peroxidase activity (P<0.001); upregulate cell mitochondrial membrane potential level (P<0.001); regulate apoptosis-related protein expression (P<0.01). Conclusion: The results validated that 6′′′-feruloylspinosin had a protective effect on SH-SY5Y cells with Aβ1-42-induced damage, and the mechanism was that 6′′′-feruloylspinosin could alleviate oxidative damage and regulate the expression of apoptosis related proteins.-
Keywords:
- Ziziphi Spinosae Semen /
- 6′′′-feruloylspinosin /
- amyloid β /
- antioxidant activity /
- mitochondria
-
酸枣仁为鼠李科植物酸枣(Ziziphus jujuba Mill.var. Spinosa (Bunge) Hu ex H. F. Chou)的干燥成熟种子,其作为卫生部颁布的第一批药食两用资源,具有温和、依赖性低、可长期服用等优点,是当前研究的热门品种。酸枣仁具有悠久的应用历史,是中医治疗失眠多梦的首选药物[1]。由于现代药理学研究的逐渐完善,酸枣仁的活性功能被进一步挖掘,现代研究证明酸枣仁具镇静催眠、抗焦虑、降血糖、降血脂、增强免疫等多种药理活性。黄酮类成分被认为是酸枣仁发挥药理活性的主要物质基础之一,目前,已经从酸枣仁中分离得到二十余种黄酮类单体,6′′′-阿魏酰斯皮诺素(6′′′-Feruloylspinosin,6-FS)是酸枣仁主要的黄酮类成分之一,不同产区酸枣仁中6′′′-阿魏酰斯皮诺素含量约为0.26~0.88 mg/g[2-3]。近年多项研究表明该黄酮在神经功能调节方面表现出显著的生物活性[4-5],但其具体作用机制尚待进一步揭示。
由于现代生活压力及全球人口老龄化的日益加剧,阿尔兹海默症(Alzheimer's disease, AD)等神经退行性疾病,已经成为人类面临的最大公共健康挑战之一。截至2015年,全世界4680万人患有阿尔茨海默症 (中国占有相当大的比重,约1/4)[6],预计2050年这一数字将达到1.52亿。AD的发生与β淀粉样蛋白(Amyloid β,Aβ)的沉积密切相关,Aβ在脑内沉积形成的不溶性斑块可诱导神经元线粒体损伤,导致氧自由基大量积聚,进一步攻击核酸、蛋白质等生物大分子物质,最终导致大量神经元凋亡,损害大脑功能[7]。传统天然药物富含多样生物活性化合物,可以多环节、多靶点发挥神经保护作用,并且部分传统天然药物具有药食同源性,安全性高,已成为调节神经功能最为有效的手段之一[8-9]。本研究利用Aβ1-42诱导SH-SY5Y细胞损伤,并给予酸枣仁黄酮类成分6′′′-阿魏酰斯皮诺素进行干预,从细胞活力、细胞氧化应激情况以及细胞线粒体状态等多角度探讨6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤的SH-SY5Y细胞的保护作用及相关机制,为酸枣仁的应用与防治AD药物的开发提供研究基础。
1. 材料与方法
1.1 材料与仪器
人神经母细胞瘤细胞株(SH-SY5Y) 南开大学惠赠;6′′′-阿魏酰斯皮诺素 成都德锐可生物科技有限公司(货号:drk1727,纯度:≥98%);Aβ1-42 上海吉尔生化有限公司(货号:SJ053487,纯度:≥95%);六氟异丙醇 北京天根生化科技有限公司;CCK-8细胞增殖检测试剂盒 Biosharp白鲨生物公司;活细胞/死细胞双染试剂盒(Calcein-AM/PI) 北京北奥莱博科技公司;B细胞淋巴瘤/白血病-2蛋白(B-cell lymphoma/leukemia-2, Bcl-2)、Bcl-2关联死亡启动子重组蛋白(Bcl-xL/Bcl-2 associated death promoter, Bad)、β-肌动蛋白(Beta Actin,β-Actin)抗体 abcam公司;BCA蛋白浓度测定试剂盒、蛋白提取试剂盒 北京索莱宝生物科技有限公司;活性氧(Reactive Oxygen Species, ROS)检测试剂盒、谷胱甘肽过氧化物酶(Glutathione peroxidase, GSH-Px)活力检测试剂盒、丙二醛(Malonic dialdehyde, MDA)检测试剂盒 南京建成生物工程研究所;线粒体膜电位(mitochondrial membrane potential, MMP)水平检测试剂盒 碧云天生物技术研究所。
BP211D电子分析天平 Sartorius AG;HNY2102恒温培养箱 天津市欧诺仪器仪表有限公司;TI-DH 600352倒置荧光显微镜 日本尼康;SpectraMax® M3多功能酶标仪 Molecular;PowerPac TMHC蛋白电泳仪 Bio-rad。
1.2 实验方法
1.2.1 细胞培养及细胞给药方式
SH-SY5Y细胞培养于含10%胎牛血清的培养基中,培养条件为37 ℃、5% CO2,细胞密度为70%~80%时进行传代。选取对数生长期细胞进行实验,细胞培养过夜后,给予含6′′′-阿魏酰斯皮诺素的培养基预处理2 h,再加入Aβ1-42寡聚体共同孵育24 h。
1.2.2 CCK-8法检测细胞活力
按照文献介绍[10]的方法制备Aβ1-42寡聚体,细胞接种于96孔板中。首先分析Aβ1-42对细胞活力的影响,培养过夜后,在含有Aβ1-42(0、0.1、0.5、1、5和10 μmol/L)的培养基中培养24 h,结束后吸弃培养基,每孔加入100 μL CCK-8工作液(10 μL CCK-8溶液+90 μL DMEM),孵育4 h后测定450 nm处吸光度值,根据吸光值读数选择Aβ1-42的给药剂量。其次,分析6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤的细胞活力的影响,细胞培养过夜后,首先利用含有不同浓度(1、5、10、20和40 μmol/L)6′′′-阿魏酰斯皮诺素的培养基预处理2 h后,加入Aβ1-42共同培养24 h。只给予Aβ1-42,未给予6′′′-阿魏酰斯皮诺素的细胞为模型组细胞。细胞给药培养结束后,按照上述方法测定不同处理组细胞吸光度值。其中,OD对照是指具有细胞、CCK-8 溶液而没有药物溶液的孔的吸光度读数;OD空白是指没有细胞存在,单独具有培养基和 CCK-8 溶液的孔的吸光度读数。
1.2.3 AM/PI染色法观察细胞形态
取对数生长期的SH-SY5Y细胞接种于12孔板中,分为以下5组:对照组、Aβ1-42模型组(5 μmol/L)、6′′′-阿魏酰斯皮诺素低剂量组(6′′′-阿魏酰斯皮诺素5 μmol/L+Aβ1-42 5 μmol/L)、6′′′-阿魏酰斯皮诺素中剂量组(6′′′-阿魏酰斯皮诺素10 μmol/L+Aβ1-42 5 μmol/L)及6′′′-阿魏酰斯皮诺素高剂量组(6′′′-阿魏酰斯皮诺素20 μmol/L+Aβ1-42 5 μmol/L)每组设置三个复孔。采用Calcein-AM/PI双染法观察细胞形态,细胞培养结束后,加入Calcein-AM/PI染色工作液避光孵育细胞(37 ℃,30 min)。使用荧光倒置显微镜在(490±10)nm激发波长下观察活细胞与死细胞,另用545 nm激发波长单独观察死细胞。
1.2.4 SH-SY5Y细胞内ROS水平测定
细胞培养及给药方式如1.2.3,培养结束后,加入DCFH-DA活性氧染色工作液孵育细胞(37 ℃,50 min),孵育结束后加入DMEM洗涤。用酶标仪测定激发波长502 nm,发射波长为530 nm处的荧光强度值并利用荧光倒置显微镜进行观察。
1.2.5 SH-SY5Y细胞内MDA水平、GSH-Px酶活力及线粒体膜电位水平测定
细胞培养及给药方式如1.2.3,培养结束后,依据试剂盒说明书介绍的方法收集及处理细胞,并测定细胞内MDA水平及GSH-Px酶活力。用JC-1法测定细胞线粒体膜电位,在细胞培养结束后加入JC-1染色工作液孵育细胞(37 ℃,30 min),用酶标仪测定激发波长490 nm,发射波长为520 nm及570 nm处的荧光强度值并利用荧光倒置显微镜进行观察。
1.2.6 Western blot分析
依据试剂盒说明书介绍的方法进行蛋白提取及浓度测定。SDS-PAGE电泳分离蛋白后将其转至PVDF膜上,用5%脱脂牛奶封闭处理1 h。添加一抗4 ℃孵育过夜,TBST缓冲液洗净后,加入二抗4 ℃孵育2 h后用TBST缓冲液洗净。ECL曝光显影,以β-actin作为内参,利用Image J软件对各个条带灰度值进行分析。
1.3 数据处理
使用Image J软件对图片中荧光强度及条带灰度值进行分析,使用GraphPad Prism 8.0.2进行统计分析,并通过t检验比较各组数据。实验数据以平均值±标准差(
2. 结果与分析
2.1 6′′′-阿魏酰斯皮诺素预处理抑制Aβ1-42诱导的SH-SY5Y细胞损伤
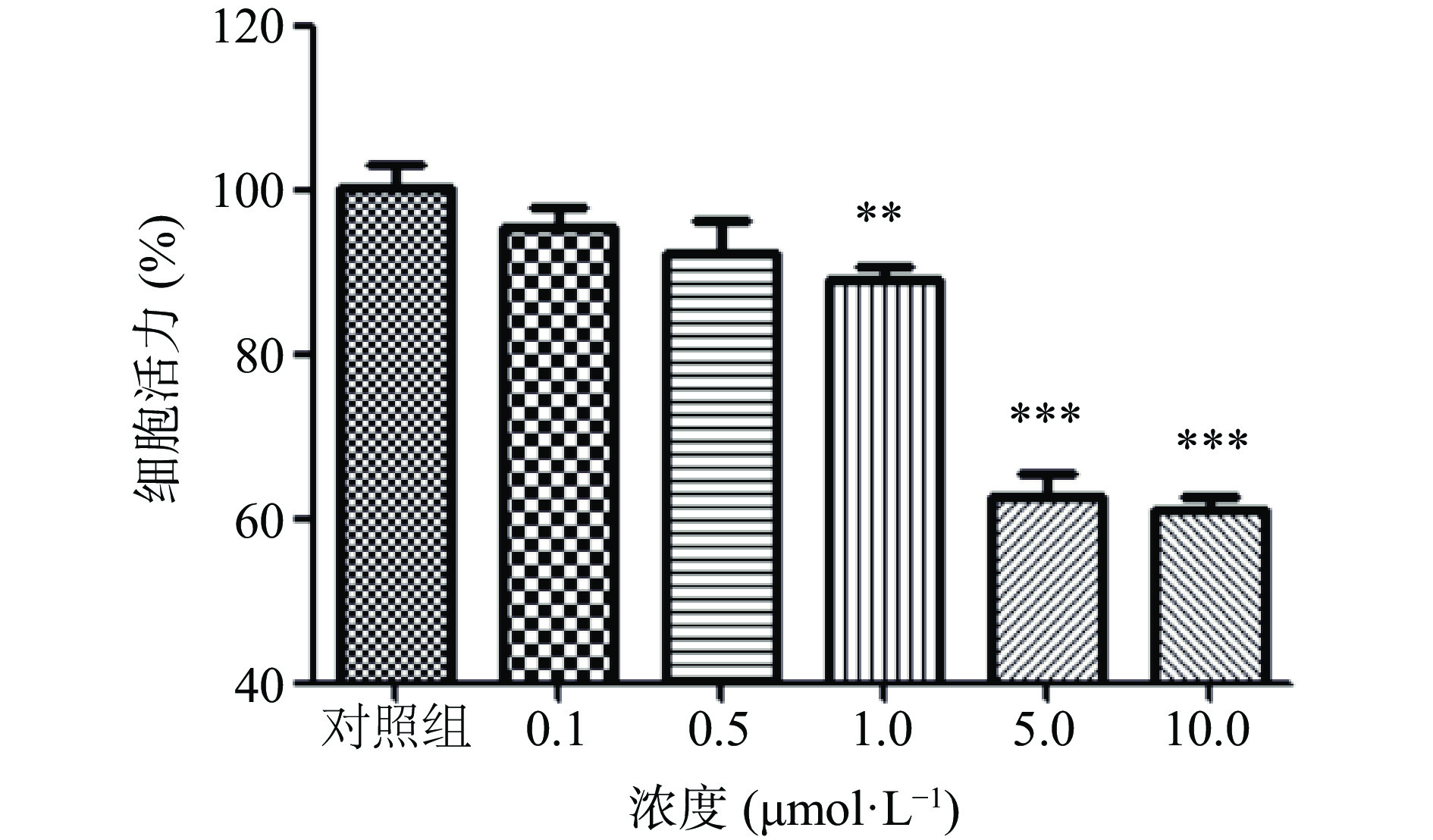
依照实验计划给予SH-SY5Y细胞6′′′-阿魏酰斯皮诺素及Aβ1-42,并利用CCK-8法检测细胞活力。Aβ1-42会对细胞造成损伤,且损伤程度呈剂量依赖性,结果如图1所示:在不给予6′′′-阿魏酰斯皮诺素的情况下,1、5、10 μmol/L Aβ1-42给药均导致细胞活力下降,与对照组相比,Aβ1-42 1 μmol/L组细胞活力降为89.17%,Aβ1-42 5 μmol/L组细胞活力降为62.18%,Aβ1-42 10 μmol/L组细胞活力降为60.96%。其中,1 μmol/L Aβ1-42虽然导致细胞活力显著下降(P<0.01),但此时细胞损伤程度较低;Aβ1-42 5 μmol/L组及Aβ1-42 10 μmol/L组细胞活力均极显著下降(P<0.001),且细胞活力下降程度相近,因此,本实验选择5 μmol/L的Aβ1-42处理细胞构建损伤细胞模型。6′′′-阿魏酰斯皮诺素预处理可以有效抑制Aβ1-42诱导的细胞活力下降,结果如图2所示。与模型组相比,使用6′′′-阿魏酰斯皮诺素预处理可以剂量依赖性地增加Aβ1-42诱导损伤的SH-SY5Y细胞活力,其中,6′′′-阿魏酰斯皮诺素低剂量组(1、5 μmol/L)效果不显著(P>0.05),中剂量组(10 μmol/L)以及高剂量组(20、40 μmol/L)效果极显著(P<0.001),本实验选用5、10、20 μmol/L 6'''-阿魏酰斯皮诺素预处理细胞,研究6'''-阿魏酰斯皮诺素预处理对Aβ1-42诱导损伤细胞的保护作用,并分析6'''-阿魏酰斯皮诺素对细胞的保护作用是否具有浓度依赖性。
![]()
![]() 图 2 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞活力的影响Figure 2. Effect of 6′′′-feruloylspinosin on the viability of SH-SY5Y cells damaged by Aβ1-42 induction
图 2 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞活力的影响Figure 2. Effect of 6′′′-feruloylspinosin on the viability of SH-SY5Y cells damaged by Aβ1-42 induction不同处理组Calcein-AM/PI染色情况如图3所示:对照组细胞生长情况良好,细胞多呈现椭圆形或梭形,生长密集,代表死细胞的红色荧光较弱;Aβ1-42处理24 h后细胞出现损伤,细胞多呈圆形,细胞生长密度减小,死细胞数量增多,不同浓度6′′′-阿魏酰斯皮诺素预处理后,死细胞数量减少,并且细胞密度上升。说明Aβ1-42处理后,细胞受损,细胞生长密度降低、形态改变且死细胞数目增多,而6′′′-阿魏酰斯皮诺素预处理可以改善上述情况,降低Aβ1-42诱导的细胞损伤。
![]() 图 3 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞形态的影响(100×)注:Calcein-AM:活细胞;PI:死细胞;Merge:相同位置两种荧光染色结果图片的叠加。Figure 3. Effect of 6′′′-feruloylspinosin on the morphology of SH-SY5Y cells damaged by Aβ1-42 induction (100×)
图 3 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞形态的影响(100×)注:Calcein-AM:活细胞;PI:死细胞;Merge:相同位置两种荧光染色结果图片的叠加。Figure 3. Effect of 6′′′-feruloylspinosin on the morphology of SH-SY5Y cells damaged by Aβ1-42 induction (100×)2.2 6′′′-阿魏酰斯皮诺素预处理缓解Aβ1-42诱导的SH-SY5Y细胞氧化损伤
SH-SY5Y细胞内ROS染色情况如图4(A)~图4(B)所示,与对照组相比,模型组细胞相对荧光强度极显著增强(P<0.001),6′′′-阿魏酰斯皮诺素预处理组细胞平均荧光强度减弱,且呈现剂量依赖性。MDA为氧自由基攻击生物膜中不饱和脂肪酸,生成的脂质过氧化产物[11];GSH-Px是一种重要的抗氧化酶,可以催化还原型谷胱甘肽与过氧化氢反应,生成氧化型谷胱甘肽与水[12]。因此,MDA水平以及GSH-Px酶活力常被用来衡量机体氧化损伤的程度及抗氧化能力的强弱。各组MDA水平及GSH-Px酶活力的检测结果如图4(C)~图4(D)所示,相较于对照组,模型组MDA水平极显著增加,GSH-Px酶活力极显著下降(P<0.001),与模型组相比,6′′′-阿魏酰斯皮诺素(20 μmol/L)预处理可以显著降低MDA水平(P<0.01),并显著提高GSH-Px酶活力(P<0.001)。
![]() 图 4 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞氧化应激情况的影响(40×)Figure 4. Effect of 6′′′-feruloylspinosin on the level of oxidative stress in SH-SY5Y cells damaged by Aβ1-42 induction (40×)
图 4 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞氧化应激情况的影响(40×)Figure 4. Effect of 6′′′-feruloylspinosin on the level of oxidative stress in SH-SY5Y cells damaged by Aβ1-42 induction (40×)2.3 6′′′-阿魏酰斯皮诺素预处理抑制Aβ1-42诱导的SH-SY5Y细胞线粒体膜电位降低
各组细胞线粒体膜电位水平测定结果如图5所示,对照组红色荧光较强,绿色荧光较弱,说明对照组中JC-1多在线粒体基质中以聚集体存在,发出红色荧光,此时线粒体膜电位较高。模型组绿色荧光较强,说明模型组细胞线粒体膜电位极显著下降(P<0.001),此时,JC-1从线粒体中扩散出来,以发出绿色荧光的JC-1单体形式存在[13]。6′′′-阿魏酰斯皮诺素组(10、20 μmol/L)红色荧光强度增强,绿色荧光强度减弱,说明6′′′-阿魏酰斯皮诺素可以显著或极显著提高Aβ1-42诱导损伤SH-SY5Y细胞线粒体膜电位(P<0.01、P<0.001)。
![]() 图 5 6′′′-阿魏酰斯皮诺素预处理对Aβ1-42诱导损伤SH-SY5Y细胞膜电位水平的影响(100×)注:Aggregation:高MMP细胞;Monomer:低MMP细胞;Merge:相同位置两种荧光染色结果图片的叠加。Figure 5. Effect of 6′′′-feruloylspinosin on the level of MMP in SH-SY5Y cells damaged by Aβ1-42 induction (100×)
图 5 6′′′-阿魏酰斯皮诺素预处理对Aβ1-42诱导损伤SH-SY5Y细胞膜电位水平的影响(100×)注:Aggregation:高MMP细胞;Monomer:低MMP细胞;Merge:相同位置两种荧光染色结果图片的叠加。Figure 5. Effect of 6′′′-feruloylspinosin on the level of MMP in SH-SY5Y cells damaged by Aβ1-42 induction (100×)2.4 6′′′-阿魏酰斯皮诺素预处理调节Aβ1-42诱导损伤的SH-SY5Y细胞内凋亡相关蛋白表达
6′′′-阿魏酰斯皮诺素及Aβ1-42对SH-SY5Y细胞中凋亡相关蛋白表达的影响如图6所示。相较于对照组,10 μmol/L 6′′′-阿魏酰斯皮诺素单独处理组细胞凋亡相关蛋白表达无显著变化(P>0.05)。5 μmol/L Aβ1-42处理后细胞内Bad蛋白表达极显著上调,Bcl-2蛋白表达极显著降低(P<0.001);6′′′-阿魏酰斯皮诺素预处理组Bad蛋白表达呈浓度依赖性地下降,Bcl-2蛋白的表达呈浓度依赖性地上升,且与模型组差异显著(P<0.01)。6′′′-阿魏酰斯皮诺素对健康细胞凋亡相关蛋白表达没有显著影响(P>0.05),但6′′′-阿魏酰斯皮诺素预处理可以调节诱导损伤细胞凋亡相关蛋白的表达,这表明6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤的SH-SY5Y细胞的保护作用可能与其调节凋亡相关蛋白表达的活性相关。
![]() 图 6 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞凋亡相关蛋白表达的影响Figure 6. Effect of 6′′′-feruloylspinosin on the expression of apoptosis-related proteins in SH-SY5Y cells damaged by Aβ1-42 induction
图 6 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞凋亡相关蛋白表达的影响Figure 6. Effect of 6′′′-feruloylspinosin on the expression of apoptosis-related proteins in SH-SY5Y cells damaged by Aβ1-42 induction3. 讨论与结论
阿尔兹海默症的典型病理特征为微管稳定蛋白质过度磷酸化形成神经原纤维缠结(neurofibrillary tangle,NFT)及Aβ组成的胞外老年斑(senile plaque,SP)[14-15]。其中,Aβ是淀粉样前体蛋白(amyloidprecursor protein,APP)经β/γ裂解酶切割产生的含有39~43个氨基酸的多肽[16-17]。早在20世纪60年代,研究者就在AD患者脑内发现了Aβ原纤维,并明确Aβ与AD的发生发展有着密切关系[18]。现代研究表明,Aβ低聚物及纤维体沉积在脑中,会引起神经细胞氧化应激、引发神经炎症、抑制海马区长时程增强效应等,导致神经细胞凋亡,从而损害患者的认知能力[19-21]。本研究利用Aβ1-42处理SH-SY5Y细胞,构建神经细胞损伤模型,发现5、10 μmol/L Aβ1-42可以极显著降低SH-SY5Y细胞活力(P<0.001),并改变SH-SY5Y细胞形态。10、20 μmol/L 6′′′-阿魏酰斯皮诺素预处理可以提高损伤SH-SY5Y细胞活力,增加细胞生长密度,改善细胞形态,这说明6′′′-阿魏酰斯皮诺素可以保护Aβ1-42诱导的损伤细胞,可能具有潜在的AD治疗作用。
大量研究表明,Aβ是AD发生发展过程中神经元氧化损伤的关键[22]。氧自由基的存在诱导AD模型小鼠脑内Aβ的沉积并加速AD病程发展[23],Aβ的存在也会损伤细胞抗氧化防御系统,导致细胞清除自由基的能力减弱[24]。丁轲等[25]采用活性追踪法对酸枣仁提取组分体外抗氧化活性进行比较,发现6′′′-阿魏酰斯皮诺素具有抗氧化活性。Yang等[10]也发现6′′′-阿魏酰斯皮诺素可以减轻热应激诱导的秀丽隐杆线虫氧化损伤,延长秀丽隐杆线虫寿命。基于此,本研究分析了6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤的SH-SY5Y细胞的保护作用与其抗氧化活性的关系。结果表明,5 μmol/L Aβ1-42处理SH-SY5Y细胞24 h后,细胞内ROS及MDA水平升高并且抗氧化酶GSH-Px酶活力极显著降低(P<0.001);20 μmol/L 6′′′-阿魏酰斯皮诺素预处理可以降低细胞内ROS及MDA累积水平并显著提高GSH-Px酶活力(P<0.01、P<0.001),减低Aβ1-42诱导的SH-SY5Y细胞氧化应激损伤。
线粒体功能障碍是AD发生的早期事件,线粒体作为细胞能量的主要来源及氧化还原反应的重要场所,极易遭受氧自由基的攻击而造成损伤,表现为线粒体呼吸链电子传递障碍、线粒体渗透性肿胀破碎、线粒体膜通透性转换孔过度开放等[26-28]。Bcl-2家族蛋白主要作用于线粒体外膜,与线粒体膜通透性及细胞凋亡密切相关[29]。正常情况下,Bcl-2家族抗凋亡蛋白成员与促凋亡蛋白成员以异源二聚体形式存在。当细胞受到刺激后,Bcl-2蛋白家族成员Bad发生去磷酸化,并从其与分子伴侣蛋白结合组成的无活性复合物中解离,破坏异源二聚体结构,置换出大量促凋亡蛋白Bcl-2相关X蛋白(Bcl2-associated X protein, Bax)聚集到线粒体外膜上,导致线粒体膜通透性转换孔过度开放,细胞色素C向胞质释放,激活caspase联级反应,诱导细胞凋亡[30-31]。本研究结果显示,5 μmol/L Aβ1-42会导致SH-SY5Y细胞线粒体膜电位极显著下降(P<0.001),并使抗凋亡蛋白Bcl-2表达极显著降低(P<0.001),促凋亡蛋白Bad表达极显著升高(P<0.001)。不同浓度6′′′-阿魏酰斯皮诺素预处理不仅可以上调Aβ1-42诱导损伤的SH-SY5Y细胞线粒体膜电位并且可以调节Bcl-2家族蛋白的表达,上调Bcl-2蛋白表达,下调Bad蛋白表达,其中,20 μmol/L 6′′′-阿魏酰斯皮诺素组效果显著(P<0.001、P<0.01、P<0.01)。
综上所述,6′′′-阿魏酰斯皮诺素可以抑制Aβ1-42诱导的SH-SY5Y细胞损伤,其作用机制可能与6′′′-阿魏酰斯皮诺素抗氧化活性及调控Bcl-2家族蛋白表达等活性相关。
-
![]()
![]()
图 2 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞活力的影响
Figure 2. Effect of 6′′′-feruloylspinosin on the viability of SH-SY5Y cells damaged by Aβ1-42 induction
![]()
图 3 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞形态的影响(100×)
注:Calcein-AM:活细胞;PI:死细胞;Merge:相同位置两种荧光染色结果图片的叠加。
Figure 3. Effect of 6′′′-feruloylspinosin on the morphology of SH-SY5Y cells damaged by Aβ1-42 induction (100×)
![]()
图 4 6′′′-阿魏酰斯皮诺素对Aβ1-42诱导损伤SH-SY5Y细胞氧化应激情况的影响(40×)
Figure 4. Effect of 6′′′-feruloylspinosin on the level of oxidative stress in SH-SY5Y cells damaged by Aβ1-42 induction (40×)
![]()
图 5 6′′′-阿魏酰斯皮诺素预处理对Aβ1-42诱导损伤SH-SY5Y细胞膜电位水平的影响(100×)
注:Aggregation:高MMP细胞;Monomer:低MMP细胞;Merge:相同位置两种荧光染色结果图片的叠加。
Figure 5. Effect of 6′′′-feruloylspinosin on the level of MMP in SH-SY5Y cells damaged by Aβ1-42 induction (100×)
-
[1] 黄之镨, 马伟光. 酸枣仁及活性物质的药理研究进展[J]. 中国民族民间医药,2018,27(3):57−60. [HUANG Z P, MA W G. Research progress in the pharmacological of Semen Ziziphi Spinosae and its active ingredients[J]. Chinese Journal of Ethnomedicine and Ethnopharmacy,2018,27(3):57−60. HUANG Z P, MA W G. Research progress in the pharmacological of Semen Ziziphi Spinosae and its active ingredients[J]. Chinese Journal of Ethnomedicine and Ethnopharmacy, 2018, 27(3): 57-60.
[2] 武宝爱, 闫锋, 申晨曦, 等. UPLC-MS/MS法结合化学计量学法分析酸枣仁和理枣仁中多指标成分[J]. 中草药,2021,52(8):2400−2407. [WU B A, YAN F, SHEN C X, et al. Study on multi-index components of Ziziphi Spinosae Semen and Ziziphi Mauritianae Semen according to UPLC-MS/MS coupled with chemometrics[J]. Chinese Traditional and Herbal Drugs,2021,52(8):2400−2407. WU B A, YAN F, SHEN C X, et al. Study on multi-index components of Ziziphi Spinosae Semen and Ziziphimauritianae Semen according to UPLC-MS/MS coupled with chemometrics[J]. Chinese Traditional and Herbal Drugs, 2021, 52(8): 2400-2407.
[3] 周赛男, 陈安家, 郭宝林, 等. 不同产区酸枣仁中斯皮诺素、6′′′-阿魏酰斯皮诺素、酸枣仁皂苷A、酸枣仁皂苷B的含量测定[J]. 中草药,2019,50(11):2712−2717. [ZHOU S N, CHEN A J, GUO B L, et al. Determination of two flavonoids and two saponins in Ziziphi Spinosae Semen from different producing areas[J]. Chinese Traditional and Herbal Drugs,2019,50(11):2712−2717. doi: 10.7501/j.issn.0253-2670.2019.11.031 ZHOU S N, CHEN A J, GUO B L, et al. Determination of two flavonoids and two saponins in Ziziphi Spinosae Semen from different producing areas[J]. Chinese Traditional and Herbal Drugs, 2019, 50(11): 2712-2717. doi: 10.7501/j.issn.0253-2670.2019.11.031
[4] YANG T, FANG L, LIN T, et al. Ultrasonicated sour Jujube seed flavonoids extract exerts ameliorative antioxidant capacity and reduces Aβ-induced toxicity in Caenorhabditis elegans[J]. Journal of Ethnopharmacology,2019,239:111886. doi: 10.1016/j.jep.2019.111886
[5] LI Y J, BI K S. Study on the therapeutic material basis of traditional Chinese medicinal preparation suanzaoren decoction[J]. Chemical & Pharmaceutical Bulletin,2006,54(6):847−851.
[6] NICHOLS E, SZOEKE CEI, VOLLSET SE, et al. Global, regional, and national burden of Alzheimer's disease and other dementias, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016[J]. The Lancet Neurology,2019,18(1):88−106. doi: 10.1016/S1474-4422(18)30403-4
[7] PRAKASH A, KUMAR A. Role of nuclear receptor on regulation of BDNF and neuroinflammation in hippocampus of beta-amyloid animal model of Alzheimer's disease[J]. Neurotoxicity Research,2014,25(4):335−347. doi: 10.1007/s12640-013-9437-9
[8] LIU Y, WENG W, GAO R, et al. New insights for cellular and molecular mechanisms of aging and aging-related diseases: Herbal medicine as potential therapeutic approach[J]. Oxidative Medicine and Cellular Longevity,2019,2019:4598167.
[9] 明敬峰, 蔡增林. 中药治疗帕金森病的研究进展及潜在价值[J]. 实用医学杂志,2017,33(12):1904−1907. [MING J F, CAI Z L. Research progress and potential value of traditional Chinese medicine in the treatment of Parkinson's disease[J]. The Journal of Practical Medicine,2017,33(12):1904−1907. doi: 10.3969/j.issn.1006-5725.2017.12.002 MING J F, CAI Z L. Research progress and potential value of traditional Chinese medicine in the treatment of Parkinson's disease[J]. The Journal of Practical Medicine, 2017, 33(12): 1904-1907. doi: 10.3969/j.issn.1006-5725.2017.12.002
[10] YANG T, ZHAO X, ZHANG Y, et al. 6‴-Feruloylspinosin alleviated beta-amyloid induced toxicity by promoting mitophagy in Caenorhabditis elegans (GMC101) and PC12 cells[J]. Science of the Total Environment,2020,715:136953. doi: 10.1016/j.scitotenv.2020.136953
[11] 翟晓虎, 杨海锋, 陈慧英, 等. 丙二醛的毒性作用及检测技术研究进展[J]. 上海农业学报,2018,34(1):144−148. [ZHAI X H, YANG H F, CHEN H Y, et al. Research progress in the toxicity and determination technology of malondialdehyde[J]. Acta Agriculturae Shanghai,2018,34(1):144−148. ZHAI X H, YANG H F, CHEN H Y, et al. Research progress in the toxicity and determination technology of malondialdehyde[J]. ActaAgriculturae Shanghai, 2018, 34(1): 144-148.
[12] BRAIDY N, ZARKA M, JUGDER B E, et al. The precursor to glutathione (GSH), γ-Glutamylcysteine (GGC), can ameliorate oxidative damage and neuroinflammation induced by Aβ 40 oligomers in human astrocytes[J]. Frontiers in Aging Neuroscience,2019,11:177. doi: 10.3389/fnagi.2019.00177
[13] 李妍, 赵东海, 张巍, 等. 胡皂苷d对人神经母细胞瘤SH-SY5Y细胞线粒体膜电位及细胞凋亡的影响[J]. 上海中医药杂志,2016,50(6):90−93. [LI Y, ZHAO D H, ZHANG W, et al. Effects of Saikosaponin d on mitochondrial membrane potential and apoptosis in human neuroblastoma SH-SY5Y cells[J]. Shanghai Journal of Traditional Chinese Medicine,2016,50(6):90−93. LI Y, ZHAO D H, ZHANG W, et al. Effects of Saikosaponin d on mitochondrial membrane potential and apoptosis in human neuroblastoma SH-SY5Y cells[J]. Shanghai Journal of Traditional Chinese Medicine, 2016, 50(6): 90-93.
[14] LI S, JIN M, LIU L, et al. Decoding the synaptic dysfunction of bioactive human AD brain soluble Aβ to inspire novel therapeutic avenues for Alzheimer's disease[J]. Acta Neuropathologica Communications,2018,6(1):121. doi: 10.1186/s40478-018-0626-x
[15] BUSCHE M A, WEGMANN S, DUJARDIN S, et al. Tau impairs neural circuits, dominating amyloid-beta effects, in Alzheimer models in vivo[J]. Nature Neuroscience,2019,22(1):57−64. doi: 10.1038/s41593-018-0289-8
[16] BUSCIGLIO J, GABUZDA DH, MATSUDAIRA P, et al. Generation of beta-amyloid in the secretory pathway in neuronal and nonneuronal cells[J]. Proceedings of the National Academy of Sciences of the United States of America, 1993(5), 90: 2092−2096.
[17] MULLER U C, DELLER T, KORTE M. Not just amyloid: Physiological functions of the amyloid precursor protein family[J]. Nature Reviews Neuroscience,2017,18(5):281−298. doi: 10.1038/nrn.2017.29
[18] KIDD M. Paired helical filaments in electron microscopy of Alzheimer's disease[J]. Nature,1963,197(4863):192−193.
[19] CHEIGNON C, TOMAS M, BONNEFONTR D, et al. Oxidative stress and the amyloid beta peptide in Alzheimer’s disease[J]. Redox Biology,2018,14:450−464. doi: 10.1016/j.redox.2017.10.014
[20] HUNTER S, BRAYNE C. Understanding the roles of mutations in the amyloid precursor protein in Alzheimer disease[J]. Molecular Psychiatry,2018,23(1):81−93. doi: 10.1038/mp.2017.218
[21] ONO K, TSUJI M. Protofibrils of amyloid-beta are important targets of a disease-modifying approach for Alzheimer's disease[J]. International Journal of Molecular Sciences,2020,21(3):952−964. doi: 10.3390/ijms21030952
[22] PANZA F, LOZUPONE M, BELLOMO A, et al. Do anti-amyloidbeta drugs affect neuropsychiatric status in Alzheimer's disease patients?[J]. Ageing Research Reviews,2019,55:100948−100952. doi: 10.1016/j.arr.2019.100948
[23] KANAMARU T, KAMIMURA N, YOKOTA T, et al. Oxidative stress accelerates amyloid deposition and memory impairment in a double-transgenic mouse model of Alzheimer’s disease[J]. Neuroscience Letters,2015,587:126−131. doi: 10.1016/j.neulet.2014.12.033
[24] FENG Y , WANG X C. Antioxidant therapies for Alzheimer's disease[J]. Oxidative Medicine and Cellular Longevity,2012,2012:472932.
[25] 丁轲, 马崟松, 韩涛, 等. 酸枣仁抗氧化成分的分离及其相互作用研究[J]. 高校化学工程学报,2013,27(5):842−853. [DING K, MA Y S, HAN T, et al. Separation and interaction of the ingredients of antioxidant activities in Zizyphi Spinosi Semen[J]. Journal of Chemical Engineering of Chinese Universities,2013,27(5):842−853. doi: 10.3969/j.issn.1003-9015.2013.05.019 DING K, MA Y S, HAN T, et al. Separation and interaction of the ingredients of antioxidant activities in Zizyphi Spinosi Semen[J]. Journal of Chemical Engineering of Chinese Universities, 2013, 27(5): 842-853. doi: 10.3969/j.issn.1003-9015.2013.05.019
[26] VARDA S B, EDNA N C, ANNA S K, et al. VDAC1, mitochondrial dysfunction, and Alzheimer's disease[J]. Pharmacological Research,2018,131:87−101. doi: 10.1016/j.phrs.2018.03.010
[27] ISLAM M T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders[J]. Neurological Research,2017,39(1):73−82. doi: 10.1080/01616412.2016.1251711
[28] 胡钰慧, 何玲. 线粒体功能障碍与阿尔兹海默病发病的相关性研究进展[J]. 药学研究,2020,39(2):101−104. [HU Y H, HE L. Advances in the study of the relationship between mitochondrialdysfunction and the pathogenesis of Alzheimer's disease[J]. Journal of Pharmaceutical Research,2020,39(2):101−104. HU Y H, HE L. Advances in the study of the relationship between mitochondrialdysfunction and the pathogenesis of Alzheimer's disease[J]. Journal of Pharmaceutical Research, 2020, 39(2): 101-104.
[29] LINDSAY J, ESPOSTI M D, GILMORE A P. Bcl-2 proteins and mitochondria-Specificity in membrane targeting for death[J]. BBA-Molecular Cell Research,2011,1813(4):532−539.
[30] HOWELLS C C, BAUMANN W T, SAMUELS D C, et al. The Bcl-2-associated death promoter (BAD) lowers the threshold at which the Bcl-2-interacting domain death agonist (BID) triggers mitochondria disintegration[J]. Journal of Theoretical Biology,2011,271(1):114−123. doi: 10.1016/j.jtbi.2010.11.040
[31] 孙秀玉, 刘立亚, 吴宥熹, 等. Bcl-2家族对线粒体质量控制的调控研究[J]. 中国药理学通报,2015,31(12):1633−1636. [SUN X Y, LIU L Y, WU Y X, et al. On mitochondrial quality control regulated by Bcl-2 family[J]. Chinese Pharmacological Bulletin,2015,31(12):1633−1636. doi: 10.3969/j.issn.1001-1978.2015.12.002 SUN X Y, LIU L Y, WU Y X, et al. On mitochondrial quality control regulated by Bcl-2 family[J]. Chinese Pharmacological Bulletin, 2015, 31(12): 1633-1636. doi: 10.3969/j.issn.1001-1978.2015.12.002
-
期刊类型引用(2)
1. 安容慧,陈兴开,常子安,任紫烟,张婕,连欢,贾连文,杨相政. 采后不同时间压差预冷对水蜜桃货架品质和香气成分的影响. 食品工业科技. 2024(09): 317-324 .  本站查看
本站查看
2. 隋海涛,陈东杰,王凤丽,邹泽宇,郭风军,马倩倩,隋青,张长峰,孙崇德. 桃果实采后品质变化机制及调控技术研究进展. 中国果菜. 2024(12): 1-8+19 . 百度学术
其他类型引用(2)


 下载:
下载:






 下载:
下载:





计量
- 文章访问数: 251
- HTML全文浏览量: 57
- PDF下载量: 25
- 被引次数: 4



