


Research Progress on Eurotium cristatum and Its Fermentation Application
-
摘要: 茯砖茶在加工过程中会“发花”,产生金黄色颗粒,而冠突散囊菌是茯砖茶“发花”的关键微生物,对茶的品质起关键作用。冠突散囊菌发酵可促进茶叶内含成分转化,改善产品香气滋味,且产生多种活性代谢物,具有抑菌、降血糖、调节肠道菌群、抗氧化等功效,已经成为重要的资源微生物得到广泛应用。本文综述了冠突散囊菌的基本生物学特性,丰富的酶系,多样的活性代谢物,安全性,以及在茶叶、谷豆类及中药材深加工等领域发酵应用的物质和生物活性变化等内容,并对存在的问题和未来的研究方向进行了分析和展望,以期为其深入研究和开发提供帮助。Abstract: Eurotium cristatum which produces golden particles, is the dominant fungus playing a key role in the “flowering” process and in the quality of Fu-brick tea. It can produce a variety of active enzymes and active metabolites, which have multiple biological effects, such as improving the quality of tea, antibacterial, hypoglycemic, regulating intestinal flora, antioxidant and other effects. As an important resource microorganism, Eurotium cristatum has been widely used. In this paper, the basic biological characteristics, rich enzyme system, diverse active metabolites, safety of Eurotium cristatum, as well as the changes of substances and biological activities in the processing of tea, beans, cereals, and Chinese herbs by Eurotium cristatum fermentation are reviewed. The existing problems and future research directions are analyzed and prospected, in order to provide reference for its in-depth research and development.
-
Keywords:
- Eurotium cristatum /
- fermentation /
- processing of Chinese herbs /
- food processing /
- enzyme
-
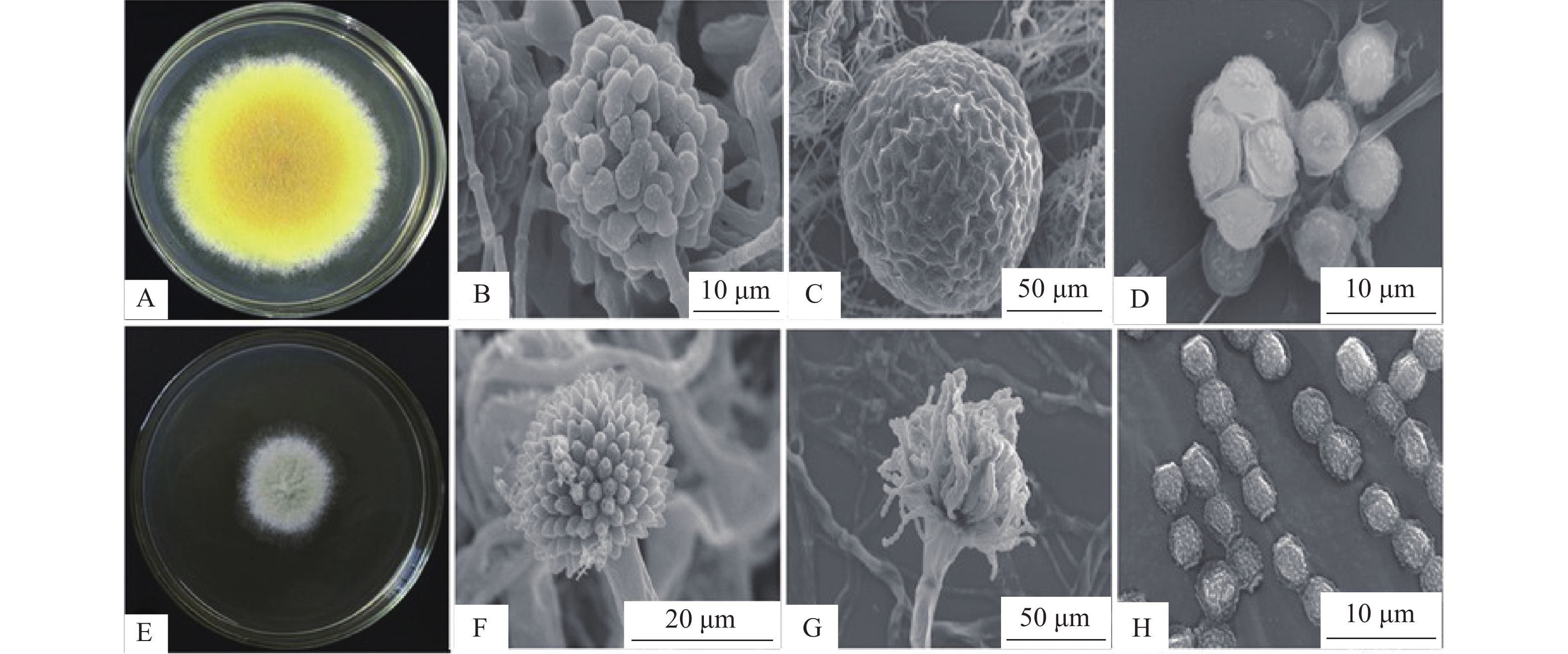
茯砖茶是以黑毛茶为原料,经过汽蒸、渥堆、发花、干燥等一系列复杂工艺制作而成的后发酵茶,其中发花是茯砖茶加工的关键步骤,决定着茶的品质。发花阶段,优势“金花菌”大量繁殖,茶叶在微生物作用下发生深度的生理生化变化,形成茯砖茶独特的风味[1-5]。通过形态学种群分析,DNA条形码和多位点测序等方法分析“金花菌”的进化关系,发现“金花”实际是冠突散囊菌的金黄色闭囊壳(图1),而冠突散囊菌是茯砖茶的唯一优势真菌[5]。在国家标准中,茯砖茶是唯一要求冠突散囊菌这项指标的黑茶类品种[6]。六堡茶等黑茶中也分离出冠突散囊菌[7]。本文主要讨论黑茶冠突散囊菌的生物学特性、酶系、代谢产物和安全性等特性,以及其发酵应用的研究进展。
![]()
1. 冠突散囊菌的生物学特性
冠突散囊菌(Eurotium cristatum)属子囊菌纲,曲霉目,曲霉科,作为产有性型和无性型的全型真菌,其典型特征是具冠状突起及表面明显粗糙具小疣的子囊孢子和具小刺的分生孢子。根据其孢子的特征,将该菌有性型鉴定为冠突散囊菌(Eurotium cristatum),无性型鉴定为针刺曲霉菌(Aspergillus spiculosus Blaster),别名为冠突曲霉菌(Aspergillus cristatum Blaster)[8-9]。
冠突散囊菌好氧,耐干燥,最适pH为5,最适生长温度为28~30 ℃。该菌适应性较强,在茶砖和低渗培养基(0.5 mol/L NaCl)上行有性繁殖,繁殖方式随渗透压升高逐渐转变为无性繁殖,高渗透压(≥3 mol/L NaCl)条件下仅行无性繁殖[10],其孢子在85 ℃冲泡5 min后仍可存活[11]。Ca2+信号途径及VeA基因在其适应高渗环境[12]和孢子形成[13]中发挥重要作用。碳源和氮源显著影响菌丝生长和产孢过程[14-16],其中碳源为产孢所必需[14]。不同来源菌株对不同种类碳源的利用程度不同。固态发酵以D-果糖、葡萄糖、蔗糖、D-木糖、麦芽糖等为佳,而对山梨醇、琼脂、单宁和淀粉等利用较差;液态发酵对碳源种类依赖性较低[14-16]。碳源浓度[15]和氮源种类[16]亦影响产孢的类型和数量。无机氮源促进孢子萌发效果较好,而有机氮源对菌落生长的促进作用较大[16],因此,在实际培养中往往采取两者结合的方式。
2. 冠突散囊菌的酶系
冠突散囊菌能分泌多种胞外酶,如纤维素酶、蛋白酶和多酚氧化酶等。在茯砖茶“发花”过程中,冠突散囊菌通过分泌胞外酶促使茶叶中的茶多酚发生氧化、缩合,形成茶黄素、茶红素、茶褐素等物质,淀粉和纤维素等不溶性多糖分解转化为葡萄糖和果糖,蛋白质分解为氨基酸等,且使氨基酸、糖类、咖啡碱等各成分之间发生聚合、缩合等一系列的复杂反应[2,4,17],消除茶叶的粗青、苦涩味,使茶叶在口味上更加醇和爽口,甜滑回甘,汤颜色红亮,形成茯砖茶特定的风味、香气、色泽,并且增加茶叶中的活性物质。纤维素酶在茯砖茶渥堆过程中能软化老叶,降解粗纤维为可溶性糖。高温杀青使茶叶中内源纤维素酶活性被急剧钝化,渥堆和“发花”过程中微生物的代谢活动使纤维素酶活性明显增强[18]。阿魏酸酯酶和木聚糖酶协同作用于晒青毛茶可显著提高其阿魏酸和低聚木糖的含量,增加茶汤活性物质[19]。胞外多酚氧化酶、漆酶等可将儿茶素氧化为茶黄素、茶红素和茶褐素等水溶性的色素[17,20-22]。过氧化物酶、过氧化氢酶、多酚氧化酶和总超氧化物歧化酶是冠突散囊菌发酵茶抗氧化的物质基础[23]。其淀粉酶和蛋白酶等消化酶,可显著提高人体肠道中蛋白酶、淀粉酶的活性[24-25],与茯砖茶的促消化作用密不可分。糖苷酶可将西洋参、高丽参的其他皂苷成分有效转化为人参皂苷Rd及多种未知结构的高保留性的皂苷产物 [26]。酶是冠突散囊菌进行发酵应用的重要基础,其性质尚需进行深度研究。
3. 冠突散囊菌的代谢产物
3.1 色素
冠突散囊菌在马铃薯等培养基上生长代谢通常会产生大量的色素物质,主要为黄色素和黑色素[27]。黄色素的含量与有性孢子数量正相关,与“金花”的颜色有关[27]。以石油醚提取陕西茯砖茶冠突散囊菌液态发酵的黄色素,发现其为脂溶性色素,最大吸收波长为398 nm,不耐高温,耐酸不耐碱,金属离子(Fe3+、Fe2+、Cu2+)、太阳光等条件会使其降解[28]。光谱学分析推测黄色素含有-C-H、C-O、C=C等官能团[28-29]。黑色素是一类结构复杂多样的酚类或吲哚类生物大分子色素的总称,具有避免光伤害、强抗氧化性、抗辐射、免疫调节等作用。冠突散囊菌所产胞外黑色素的最大吸收波长298 nm,在70 ℃以下耐热性强,Mg2+、Zn2+、Fe3+对其的稳定性有增强作用,遇光、氧化还原剂易分解[30-31]。冠突散囊菌色素的理化性质研究尚有待深入。
3.2 生物活性代谢物
冠突散囊菌含有多种生物活性代谢物,如黄酮、生物碱、哌嗪二酮衍生物和多糖[32-33]等。在脂多糖诱导的RAW264.7炎症细胞模型中,苯甲醛衍生物通过抑制iNOS和COX-2的表达发挥抗炎作用[34]。水溶性二酮哌嗪二聚体生物碱eurocristatine在糖尿病老鼠模型中可通过调节PI3K/AKT信号通路达到与二甲双胍相当的降糖效果[32- 33]。脂溶性组分包含异戊烯基吲哚二酮哌嗪生物碱cristatumin F以及echinulin、dehydroechinulin、 neoechinulin A和variecolorin O,其中cristatumin F有DPPH自由基清除活性和对3T3L1细胞的边缘衰减作用[35]。真菌多糖是冠突散囊菌的重要的活性代谢物。菌株MK346334的粗糖提取液,尤其是EPS2组分,显示出与肠道微生物群调节相关的免疫调节活性[36]。从菌丝体分离的多糖ECIP-1A,分子量为24.3 kD,主要由甘露糖、葡萄糖和半乳糖等单糖组成,体外对巨噬细胞RAW264.7有显著的免疫调节活性[37]。发酵初期的冠突散囊菌菌丝能产生较高含量的洛伐他汀(降胆固醇药物)[14,38]。混菌共培养可拓宽真菌次级代谢产物谱,冠突散囊菌与金黄色葡萄球菌固体混合发酵产生两种新化合物(tetrahydroauroglaucine和epiheveadride),其混合代谢物对A549细胞等有较好的细胞毒活性[39]。
4. 冠突散囊菌的安全性
冠突散囊菌曾被认为是有毒真菌。但急性毒理学试验显示,大鼠经口灌胃对冠突散囊菌的最大耐受剂量大于7.31×1010个/mL,对其发酵成的茯砖茶的LD50大于5001 mg/kg。喂养30 d后,各剂量组菌液和茶叶浸出液对大鼠在体重、血液学指标、脏器系数及一般行为方面均无显著性影响[11,40]。Ge等[10]通过系统分析比较转录组学和基因组学的数据发现冠突散囊菌不含毒素生物合成相关的基因簇。进一步检测常见的6种毒素发现,其仅产生低剂量的伏马菌素B1(0.17 ppm),远低于美国食品与药品监督总局食品安全国家标准的限量(2 ppm),而我国对此毒素尚无限量标准。以冠突散囊菌(CB10001)或其孢子饲喂高脂饮食的C57BL/6J小鼠,发现该菌可在小鼠肠道存活(超过2周时间),并可有效阻止小鼠增重,缓解炎症,调节肠道菌群,恢复肠道稳态[11]。冠突散囊菌可通过抑制NF-κB和MAPK信号通路的激活来抑制结肠炎症反应,保护肠道屏障,从而改善溃疡性结肠炎小鼠的结肠缩短、体质量下降和评定疾病活动指数评分等指标,修复葡萄糖硫酸钠造成的病理组织学损害[41]。另外,茯砖茶在我国西北地区有悠久的饮用历史,研究显示其具有降脂减肥、增强免疫、调节糖代谢和调节肠道菌群等多种生物学功效 [11,42-45]。因此,判定冠突散囊菌为实际无毒物,且为潜在的益生菌,为其推广应用提供了依据。
5. 冠突散囊菌的发酵应用
5.1 茶叶加工
茶叶加工过程中人工接种冠突散囊菌可促进茶品质转化,加速发酵进程,缩短生产周期,利于全程清洁化生产,保证品质均一性。发酵过程以固态发酵为主,又可分为普通茶发酵和植物代茶发酵两种。多数茶叶经冠突散囊菌发酵后,苦涩味降低,散发独特菌花香,茶褐素增多,口感和品质得到提升。茶叶所含的化合物——香味物质、呈味物质、色泽和生物活性物质等均发生显著变化,进一步影响了茶叶的感官性状和功能活性(表1)。
表 1 不同品种茶在冠突散囊菌发酵前后性状对比Table 1. Comparison of characteristics of different tea varieties before/after fermentation by Eurotium cristatum类型 茶品种 发酵条件 主成分变化 生物学活性 感官变化 文献 茶 茯砖茶 固态 提高:茶多糖和茶褐素;
降低:茶多酚、EGCG、游离氨基酸、
咖啡碱、茶黄素、茶红素香味提升,色泽更佳 [4-5] 普洱茶 固态 提高:总黄酮(19.15%),非酯型儿茶素(52.85%),酯类气体(29.78%)、醇类气体(20.84%);
降低:酯型儿茶素(33.64%)、总酚(22.31%)、
氨基酸(24.34%)苦涩味降低,香味提升 [48] 绿茶 固态/液态 降低:茶多酚 口感醇和、苦涩味降低;香味提升,兼具茶香、菌花香和甜香 [17,46,49] 红茶 液态 香味提升 [49] 复配藤茶 固态 提高:可溶性糖(4.65%)、咖啡碱(3.9%);
降低:多酚(49.68%)、氨基酸(56.79%)、二氢杨梅(39.29%)、儿茶素酸涩味降低、
口感提升[53] 白毫毛尖茶 固态 降低:茶多酚 降低:抗氧化、清除自由基能力 香味提升 [7] 植
物
代
茶蒲公英茶 固态 降低:多酚(19.76%) 香气协调、无异味 [57] 荔枝草茶 固态 提高:茶黄素(3.02倍)、茶红素(1.73倍)、茶褐素(1.29倍);
降低:茶多酚(31.9%)、氨基酸(36.3%)、可溶性糖(25.2%)苦涩味减少、
香味提升[58] 刺五加茶 固态 提高:茶褐素(18.64%);降低:茶多酚(66.75%)、氨基酸(69.05%)、可溶性糖(55.1%) 总抗氧化活性提高、对DPPH自由基
清除率下降苦涩味减少、
香味提升[56] 甜茶 固态 提高:茶多酚、黄酮;降低:游离氨基酸和可溶性糖 提高:抗氧化;降低:清除自由基能力 香味提升 [54] 桑叶茶 固态 提高:还原糖(10.47%)、总糖(22.36%)、氨基酸(22.87%) 营养、保健品质提升 [50−52] 苦丁茶 固态 苦味、粗青气下降;增加菌花香和甜香 [59] 杜仲茶 固态 降低:茶多酚、咖啡碱、氨基酸、可溶性糖 醇和、回甘、菌花香浓 [51] 栘㐿茶 固态 提高:根皮素;
降低:根皮苷抗氧化和抑菌
活性增强[60] 5.1.1 气味物质变化
经冠突散囊菌适度发酵的茶叶,风味差异明显,香气物质含量与种类均有所增加,香气协调,兼具茶香和菌花香[17,46-48]。发酵可促进萜醇类物质的代谢,尤其是芳樟醇氧化物的含量显著增加。发酵茶汤所含的醛类气体减低,而酯类气体含量显著提高[17,46-48]。研究显示,在以茶叶的碳水化合物为碳源时,冠突散囊菌一方面通过MEP途径分解糖苷前体提供能量,产生大量的游离芳香化合物[47];另一方面积累了大量的苯丙酸/苯环类化合物途径的中间代谢物,包括苯甲醇和苯乙醇,以及大量的甲酯化合物,如水杨酸甲酯、苯甲酸甲酯和苯乙酸甲酯[48]。这些化合物共同作用产生了特有的茶香和花香味。
5.1.2 呈味物质变化
冠突散囊菌发酵的茶汤,滋味甘甜醇厚。茶叶中负责不同口感的各种氨基酸含量显著减低,可能与美拉德反应和菌的生长代谢相关[17]。研究表明,冠突散囊菌发酵后的茶叶中总酚,尤其是茶叶苦涩味形成的关键化合物儿茶素类含量显著下降[17,46,48-49]。其中,表没食子儿茶素没食子酸酯(EGCG)、表没食子儿茶素(EGC)、表儿茶素没食子酸酯(ECG)和表儿茶素(EC)等茶多酚含量下降最为显著[17,46]。酯型儿茶素水解能产生非酯型儿茶素和没食子酸(GA)[17,48]。EC、GA和EGC的含量随着菌代谢进一步降低[17]。另外,在影响茶叶苦味的生物碱(茶碱、咖啡碱、可可碱等)中,主要是咖啡碱的含量显著下降[17]。
5.1.3 色泽变化
冠突散囊菌“发花”过程对茶汤颜色影响显著,多数随着发酵程度的加深,茶叶颜色变深。茶黄素、茶红素等的含量先增加后减少,而茶褐素大幅增多[17,46-48]。过氧化物酶和漆酶能持续转化茶多酚成茶红素、茶黄素,而茶黄素、茶红素能与儿茶素以及其他化合物进一步通过聚合、缩合、偶联和氧化等作用形成茶褐素[17,46]。通过分泌多种胞外酶可促进了茶褐素的形成。茶褐素生物学活性较高,其含量的增加使茶汤颜色呈现棕红色,且可能是发酵茶主要的呈味物质[17,46]。另外,冠突散囊菌自身产生的黄色素、黑色素等色素,是否也对茶叶、茶汤的颜色产生了影响,目前尚无定论。
5.1.4 其他物质变化
由于纤维素酶、淀粉酶等酶的活性以及发酵环境的差异,冠突散囊菌发酵对茶叶可溶性糖含量的影响不尽一致。桑叶茯茶、杜仲茯茶和MF800948发酵的秋季绿茶的水溶性碳水化合物皆降低[17,50-52],而“金花藤茶”的可溶性糖含量增加[53]。“金花”普洱[48]和“金花”甜茶[54]的总黄酮含量提高,而藤茶拼配红茶发酵制作的“金花藤茶”多酚、二氢杨梅素的含量显著减少[53]。
5.1.5 生物活性变化
茶叶内含物的变化影响发酵茶的生物学活性[55]。“金花”普洱总抗氧化活性和自由基清除能力皆提升[48],冠突散囊菌发酵的甜茶[54]和刺五加茶[56]的总抗氧化活性提升,而羟自由基与DPPH自由基清除活性却显著下降。以广西六堡茶冠突曲霉(E6)发酵当地的“白毫毛尖”[7]和桑叶茶[50-52],其DPPH降解能力、总抗氧化能力均有变化。上述研究为冠突散囊菌发酵茶产品的开发奠定了基础。
5.2 谷类和豆类食品加工
冠突散囊菌可利用多种碳源生长,亦被用于谷类和豆类的加工过程,见表2。其发酵的燕麦面酱,色泽鲜艳,酱香浓郁,咸甜适宜,多酚含量为原料燕麦的6倍[61]。添加茯砖茶发酵糯米产生了35种特异性的香味物质,α−淀粉酶、蛋白酶和β−葡萄糖苷酶活性增强,总酚含量提高[62−63]。冠突散囊菌固态发酵甜荞,相比毛霉或根霉,对黄酮和总酚含量的促进作用更强[64-65];发酵苦荞,清蛋白、球蛋白、醇蛋白和总蛋白的含量增加[66]。以冠突散囊菌(YL-1)固态发酵黄豆[67]和黑豆[68],总酚、黄豆苷元、金雀异黄酮、大豆异黄酮和总苷元等的含量均显著提高;挥发性物质差异明显,其中酯含量增加,若干醇和醛含量减少。冠突散囊菌发酵的豆渣产品Okara,以两种蒽醌(大黄素和大黄素甲醚)为标志性代谢物,其α-糖苷酶抑制活性更强,能有效降低小鼠餐后血糖水平[69]。研究显示,固态发酵能破坏植物细胞壁或水解细胞壁组分的共价键和不可溶的结合性多酚,导致酚类化合物的释放[65]。上述加工食品中多酚和黄酮含量的升高,与发酵过程中酶的作用,尤其是冠突散囊菌水解酶活性密切相关[65,67]。多酚和黄酮含量的升高,使得冠突散囊菌的谷类和豆类食品体外抗氧化性、自由基清除能力以及糖脂代谢相关酶抑制率等生物学活性增强显著[65-68]。因此,这类发酵制品可作为功能性食品或者天然食品添加剂而成为有效的抗氧化剂。
表 2 谷类和豆类经冠突散囊菌发酵前后性状对比Table 2. Comparison of characteristics of different food varieties before/after fermentation by Eurotium cristatum食材 发酵状态 化合物变化 生物学活性 感官变化 文献 燕麦 固态 氨基酸、还原糖、多酚含量增加 色泽鲜艳,酱香浓郁,咸甜和粘稠适宜 [61]
糯米
固态总酚,35种特异性香味物质 α−淀粉酶、蛋白酶和β−葡萄糖苷酶活性增强;自由基清除能力和抗氧化能力皆提高,保护机体DNA免受氧化损伤的能力增强
香味更加丰富
[62−63]甜荞 固态 总酚(2.3倍),黄酮(2倍) 增强体外抗氧化性、自由基清除能力和糖脂代谢抑制率 [64−65] 苦荞 固态 清蛋白、球蛋白、醇蛋白含量显著提高 增强体外抗氧化性、自由基清除能力和糖脂代谢抑制率 [66] 黄豆 固态 总酚(1.94倍)、黄豆苷元(10.4倍)、金雀异黄酮(8.4倍)、总苷元(9.4倍);糖苷型黄酮含量降低 自由基清除能力显著提高 [67] 黑豆 固态 总酚、黄豆苷元、金雀异黄酮、大豆异黄酮含量提高;挥发性物质数量差异显著 抗氧化活性和自由基清除能力增强 香味提升 [68] OKARA 固态 大黄素和大黄素甲醚含量提高 α-糖苷酶抑制活性提高,降小鼠餐后血糖 [69] 5.3 中药材深加工
冠突散囊菌被用于中药材的深加工过程,可以有效促进中药的解毒和新型天然产物的产生,提高药效,见表3。人工接种冠突散囊菌发酵银杏叶或种子,可有效脱除银杏毒素和银杏酸,增加总黄酮、聚戊烯乙酸酯和芳香物质的含量[70-72];发酵茶“金花”茂盛,汤色橙黄明亮,滋味醇和,有金花香气[70];发酵物抗氧化能力提高,淀粉酶和蛋白酶活性增强[71-72]。葛根经冠突散囊菌固态发酵后,中药味得到显著改善,总黄酮和葛根素含量及抗氧化活性显著增加[73-74]。Xiang等[75]通过靶向代谢组学技术比较了冠突散囊菌和红曲霉发酵前后的姜黄素类化合物,共发现115种姜黄素类化合物,其中有48种结构比较新颖,14种为姜黄中首次发现,31种为发酵姜黄过程中新产生,其中的27种姜黄素类化合物为冠突散囊菌新发酵产生,23种为冠突散囊菌发酵特异性。为刺激其沉默的次级代谢产物合成基因簇的表达,陆承云[76]将冠突散囊菌与桑黄共培养并分别固体发酵金银花(叶)、葛根和黄芪。结果显示,发酵金银花(叶)的木犀草素含量增加,绿原酸、芦丁的含量显著降低,发酵黄芪的总黄酮含量显著提高,药效增加[76]。以保加利亚乳杆菌、凝结芽孢杆菌、鼠李糖乳杆菌和冠突散囊菌等食品工业中常用的微生物菌种液态发酵含有皂苷的三七、人参和西洋参几味中药,结果发现,仅冠突散囊菌能有效地将西洋参和高丽参中的其他皂苷成分转化为稀有人参皂苷Rd以及多种未知结构的高保留性皂苷产物[26]。冠突散囊菌发酵有效提高了枸杞[77]和白芷[78]的抗氧化活性以及自由基清除能力。以不同植物内生菌固态发酵“化症回生”口服液的残留药渣,发现来源于茯砖茶的冠突曲霉(CB10002)能有效回收利用药渣中的营养成分,并转化产生2.0 mg/g的羟基大黄素(citreorosein)和7.5 mg/g总蒽醌(anthraquinone)[79]。以冠突散囊菌转化中草药活性成分显示出良好的应用前景。
表 3 冠突散囊菌发酵不同中药材前后性状对比Table 3. Comparison of characteristics of different Chinese herbs before/after fermentation by Eurotium cristatum中药材 发酵菌种/状态 化合物变化 生物学活性 感官变化 文献 银杏叶 E.c/固态 提高:总黄酮(24.46%~64.9%));聚戊烯乙酸酯(10.6%);芳香物质;
降低:银杏酸(52.4%~55.63%);木质素(38.5%)和纤维素(20.1%)抗氧化活性、淀粉酶和
蛋白酶活性提高悦人香味 [70−71] 银杏籽粒 E.c (CCICC 2650)/液态 提高:洛伐他汀(32.97±0.17 μg mL−1);
降低:银杏毒素(40.15%),银杏酸(100%)抗氧化活性提高(2倍) [72] 葛根 E.c (KR812327,
HC-18)/固态提高:葛根素、总黄酮 抗氧化活性提高,提高药效 [73−74] 姜黄 E.c/固态 新产生27种姜黄素类化合物,23种菌特异性
姜黄素类化合物− [75] 黄芪 E.c与桑黄共发酵/固态 提高:总黄酮 提高药效 [76] 金银花/叶 E.c与桑黄共发酵/固态 增加:木犀草素
降低:绿原酸、芦丁[76] 西洋参/
高丽参E.c/液态 增加:Rd
降低:Rb1增加高活性稀有
皂苷的含量[26] 枸杞 E.c(CICC JH120502)/液态 蛋白质减少;
多酚、黄酮等先升高后降低抗氧化活性和自由基
清除能力提高[77] 白芷 E.c/固态 鉴定5种呋喃并香豆素、5种生物碱、2种聚酮 抗氧化活性提高,自由基
清除能力提高[78] 化症回生
药渣E.c(CB10002)/固态 产生羟基大黄素(2.0 mg/g)、
总蒽醌(7.5 mg/g)产生活性成分,减少
环境废物[79] 注:E.c为冠突散囊菌(Eurotium cristatum)的缩写。 5.4 其他应用
部分研究关注了对冠突散囊菌的活性代谢产物的开发。通过对液态发酵和提取的条件进行优化,胞外黑色素的产量可达4.27 g/L[29-30]。尹慧慧等考察以甜菜粉等为原料用冠突散囊菌发酵产洛伐他汀的最佳条件,得出最优组合的产量为146.4 mg/kg[80]。此外,将冠突散囊菌制成银纳米微粒(AgNP),对铜绿假单胞菌等多种菌有抑制效果,且表现出与新霉素等的协同抗菌效应[81],有潜力成为新型抗菌材料。茯砖茶来源的菌株ET1具有较强的降解β-氯氰菊酯和3-PBA的性能[82],可用于污染环境的生物修复。
6. 存在的问题及发展趋势
基于其强适应性、丰富的酶系、多样的活性代谢物和安全性等特性,冠突散囊菌在茶叶、豆谷类及中药材深加工等领域已得到较广泛应用。然而冠突散囊菌虽是潜在的益生菌,尚未被列入国家食品安全微生物名录;发酵过程多以固态为主,液态形式较少[83]。因此,该菌仍有较大的开发空间。代谢物方面,研究多集中于真菌转化基质总化合物库的变化分析,对仅来源于真菌菌丝或孢子的天然产物研究较少,对化合物转化的原因挖掘亦不透彻,可能是未来重要的研究方向。另外,多数基质经冠突散囊菌发酵后,其抗氧化性等生理学活性提高,但具体产生了哪些活性物质及如何发挥作用并不清楚。Zhou等通过在线HPLC-PDA-三项-TOF-MS/MS-ABTS系统,筛选并鉴定了冠突散囊菌发酵的白芷中发挥抗氧化作用的5种呋喃并香豆素、5种生物碱和2种聚酮[78],为此类研究提供了新的思路。综上,未来为使冠突散囊菌在食品加工、中药材加工等领域得到更好的应用,需综合运用多种技术手段在检测发酵过程,分析代谢物动态变化[46,74],锁定关键酶或相关生物代谢途径[16],探究菌体本身及其发酵制品的生理学活性及作用机制等方面展开深入细致地研究。
-
![]()
表 1 不同品种茶在冠突散囊菌发酵前后性状对比
Table 1 Comparison of characteristics of different tea varieties before/after fermentation by Eurotium cristatum
类型 茶品种 发酵条件 主成分变化 生物学活性 感官变化 文献 茶 茯砖茶 固态 提高:茶多糖和茶褐素;
降低:茶多酚、EGCG、游离氨基酸、
咖啡碱、茶黄素、茶红素香味提升,色泽更佳 [4-5] 普洱茶 固态 提高:总黄酮(19.15%),非酯型儿茶素(52.85%),酯类气体(29.78%)、醇类气体(20.84%);
降低:酯型儿茶素(33.64%)、总酚(22.31%)、
氨基酸(24.34%)苦涩味降低,香味提升 [48] 绿茶 固态/液态 降低:茶多酚 口感醇和、苦涩味降低;香味提升,兼具茶香、菌花香和甜香 [17,46,49] 红茶 液态 香味提升 [49] 复配藤茶 固态 提高:可溶性糖(4.65%)、咖啡碱(3.9%);
降低:多酚(49.68%)、氨基酸(56.79%)、二氢杨梅(39.29%)、儿茶素酸涩味降低、
口感提升[53] 白毫毛尖茶 固态 降低:茶多酚 降低:抗氧化、清除自由基能力 香味提升 [7] 植
物
代
茶蒲公英茶 固态 降低:多酚(19.76%) 香气协调、无异味 [57] 荔枝草茶 固态 提高:茶黄素(3.02倍)、茶红素(1.73倍)、茶褐素(1.29倍);
降低:茶多酚(31.9%)、氨基酸(36.3%)、可溶性糖(25.2%)苦涩味减少、
香味提升[58] 刺五加茶 固态 提高:茶褐素(18.64%);降低:茶多酚(66.75%)、氨基酸(69.05%)、可溶性糖(55.1%) 总抗氧化活性提高、对DPPH自由基
清除率下降苦涩味减少、
香味提升[56] 甜茶 固态 提高:茶多酚、黄酮;降低:游离氨基酸和可溶性糖 提高:抗氧化;降低:清除自由基能力 香味提升 [54] 桑叶茶 固态 提高:还原糖(10.47%)、总糖(22.36%)、氨基酸(22.87%) 营养、保健品质提升 [50−52] 苦丁茶 固态 苦味、粗青气下降;增加菌花香和甜香 [59] 杜仲茶 固态 降低:茶多酚、咖啡碱、氨基酸、可溶性糖 醇和、回甘、菌花香浓 [51] 栘㐿茶 固态 提高:根皮素;
降低:根皮苷抗氧化和抑菌
活性增强[60]  下载: 导出CSV
下载: 导出CSV
表 2 谷类和豆类经冠突散囊菌发酵前后性状对比
Table 2 Comparison of characteristics of different food varieties before/after fermentation by Eurotium cristatum
食材 发酵状态 化合物变化 生物学活性 感官变化 文献 燕麦 固态 氨基酸、还原糖、多酚含量增加 色泽鲜艳,酱香浓郁,咸甜和粘稠适宜 [61]
糯米
固态总酚,35种特异性香味物质 α−淀粉酶、蛋白酶和β−葡萄糖苷酶活性增强;自由基清除能力和抗氧化能力皆提高,保护机体DNA免受氧化损伤的能力增强
香味更加丰富
[62−63]甜荞 固态 总酚(2.3倍),黄酮(2倍) 增强体外抗氧化性、自由基清除能力和糖脂代谢抑制率 [64−65] 苦荞 固态 清蛋白、球蛋白、醇蛋白含量显著提高 增强体外抗氧化性、自由基清除能力和糖脂代谢抑制率 [66] 黄豆 固态 总酚(1.94倍)、黄豆苷元(10.4倍)、金雀异黄酮(8.4倍)、总苷元(9.4倍);糖苷型黄酮含量降低 自由基清除能力显著提高 [67] 黑豆 固态 总酚、黄豆苷元、金雀异黄酮、大豆异黄酮含量提高;挥发性物质数量差异显著 抗氧化活性和自由基清除能力增强 香味提升 [68] OKARA 固态 大黄素和大黄素甲醚含量提高 α-糖苷酶抑制活性提高,降小鼠餐后血糖 [69]
下载: 导出CSV
表 3 冠突散囊菌发酵不同中药材前后性状对比
Table 3 Comparison of characteristics of different Chinese herbs before/after fermentation by Eurotium cristatum
中药材 发酵菌种/状态 化合物变化 生物学活性 感官变化 文献 银杏叶 E.c/固态 提高:总黄酮(24.46%~64.9%));聚戊烯乙酸酯(10.6%);芳香物质;
降低:银杏酸(52.4%~55.63%);木质素(38.5%)和纤维素(20.1%)抗氧化活性、淀粉酶和
蛋白酶活性提高悦人香味 [70−71] 银杏籽粒 E.c (CCICC 2650)/液态 提高:洛伐他汀(32.97±0.17 μg mL−1);
降低:银杏毒素(40.15%),银杏酸(100%)抗氧化活性提高(2倍) [72] 葛根 E.c (KR812327,
HC-18)/固态提高:葛根素、总黄酮 抗氧化活性提高,提高药效 [73−74] 姜黄 E.c/固态 新产生27种姜黄素类化合物,23种菌特异性
姜黄素类化合物− [75] 黄芪 E.c与桑黄共发酵/固态 提高:总黄酮 提高药效 [76] 金银花/叶 E.c与桑黄共发酵/固态 增加:木犀草素
降低:绿原酸、芦丁[76] 西洋参/
高丽参E.c/液态 增加:Rd
降低:Rb1增加高活性稀有
皂苷的含量[26] 枸杞 E.c(CICC JH120502)/液态 蛋白质减少;
多酚、黄酮等先升高后降低抗氧化活性和自由基
清除能力提高[77] 白芷 E.c/固态 鉴定5种呋喃并香豆素、5种生物碱、2种聚酮 抗氧化活性提高,自由基
清除能力提高[78] 化症回生
药渣E.c(CB10002)/固态 产生羟基大黄素(2.0 mg/g)、
总蒽醌(7.5 mg/g)产生活性成分,减少
环境废物[79] 注:E.c为冠突散囊菌(Eurotium cristatum)的缩写。
下载: 导出CSV
-
[1] 李佳莲, 胡博涵, 刘素纯, 等. 微生物与茯砖茶品质形成研究进展[J]. 食品工业科技,2010,31(9):406−408,413. [LI J L, HU B H, LIU S C, et al. Research progress of microorganism and the formation of Fuzhuan tea quality[J]. Science and Technology of Food Industry,2010,31(9):406−408,413. [2] LIN Q, JIN Y L, JIANG R G, et al. Dynamic changes in the metabolite profile and taste characteristics of Fu brick tea during the manufacturing process[J]. Food Chemistry,2021,344:128576. doi: 10.1016/j.foodchem.2020.128576
[3] 王亚丽, 秦俊哲, 黄亚亚, 等. 冠突散囊菌对茯砖茶品质形成的影响[J]. 食品与发酵工业,2018,44(4):194−197. [WANG Y L, QIN J Z, HUANG Y Y, et al. Effect of Eurotium cristatum on quality formation of Fuzhuan tea[J]. Food and Fermentation Industries,2018,44(4):194−197. [4] XIAO Y, ZHONG K, BAI J, et al. Insight into effects of isolated Eurotium cristatum from Pingwu Fuzhuan brick tea on the fermentation process and quality characteristics of Fuzhuan brick tea[J]. Journal of the Science of Food and Agriculture,2020,100(9):3598−3607. doi: 10.1002/jsfa.10353
[5] ZHU M Z, LI N, ZHOU F, et al. Microbial bioconversion of the chemical components in dark tea[J]. Food Chemistry,2020,312:126043. doi: 10.1016/j.foodchem.2019.126043
[6] 中国国家标准化管理委员会. GB/T 9833.3-2013 紧压茶 第3部分: 茯砖茶[S]. 北京: 中国标准出版社, 2013. China National Standardization Administration Committee. GB/T 9833.3-2013 Compressed tea - Part 3: Fu zhuan tea[S]. Beijing: China Standards Press, 2013.
[7] 陈皓睿, 孙强, 闻东明, 等. 三株散囊菌发酵茶叶的初步研究[J]. 工业微生物,2019,49(1):21−25. [CHEN H R, SUN Q, WEN D M, et al. Preliminary study on three strains of the fermentation of tea by the three strains of bursa spp doi: 10.3969/j.issn.1001-6678.2019.01.004 J]. Industrial microorganism,2019,49(1):21−25. doi: 10.3969/j.issn.1001-6678.2019.01.004
[8] 温琼英. 茯砖茶中优势菌的种名鉴定[J]. 中国茶叶,1990,6:2−3. [WEN Q Y. Species identification of dominant bacteria in Fuzhuan tea[J]. Chinese Tea,1990,6:2−3. [9] HUKBA V, KOLARIK M, KUBATOVA A, et al. Taxonomic revision of Eurotium and transfer of species to Aspergillus[J]. Mycologia,2013,105(4):912−937. doi: 10.3852/12-151
[10] GE Y Y, WANG Y C, LIU Y X, et al. Comparative genomic and transcriptomic analyses of the Fuzhuan brick tea-fermentation fungus Aspergillus cristatus[J]. BMC Genomics,2016,17(1):428. doi: 10.1186/s12864-016-2637-y
[11] KANG D, SU M, DUAN Y, et al. Eurotium cristatum, a potential probiotic fungus from Fuzhuan brick tea, alleviated obesity in mice by modulating gut microbiota[J]. Food Function,2019,10(8):5032−5045. doi: 10.1039/C9FO00604D
[12] REN X X, WANG Y, LIU Y X, et al. Comparative transcriptome analysis of the calcium signaling and expression analysis of sodium/calcium exchanger inAspergillus cristatus[J]. Journal of Basic Microbiology,2018,58(1):76−87. doi: 10.1002/jobm.201700277
[13] LIU H, SANG S L, WANG H, et al. Comparative proteomic analysis reveals the regulatory network of the veA gene during asexual and sexual spore development of Aspergillus cristatus[J]. Bioscience Reports,2018,38(4):BSR20180067. doi: 10.1042/BSR20180067
[14] 尹旭敏. 四川“金花”菌生物学特性及其发酵液态茶工艺的初步研究[D]. 成都: 四川农业大学, 2006. YIN X M. Preliminary study on biological characteristics of Sichuan Jinhua fungus and its fermentation technology of liquid tea[D]. Chengdu: Sichuan Agricultural University, 2006.
[15] 郑欣欣. 茯砖茶中“金花”菌产孢机制及其功能性的研究[D]. 西安: 陕西科技大学, 2015. ZHENG X X. Studies on sporulation mechanism and function of “Jinhua” fungus in Fuzhuan Tea [D]. Xi’an: Shaanxi University of science and technology, 2015.
[16] 刘作易, 秦京. 茯砖茶“金花”菌对营养成分的利用[J]. 贵州农业科学,1992(1):36−40. [LIU Z Y, QIN J. Utilization of nutrients by “Jinhua” fungus of Fuzhuan tea[J]. Guizhou Agricultural Sciences,1992(1):36−40. [17] XIAO Y, LI M Y, LIU Y, et al. The effect of Eurotium cristatum (MF800948) fermentation on the quality of autumn green tea[J]. Food Chemistry,2021,358:129848. doi: 10.1016/j.foodchem.2021.129848
[18] 蔡正安, 刘素纯, 刘仲华, 等. 茯砖茶中冠突散囊菌纤维素酶的酶学性质研究[J]. 茶叶科学,2010,30(1):57−62. [CAI Z A, LIU S C, LIU Z H, et al. Cellulase enzymatic property of Eurotium Cristatum from brick tea[J]. Tea Science,2010,30(1):57−62. doi: 10.3969/j.issn.1000-369X.2010.01.009 [19] 雷林超. 冠突散囊菌产阿魏酸酯酶及其在黑茶制备中的应用[D]. 泉州: 华侨大学, 2020. LEI L C. Production of ferulic acid esterase by Eurotium cristatum and its application in black tea preparation[D]. Quanzhou: Huaqiao University, 2020.
[20] 黄建安, 刘仲华, 施兆鹏. 茯砖茶制造中主要酶类的变化[J]. 茶叶科学,1991(增刊ement):63−68. [HUANG J A, LIU Z H, SHI Z P. Changes of main enzymes in Fuzhuan tea production[J]. Tea Science,1991(Supplement):63−68. [21] 刘仲华, 黄建安, 王增盛, 等. 茯砖茶加工中色素物质的变化与色泽品质的形成[J]. 茶叶科学,1991(增刊ement):76−78. [LIU Z H, HUANG J A, WANG Z S, et al. The change of pigment substance and the formation of color quality in Fuzhuan tea processing[J]. Tea Science,1991(Supplement):76−78. [22] WANG Q, GONG J, CHISTI Y, et al. Fungal isolates from a Pu-erh type tea fermentation and their ability to convert tea polyphenols to theabrownins[J]. Journal of Food Science,2015,80(4):M809−M817. doi: 10.1111/1750-3841.12831
[23] 谢晓晨, 唐万达, 万晴, 等. 不同发酵茶中优势真菌的抗氧化酶活性[J]. 中国微生态学杂志,2020,32(6):637−641,655. [XIE X C, TANG W D, WAN Q, et al. Antioxidant enzyme activities of dominant fungi in different fermented teas[J]. Chinese Journal of Microecology,2020,32(6):637−641,655. [24] 丁婷. 茯砖茶中“金花菌”的生物学特性及其产消化酶活性的研究[D]. 西安: 陕西科技大学, 2012. DING T. Study on biological characteristics and digestive enzyme activity of "Jinhua fungus" in Fuzhuan tea[D]. Xi’an: Shaanxi University of Science and Technology, 2012.
[25] 吴志超. 黑茶发酵优势菌株的胞外酶检测及蛋白组学初探[D]. 福州: 福建师范大学, 2018. WU Z C. Detection of extracellular enzymes and preliminary proteomics of dominant strains for black tea fermentation[D]. Fuzhou: Fujian Normal University, 2018.
[26] 李粟琳, 张翔宇, 王洋, 等. 可发酵三七等中药材的食用菌种筛选和皂苷生物转化产物的分析[J]. 食品与发酵工业,2017,43(12):164−168. [LI S L, ZHANG X Y, WANG Y, et al. Screening of edible fungi for fermentation of Panax notoginseng Chinese medicinal herbs and analysis of biotransformation products of saponins[J]. Food and Fermentation Industries,2017,43(12):164−168. [27] 郑欣欣. 茯砖茶中“金花”菌产孢机制及其功能性研究[D]. 西安: 陕西科技大学, 2015. ZHENG X X. Sporulation mechanism and functional study of "Golden Flower" bacteria in Poria brick tea[D]. Xi’an: Shaanxi University of Science and Technology, 2015.
[28] 李文娟, 吕嘉枥, 杜冰, 等. 冠突散囊菌液体发酵产色素理化性质的研究[J]. 食品工业,2016,37(1):27−31. [LI W J, LV J L, DU B, et al. Study on properties of pigment produced during the liquid fermentation of Eurotium cristatum[J]. Food Industry,2016,37(1):27−31. [29] 韩蓉, 吕嘉枥, 王珊. 超声波作用对茯砖茶金花菌色素提取率的影响[J]. 食品科技,2014,39(1):240−243. [HAN R, LV J L, WANG S. The effect of ultrasonic on the extraction rate of pigment from Fu brick tea[J]. Food Science and Technology,2014,39(1):240−243. [30] 杨妮, 刘素纯, 王继刚, 等. 冠突散囊菌产胞外黑色素发酵条件优化及稳定性研究[J]. 食品与发酵工业,2020,46(16):37−42. [YANG N, LIU S C, WANG J G, et al. Optimization of fermentation conditions and stability of extracellular melanin production by Eurotium cristatum[J]. Food and Fermentation Industries,2020,46(16):37−42. [31] 杨妮, 刘素纯, 王继刚, 等. 冠突散囊菌胞外黑色素碱法提取工艺的研究[J]. 中国酿造,2020,39(3):63−67. [YANG N, LIU S C, WANG J G, et al. Study on alkaline extraction technology of extracellular melanin from Eurotium cristatum[J]. China Brewing,2020,39(3):63−67. doi: 10.11882/j.issn.0254-5071.2020.03.013 [32] LIU G, DUAN Z G, WANG P, et al. Purification, characterization, and hypoglycemic properties of eurocristatine from Eurotium cristatum spores in Fuzhuan brick tea[J]. RSC Advances,2020,10:22234. doi: 10.1039/D0RA03423A
[33] ZHANG H, HUI J F, YANG J, et al. Eurocristatine, a plant alkaloid from Eurotium cristatum, alleviates insulin resistance in db/db diabetic mice via activation of PI3K/AKT signaling pathway[J]. European Journal of Pharmacology,2020,887:173557. doi: 10.1016/j.ejphar.2020.173557
[34] SHI J, LIU J, KANG D, et al. Isolation and characterization of benzaldehyde derivatives with anti-inflammatory activities from Eurotium cristatum, the dominant fungi species in Fuzhuan brick tea[J]. ACS Omega,2019,4(4):6630−6636. doi: 10.1021/acsomega.9b00593
[35] ZOU X W, LI Y, ZHANG X N, et al. A new prenylated indole diketopiperazine alkaloid from Eurotium cristatum[J]. Molecules,2014,19:17839−17847. doi: 10.3390/molecules191117839
[36] XIE Z, BAI Y, CHEN G, et al. Modulation of gut homeostasis by exopolysaccharides from Aspergillus cristatus (MK346334), a strain of fungus isolated from Fuzhuan brick tea, contributes to immunomodulatory activity in cyclophosphamide-treated mice[J]. Food and Function,2020,11(12):10397−10412. doi: 10.1039/D0FO02272A
[37] LI H H, MI Y, DUAN Z G, et al. Structural characterization and immunomodulatory activity of a polysaccharide from Eurotium cristatum[J]. International Journal of Biological Macromolecules,2020,162:609−617. doi: 10.1016/j.ijbiomac.2020.06.099
[38] 王珊. 茯砖茶中降胆固醇类物质的研究[D]. 西安: 陕西科技大学, 2014. WANG S. Study on cholesterol lowering substances in Fu brick tea[D]. Xi’an: Shaanxi University of Science and Technology, 2014.
[39] 杨娟娟, 邹先伟, 黄昀, 等. 冠突散囊菌与金黄色葡萄球菌混合发酵次级代谢产物及细胞毒活性[J]. 科技导报, 2016, 34(9): 96−101. YANG J J, ZOU X W, HUANG J, et al. Secondary metabolites and cytotoxic activity of mixed fermentation of Eurotium cristatum and Staphylococcus aureus[J], Science and Technology Guide, 2016, 34(9): 96−101.
[40] 苏凤. 冠突散囊菌接种发酵茯砖茶的初步研究及其安全性评价[D]. 武汉: 武汉工业学院, 2011. SU F. Preliminary study on the fermentation of Fuzhuan tea inoculated with Eurotium cristatum and its safety evaluation[D]. Wuhan: Wuhan Institute of Technology, 2011.
[41] 景悦, 路晓杰, 曹永国, 等. 冠突散囊菌对溃疡性结肠炎模型小鼠的保护作用及机制[J]. 中国兽医学报,2021,41(4):683−688,784. [JING Y, LU X J, CAO Y G, et al. Protective effect and mechanism of Eurotium cristatum on ulcerative colitis in mice[J]. Chinese Journal of veterinary medicine,2021,41(4):683−688,784. [42] 覃金球, 罗美玲, 张旋, 等. 冠突散囊菌医药价值研究进展[J]. 食品工业科技,2018,39(24):336−339. [TAN J Q, LUO M L, ZHANG X, et al. Research progress on medical value of Eurotium cristatum[J]. Science and Technology of Food Industry,2018,39(24):336−339. [43] ZHOU C B, ZHOU X L, WEN Z R, et al. Compound Fu brick tea modifies the intestinal microbiome composition in high-fat diet-induced obesity mice[J]. Food Science and Nutrition,2020,8(10):5508−5520. doi: 10.1002/fsn3.1850
[44] LIU D M, HUANG J A, LUO Y, et al. Fuzhuan brick tea attenuates high-fat diet-induced obesity and associated metabolic disorders by shaping gut microbiota[J]. Journal of Agricultural and Food Chemistry,2019,67(49):13589−13604. doi: 10.1021/acs.jafc.9b05833
[45] FU D H, RYAN E P, HUANG J N, et al. Fermented Camellia sinensis, Fuzhuan tea, regulates hyperlipidemia and transcription factors involved in lipid catabolism[J]. Food Research International,2011,44(9):2999−3005. doi: 10.1016/j.foodres.2011.07.008
[46] XIAO Y, ZHONG K, BAI J R, et al. The biochemical characteristics of a novel fermented loose tea by Eurotium cristatum (MF800948) and its hypolipidemic activity in a zebrafish model[J]. LWT,2020,117:108629. doi: 10.1016/j.lwt.2019.108629
[47] CHEN Q, ZHANG M Y, CHEN M X, et al. Influence of Eurotium cristatum and Aspergillus niger individual and collaborative inoculation on volatile profile in liquid-state fermentation of instant dark teas[J]. Food Chemistry,2021,350:129234−129234. doi: 10.1016/j.foodchem.2021.129234
[48] JIANG C K, ZHEN Z G, HUANG Y H, et al. Chemical compositions of Pu'er tea fermented by Eurotium cristatum and their lipid-lowering activity[J]. LWT-Food Science and Technology,2018,98:204−211. doi: 10.1016/j.lwt.2018.08.007
[49] 郑梦霞. 冠突散囊菌发酵对绿茶及红茶提取液香气成分的影响[D]. 南京: 南京农业大学, 2016. ZHENG M X. Effect of fermentation by Eurotium cristatum on aroma components of green tea and black tea extracts[D]. Nanjing: Nanjing Agricultural University, 2016.
[50] 梁贵秋, 谢婷婷, 高艳芳, 等. 3株冠突散囊菌(Eurotium cristatum)发酵对桑叶茶品质的影响[J]. 蚕业科学,2020,46(5):614−621. [LIANG G Q, XIE T T, GAO Y F, et al. Effect of Eurotium cristatum fermentation on the quality of mulberry leaf tea[J]. Science of Sericulture,2020,46(5):614−621. [51] 杨乔. 杜仲叶和桑叶两种保健功能型茯茶加工工艺研究[D]. 咸阳: 西北农林科技大学, 2019. YANG Q. Study on the processing technology of two kinds of health functional Fu tea with eucommia leaves and mulberry leaves[D]. Xianyang: Northwest University of Agriculture and Forestry Science and Technology, 2019.
[52] LI F M, LI Y P, SHAO Y Y, et al. Effect of different pile fermentation time on the blooming effect and sensory quality of compound mulberry leaf Fuzhuan brick tea[J]. China Sericulture,2016,37(3):7−11,16.
[53] 赵勇彪. 发酵藤茶生产技术及其风味形成机制研究[D]. 吉首: 吉首大学, 2013. ZHAO Y B. Production technology and flavor formation mechanism of fermented Ampelopsis grossedentata[D]. Jishou: Jishou University, 2013.
[54] 朱雯, 马煜明, 曾莉, 等. 金花菌发酵甜茶生化品质及抗氧化活性分析[J]. 茶叶通讯,2019,46(3):318−322. [ZHU W, MA Y M, ZENG L, et al. Analysis of biochemical quality and antioxidant activity of sweet tea fermented by Jinhua fungus[J]. Tea Communication,2019,46(3):318−322. doi: 10.3969/j.issn.1009-525X.2019.03.013 [55] 欧阳梅, 熊昌云, 屠幼英, 等. 冠突散囊菌对茶叶品质成分及其抗氧化活性影响[J]. 菌物学报,2011,30(2):343−348. [OUYANG M, XIONG C Y, TU Y Y, et al. Effect of Eurotium cristatum on quality components and antioxidant activity of tea[J]. Acta Mycologica Sinica,2011,30(2):343−348. [56] 杨立娜, 吴凯为, 林杉, 等. 基于感官评价的刺五加茶正交优化工艺研究[J]. 渤海大学学报(自然科学版),2019,40(1):11−18. [YANG L N, WU K W, LIN S, et al. Orthogonal optimization of Acanthopanax senticosus tea based on sensory evaluation[J]. Journal of Bohai University (Natural Science Edition),2019,40(1):11−18. [57] 丁磊, 李鑫洋, 刘佳鑫, 等. 蒲公英发酵茶工艺研究[J]. 食品研究与开发,2020,41(15):75−79. [DING L, LI X Y, LIU J X, et al. Study on the fermentation technology of dandelion tea[J]. Food Research and Development,2020,41(15):75−79. doi: 10.12161/j.issn.1005-6521.2020.15.014 [58] 杨立娜, 吴凯为, 徐清莹, 等. 冠突散囊菌发酵对荔枝草茶主要成分及风味的影响[J]. 食品与发酵工业,2019,45(13):121−125. [YANG L, WU K W, XU Q Y, et al. Effect of fermentation by Eurotium cristatum on main components and flavor of litchi grass tea[J]. Food and Fermentation Industries,2019,45(13):121−125. [59] 陈敏, 谢发, 游玲, 等. 冠突散囊菌发酵苦丁茶工艺研究[J]. 食品与发酵工业,2020,46(6):224−228. [CHEN M, XIE F, YOU L, et al. Fermentation of Ligustrum robustum by Eurotium cristatum[J]. Food and Fermentation Industries,2020,46(6):224−228. [60] XIE J, CHEN J, MEI X R, et al. Biotransformation of phlorizin by Eurotium cristatum to increase the antioxidant and antibacterial activity of Docynia indica leaves[J]. Current microbiology,2021,78(4):1590−1601. doi: 10.1007/s00284-021-02366-3
[61] 唐梓尧, 刘素纯, 林辉, 等. 冠突散囊菌发酵制备燕麦面酱工艺研究[J]. 中国酿造,2019,38(2):117−121. [TANG Z Y, LIU S C, LIN H, et al. Preparation technology of oat flour paste fermented by Eurotium cristatum[J]. China Brewing,2019,38(2):117−121. doi: 10.11882/j.issn.0254-5071.2019.02.022 [62] XU X, ZHOU S D, MCCLEMENTS D J, et al. Multistarter fermentation of glutinous rice with Fu brick tea: Effects on microbial, chemical, and volatile compositions[J]. Food Chemistry,2020,309:125790. doi: 10.1016/j.foodchem.2019.125790
[63] XU X, SHEN C, YUAN B, et al. Glutinous rice fermented with Fu brick tea: Effects on phenolic content, antioxidant activities and DNA damage protection[J]. FASEB Journal,2020,34(S1):1−1.
[64] 黄俊. 冠突散囊菌发酵甜荞主要功能性成分变化和功效研究[D]. 长沙: 湖南农业大学, 2019. HUANG J. Study on the change and efficacy of main functional components of Fagopyrum esculentum fermented by Eurotium cristatum[D]. Changsha: Hunan Agricultural University, 2019.
[65] XIAO Y, WU X, YAO X S, et al. Metabolite profiling, antioxidant and α-glucosidase inhibitory activities of buckwheat processed by solid-state fermentation with Eurotium cristatum YL-1[J]. Food Research International,2021,143:110262. doi: 10.1016/j.foodres.2021.110262
[66] 兰晓勇, 刘素纯, 李再贵. 冠突散囊菌不同发酵条件对苦荞蛋白质的影响[J]. 中国酿造,2019,38(10):72−76. [LAN X Y, LIU S C, LI Z G. Effects of different fermentation conditions on tartary buckwheat protein[J]. China Brewing,2019,38(10):72−76. doi: 10.11882/j.issn.0254-5071.2019.10.015 [67] CHEN Y, WANG Y, CHEN J, et al. Bioprocessing of soybeans (Glycine max L. ) by solid-state fermentation with Eurotium cristatum YL-1 improves total phenolic content, isoflavone aglycones, and antioxidant activity[J]. RSC Advances,2020,10(29):16928−16941. doi: 10.1039/C9RA10344A
[68] XIAO Y, HUANG Y X, CHEN Y L, et al. Effects of solid-state fermentation with Eurotium cristatum YL-1 on the nutritional value, total phenolics, isoflavones, antioxidant activity, and volatile organic compounds of black soybeans[J]. Agronomy,2021,11:1029. doi: 10.3390/agronomy11061029
[69] CHAN L Y, TAKAHASHI M, LIM P J, et al. Eurotium Cristatum fermented Okara as a potential food ingredient to combat diabetes[J]. Scientific Reports,2019,9(1):17536. doi: 10.1038/s41598-019-54021-4
[70] 秦俊哲, 任金玫, 李银亮, 等. 银杏茶人工接种发酵工艺优化研究[J]. 食品科技,2014,39(6):114−118. [QIN J Z, REN J M, LI Y L, et al. Optimization of fermentation process of Ginkgo biloba tea by artificial inoculation[J]. Food Science and Technology,2014,39(6):114−118. [71] ZOU M, GUO X, HUANG Y, et al. Improvement of the quality of Ginkgo biloba leaves fermented by Eurotium cristatum as high value-added feed[J]. Processes,2019,7(9):627. doi: 10.3390/pr7090627
[72] ZOU M M, ZHANG W, DONG Q H, et al. Submerged fermentation of Ginkgo biloba seed powder using Eurotium cristatum for the development of ginkgo seeds fermented products[J]. Journal of the Science of Food and Agriculture,2021,101(5):1782−1791.
[73] 胡谢馨, 易有金, 柏连阳, 等. 茯砖茶“金花”菌的分离、鉴定及转化葛根产物研究[J]. 茶叶科学,2016,36(3):268−276. [HU X X, YI Y J, BAI L Y, et al. Isolation, identification and transformation of Pueraria lobata from Fuzhuan tea[J]. Tea Science,2016,36(3):268−276. doi: 10.3969/j.issn.1000-369X.2016.03.007 [74] ZHANG B, ZHOU L, XIAO X, et al. Solid-state fermentation with Eurotium cristatum HC-18 to improve antioxidant activity of kudzu (Pueraria lobata) root[J]. Journal of Food and Nutrition Research,2018,57(4):384−395.
[75] XIANG X, SONG C, SHI Q, et al. A novel predict-verify strategy for targeted metabolomics: Comparison of the curcuminoids between crude and fermented turmeric[J]. Food Chemistry,2020,331:127281. doi: 10.1016/j.foodchem.2020.127281
[76] 陆承云. 两种药用真菌新化合物发现及其对中药材的转化作用[D]. 贵阳: 贵州师范大学, 2018. LU C Y. Discovery of two new compounds of medicinal fungi and their transformation effect on traditional Chinese medicine[D]. Guiyang: Guizhou Normal University, 2018.
[77] 李鹏程, 肖咪, 薛宏坤, 等. 冠突散囊菌发酵枸杞工艺优化及其抗氧化活性研究[J]. 食品科技,2021,46(2):51−58. [LI P C, XIAO M, XUE H K, et al. Optimization of fermentation technology and antioxidant activity of Lycium barbarum by Eurotium cristatum[J]. Food Technology,2021,46(2):51−58. [78] ZHOU S D, XU X, LIN Y F, et al. On-line screening and identification of free radical scavenging compounds in Angelica dahurica fermented with Eurotium cristatum using an HPLC-PDA-Triple-TOF-MS/MS-ABTS system[J]. Food Chemistry,2019,272:670−678. doi: 10.1016/j.foodchem.2018.07.173
[79] KONG W, HUANG C, SHI J, et al. Recycling of Chinese herb residues by endophytic and probiotic fungus Aspergillus cristatus CB10002 for the production of medicinal valuable anthraquinones[J]. Microbial Cell Factories,2019,18(1):102. doi: 10.1186/s12934-019-1150-9
[80] 尹慧慧, 陈育如, 赵文文. 冠突散囊菌利用不同原料产洛伐他汀的条件优化[J]. 南京师大学报(自然科学版),2018,41(2):93−98. [YIN H H, CHEN Y R, ZHAO W W. Optimization of conditions for producing lovastatin by Eurotium cristatum from different raw materials[J]. Journal of Nanjing Normal University (Natural Science Edition),2018,41(2):93−98. [81] LIN P, WANG F Q, LI C T, et al. An enhancement of antibacterial activity and synergistic effect of biosynthesized silver nanoparticles by Eurotium cristatum with various antibiotics[J]. Biotechnology and Bioprocess Engineering,2020,25(3):450−458. doi: 10.1007/s12257-019-0506-7
[82] HU K D, DENG W Q, ZHU Y T, et al. Simultaneous degradation of beta-cypermethrin and 3-phenoxybenzoic acid by Eurotium cristatum ET1, a novel “golden flower fungus” strain isolated from Fu brick tea[J]. Microbiology open,2019,8(7):e776.
[83] 徐小倩. 液态茶饮料的混菌发酵工艺研究[D]. 合肥: 安徽农业大学, 2016. XU X Q. Study on mixed fermentation technology of liquid tea beverage[D]. Hefei: Anhui Agricultural University, 2016.
-
期刊类型引用(13)
1. 谢耀慧,李成义,冯慧敏,贾妙婷,魏小成. 中药活性成分微生物发酵研究进展. 中成药. 2025(01): 182-187 .  百度学术
百度学术
2. 李萌,曾婷婷,杨金初,徐永明,李欢,马林,饶智. 冠突散囊菌GT-1固态发酵对低次烟叶品质的影响. 陕西科技大学学报. 2024(03): 38-45 . 百度学术
3. 赵千慧,周慧玲,秦洋. 冠突散囊菌对高粱酒酿造过程糖化效率的促进作用与原理. 食品与发酵工业. 2024(15): 72-78 . 百度学术
4. 吴云迪,刘谢缘,李利君,倪辉. 过氧化氢酶对白芽奇兰茶叶香气成分转化的影响. 食品科学. 2024(22): 164-172 . 百度学术
5. 王莹,贾敏,程文婷,袁亚娟. 不同均质时间和稀释液对茯砖茶中冠突散囊菌计数的影响研究. 中外食品工业. 2024(14): 37-39 . 百度学术
6. 谢宁轩,张文刚,党斌,郑万财,杨希娟. 冠突散囊菌发酵对青稞多酚组成及其体外抗氧化及糖脂代谢酶抑制活性的影响. 食品与发酵工业. 2024(23): 87-94 . 百度学术
7. 冯璞阳,白亚妮,冯志珍,秦涛,陈卫锋. 冠突散囊菌散茶发花工艺提高茯茶品质研究进展. 茶叶. 2023(01): 9-12 . 百度学术
8. 尹家全,刘歆旖,王兰,刘素纯. 冠突散囊菌发酵罗汉果渣过程中功能性成分及抗氧化活性的变化. 现代食品科技. 2023(04): 88-95 . 百度学术
9. 肖咪,刘利敏,李鹏程,薛宏坤,邹先伟,唐劲天,苏彦奇,蔡旭. 冠突散囊菌发酵湖北海棠茶过程中主要成分及活性研究. 食品科技. 2022(01): 79-86 . 百度学术
10. 吉巧琳,陈鑫,莫海英,蔡国磊,杨海英,杜刚. 茯茶中冠突曲霉的分离及比较基因组研究. 食品科技. 2022(05): 21-26 . 百度学术
11. 刘璐,袁亚宏,岳田利. 葛根降血压茶的制备及对自发性高血压大鼠的降压作用. 食品与机械. 2022(07): 213-219 . 百度学术
12. 党旭辉,周秦羽,刘梦圆,甘汉文,张犇,刘锦,李敏,覃丽,肖文军. 黑茶金花体外抗氧化及降血脂活性研究. 食品安全质量检测学报. 2022(24): 7927-7933 . 百度学术
13. 赵敏晶. 茯茶品牌的发展路径探索. 炎黄地理. 2022(11): 53-55 . 百度学术
其他类型引用(10)
 下载:
下载:

计量
- 文章访问数: 1123
- HTML全文浏览量: 1057
- PDF下载量: 131
- 被引次数: 23



