


Screening and Identification of Alginate Lyase-producing Strains and Optimization of Its Enzyme-production Conditions
-
摘要: 以海藻酸钠为唯一碳源培养基,对广西北海涠洲岛采集的马尾藻进行微生物的筛选和分离,采用 3, 5-二硝基水杨酸法 (DNS)进行海藻酸钠裂解酶相对酶活力测定,获得高产酶菌株M0101;依据形态、生理生化特征及 16S rDNA 序列分析对其进行鉴定,该菌属于弧菌属,命名为Vibrio gangliei M0101;通过单因素实验确定了菌株M0101的产酶优化方案:海藻酸钠50 g/L,(NH4)2SO4 2 g/L,K2HPO4 1 g/L,NaCl 100 g/L,MgSO4·7H2O 1 g/L,FeSO4·7H2O 0.01 g/L,培养温度30 ℃,初始pH9.0,200 r/min;优化后菌株 M0101产酶最大活性可达221.90 U/mL,相较于优化前最大酶活36.32 U/mL,优化后酶活力是优化前的6.11倍。表明M0101能高效水解高浓度海藻酸钠,是强耐盐的菌株,具有大规模制备海藻酸钠裂解酶的潜力。Abstract: Using sodium alginate as the sole carbon source medium, the microorganisms in Sargassum collected from Weizhou Island, Guangxi, were screened and isolated. The relative enzyme activity was determined by 3,5-Dinitrosalicylic acid (DNS) method, and the high-yield strain, marked M0101, was obtained. According to the morphological, physiological and biochemical characteristics and 16S rDNA sequence analysis, the target strain was identified to belong to the genus of Vibrio, named as Vibrio gangliei M0101. The prioritization scheme of enzyme production of M0101 was confirmed by a single factor test: Alginate 50 g/L, (NH4)2SO4 2 g/L, K2HPO4 1 g/L, NaCl 100 g/L, MgSO4·7H2O 1 g/L, FeSO4·7H2O 0.01 g/L, 30 ℃, 200 r/min, initial pH9.0; and the maximum enzyme activity of strain M0101 reached 221.90 U/mL after optimation. It was 6.11 times higher than before(36.32 U/mL). Its efficiently hydrolysis of high concentration alginate and high salt tolerance, which has potential for large-scale preparation of alginate lyase.
-
褐藻胶是巨藻、墨角藻、马尾藻和海带等褐藻细胞壁和细胞间质中的一类水溶性酸性多糖物质,也是褐藻中含量最多的多糖,其在地球上的含量仅次于纤维素,可作为潜在的生物质能源原料。然而褐藻胶高粘、高凝胶、不易降解、难以被机体吸收利用的特点,限制了它的应用[1],褐藻胶的降解产物海藻酸寡糖在抗肿瘤、抗凝血、抗菌、抗病毒、增强宿主免疫力、抗氧化、神经保护、肠道消化吸收、植物的生长及保鲜等方面有良好的生物活性,被广泛应用于食品、医疗及农业等领域[2-5]。传统制备褐藻胶寡糖的方法主要有物理降解法、化学降解法和生物酶解法[6]等,其中生物酶解法因其专一性高、反应条件温和、易控制、目标产物得率高等优点而备受关注。
褐藻胶裂解酶通过β-消除反应将褐藻胶降解为一系列不饱和的海藻酸寡糖,根据底物专一性不同将褐藻胶裂解酶分为三类:转移性裂解1,4-α-D-甘露糖醛酸片段的裂解酶(Aly;EC4.2.2.3),专一性裂解1,4-α-L-古罗糖醛酸片段的裂解酶(Aly;EC4.2.2.11)以及两种片段都可裂解的双功能裂解酶[7]。褐藻胶裂解酶来源广泛,主要来源包括藻类、动物内脏和微生物等,其中海洋微生物是褐藻胶裂解酶的重要来源之一,目前的研究表明来自于海洋动物的微生物产生的裂解海藻酸钠的酶多数为复合酶,很难分离纯化,如假交替单胞菌Pseudoalteromonas elyakovii IAM 14954 至少能产生五种不同的海藻酸钠裂解酶,其中一个是胞外酶,四个是胞内酶[8]。现有大部分微生物来源的褐藻胶裂解酶依然存在着酶活力较低、作用位点单一等缺陷,造成酶解法制备海藻提取物的成本过高,且没有统一的发酵工程菌及系统发酵条件,褐藻胶裂解酶稳定性差,达不到工业发酵生产要求[9],发展受限。不同来源的海藻酸钠降解酶所获得的产物聚合度也会有所不同,进而对寡糖的生物活性产生影响。从不同的途径寻找和生产具有高活性的海藻酸钠裂解酶已成为研究焦点[10]。筛选高效降解褐藻胶菌株是开发褐藻资源的重要新途径,目前鲜少有报道在50 g/L高浓度海藻酸钠培养基中仍保持酶活力较强的菌株,近年来多数报道的降解菌直接利用褐藻胶的浓度低于50 g/L[11-13]。
本研究从北海涠洲岛采集的马尾藻中筛选得到一株褐藻胶降解菌Vibrio gangliei M0101,该菌株对海藻酸钠具备很强的水解能力,通过对菌株 M0101的产酶条件进行优化,为后期该菌株在生产上的应用提供依据。
1. 材料与方法
1.1 材料与仪器
菌株 筛选自广西北海涠洲岛(109°08′00″E,21°01′00″N)的马尾藻;海藻酸钠培养基:(NH4)2SO4 5 g/L,K2HPO4 1 g/L,NaCl 15 g/L,MgSO4·7H2O 1 g/L,FeSO4·7H2O 0.01 g/L,海藻酸钠6 g/L,pH7.5;(NH4)2SO4 分析纯,广东光华科技股份有限公司;K2HPO4 分析纯,天津市大茂化学试剂厂;NaCl 分析纯,成都金山化学试剂有限公司;MgSO4·7H2O 分析纯,天津博迪化工股份有限公司;3, 5-二硝基水杨酸 化学纯,成都市科龙化工试剂厂;FeSO4·7H2O 分析纯,国药集团化学试剂有限公司;海藻酸钠 分析纯,阿拉丁;H2SO4 分析纯,廉江市廉化试剂有限公司。
HH-2数显恒温水浴锅 国华电器有限公司;TU-1901双光束紫外可见光分光光度计 北京普析通用仪器有限责任公司;TS-200B恒温摇床 上海天呈实验仪器制造有限公司;LHS-250SC恒温恒湿培养箱 上海天呈实验仪器制造有限公司;Tprofessional PCR仪 德国biometra;1-14K小型高速冷冻台式离心机 德国Sigma;Sherolock全自动细菌鉴定系统 美国MIDI(Microbial Identifica-tion)公司;HP6890气相色谱仪 美国安捷伦科技有限公司。
1.2 实验方法
1.2.1 海藻酸钠降解菌的分离
称取0.2 g马尾藻,置于1.5 mL的无菌离心管中,加入0.8 mL无菌水,用电动研磨棒研磨成匀浆液,取匀浆液进行10倍梯度稀释,取0.1 mL稀释液涂布于分离培养基上,将平板倒置于30 ℃的培养箱中培养2~5 d,挑取生长好、形态和颜色不同的菌落进行纯化培养,直至获得纯培养物。
1.2.2 产酶菌株的筛选
挑取单菌落点接于海藻酸钠培养基上,30 ℃培养3 d,以10%(w/v)氯代十六烷基吡啶覆盖平板染色(chlorhexadecyl pyridine,CPC)15 min,将显示水解圈的菌株进行褐藻胶裂解酶相对酶活力测定。
1.2.3 菌种鉴定
1.2.3.1 生理生化特征检测
在平板上挑取单菌落至液体分离培养基中,30 ℃,200 r/min培养 24 h,将菌液用无菌生理盐水稀释到适当浓度,取 100 μL 涂布于初筛培养基中,30 ℃下倒置培养3 d,观察菌株形态,形态鉴定参照《常见细菌系统鉴定手册》[14],将菌体送至中国科学院微生物研究所,由其完成生理生化特征检测和细胞脂肪酸组成测定。
1.2.3.2 细胞脂肪酸组成测定
使用Sherolock全自动细菌鉴定系统进行菌体脂肪酸成分分析,采用HP6890气相色谱仪进行色谱分析,配备分流/不分流进样口,氢火焰离子检测器(FID)及HP气相色谱化学工作站(HP CHEMSTATION ver A 5.05);色谱柱为UItra-2柱,长25 m,内径0.2 mm,滤膜厚度0.33 μm;炉温为二阶程序升温:起始温度170 ℃,每分钟5 ℃升至260 ℃,随后以40 ℃/min升至310 ℃,维持1.5 min;进样口温度250 ℃,载气为氢气,流速0.5 mL/min,分流进样模式,分流比100:1,进样量2 μL;检测器温度300 ℃,氢气流速30 mL/min,空气流速216 mL/min,补充气(氮气)流速30 mL/min。
1.2.3.3 分子生物学鉴定
菌株DNA的提取采用 Chelex-100法,用无菌牙签挑取少量菌体于1.5 mL的离心管中,加入50 μL 10% 的Chelex-100溶液,振荡混匀,于100 ℃加热10 min,冷却至室温后,12000 r/min离心10 min,上清液即可用于后续的DNA扩增。采用 16S rRNA 通用引物 27F(5' GAGTTTGATCCTGGCTCAG 3')、1492R(5'GGTTACCTTGTTACGACTT 3')进行 PCR扩增,扩增产物送至上海生工进行测序,测序结果在NCBI(National Center for Biotechnology Information)核苷酸数据库上进行 BLAST 分析,选取同源性高的 16S rRNA 序列,采用MEGA7.0(邻接法,Neighbor-Joining)构建菌株系统发育树。
1.2.4 菌株产酶活力测定
待测菌株用海藻酸钠液体培养基活化24 h作为种子培养液,按接种量3%(v/v)接种于装液量为50 mL的150 mL三角瓶中,置于摇床上,30 ℃,200 r/min发酵培养48 h后,取发酵液于4 ℃,12000 r/min离心5 min,上清作为粗酶液进行产酶活力测定。采用 3,5-二硝基水杨酸法(DNS)进行海藻酸钠裂解酶活力测定[15],方法略有改动。取100 μL发酵上清液和900 μL10 g/L 褐藻酸钠溶液(pH7.5的PBS配制)混合,在30 ℃下水浴30 min后,加入0.5 mL DNS缓冲液沸水浴5 min终止反应,对照组不水浴直接添加0.5 mL DNS 试剂后沸水浴 5 min,冷却后定容至5 mL,测定OD540 nm。
酶活力单位(U/mL)定义为: 在 30 ℃ 条件下,每分钟催化底物产生 1 μg 还原糖(以葡萄糖计)需要的酶量为一个酶活力单位。
相对酶活力测定: 将试验组的最高酶活力定义为 100%,将其他条件下的酶活力与最高酶活力的比值定义为相对酶活力。
1.2.5 产酶条件优化
以海藻酸钠培养基(如1.1所列)进行产酶条件的优化,在(NH4)2SO4 浓度为5 g/L,NaCl浓度为 15 g/L,初始pH为7.5,培养温度为30 ℃,转速为 200 r/min的条件下考察海藻酸钠浓度(18、20、22、24、26、28、32、40、50 g/L)对菌株产酶能力的影响;并依次考察氮源浓度(0.5、1、2、3、4、5、6、7、8 g/L);温度(25、30、35、40、45 ℃)、转速(100、150、200、250、300 r/min)、初始pH(5.5、6.0、6.5、7.0、7.5、8.0、8.5、9、9.5、10.0)及NaCl浓度(0、20、30、40、60、80、100、120、140、160 g/L)对菌株产酶能力的影响,以确定一个因素的最适条件为基准,依次对其余因素进行优化,确定菌株的产酶优化方案。
1.2.6 菌株生物量的测定
生物量通过OD600的大小来反映,以8 g/L的海藻酸钠液体培养基为空白对照,按一定倍数稀释菌液,应用TU-1901双光束紫外可见光分光光度计进行OD600值测定。
1.3 数据处理
运用软件Origin8.5对数据进行处理、统计和绘图。所有实验均重复三次。
2. 结果与分析
2.1 产酶菌株的筛选与鉴定
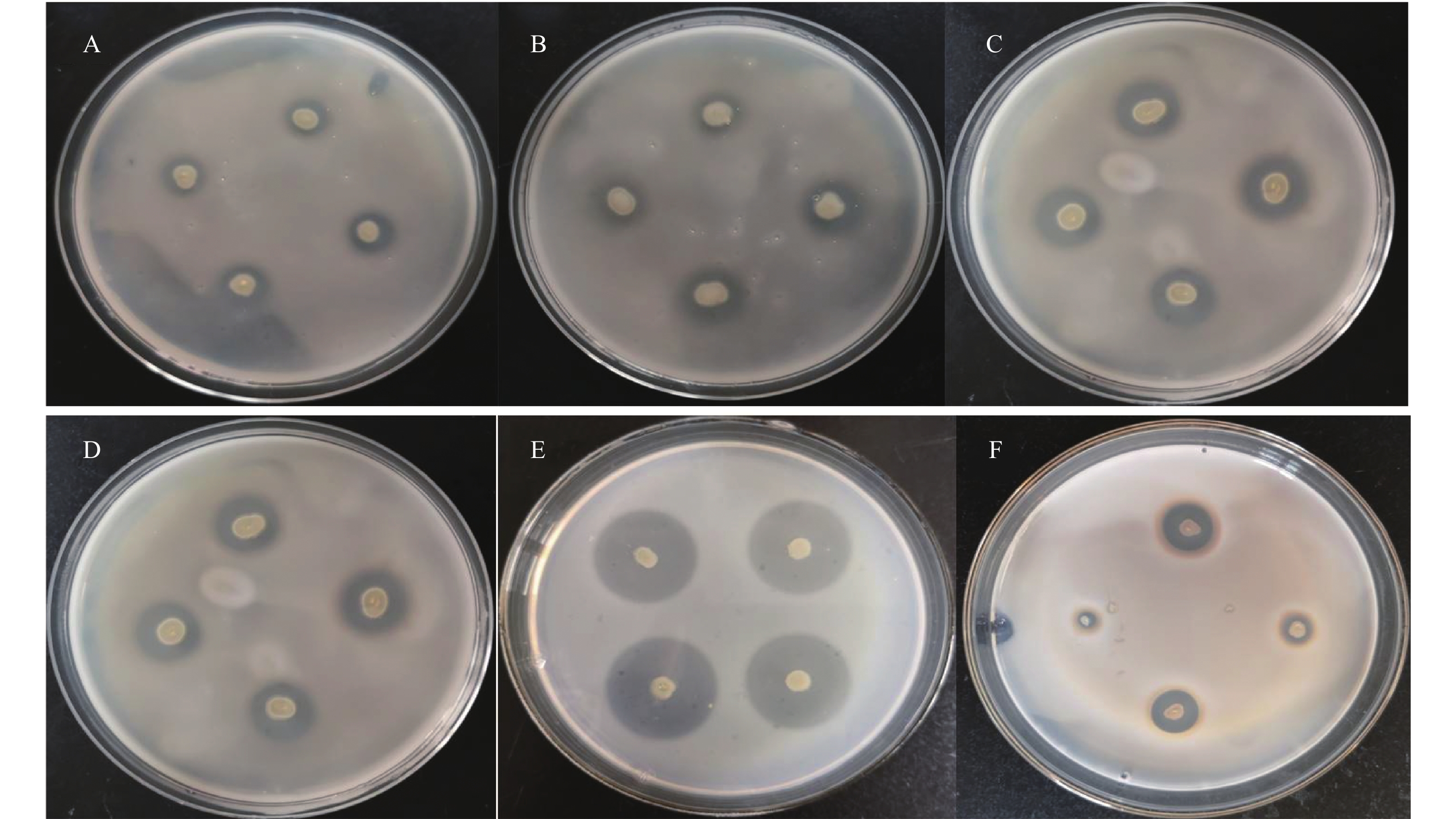
在以褐藻胶为唯一碳源的培养基上进行初筛,经10%(w/v)氯代十六烷基吡啶(Chlorhexadecyl pyridine,CPC)染色后,获得6株产酶菌株。将获得的6株产酶菌株进行液体发酵培养,测定其相对酶活力,发现菌株M0101的相对酶活力最高(结果见图1、图2),后续将进一步对M0101的产酶条件进行优化。
![]() 图 1 不同产海藻酸钠裂解酶菌株CPC(chlorhexadecyl pyridine)染色结果注:A~F:407-3, XW6102, 209, XW119, M0101, R202。Figure 1. CPC straining of different alginate lyases-producing bacterial strains
图 1 不同产海藻酸钠裂解酶菌株CPC(chlorhexadecyl pyridine)染色结果注:A~F:407-3, XW6102, 209, XW119, M0101, R202。Figure 1. CPC straining of different alginate lyases-producing bacterial strains![]() 图 2 不同产海藻酸钠裂解酶菌株相对酶活力Figure 2. Relative enzyme activity of different alginate lyases-producing bacterial strains
图 2 不同产海藻酸钠裂解酶菌株相对酶活力Figure 2. Relative enzyme activity of different alginate lyases-producing bacterial strains菌株M0101在筛选培养基上30 ℃培养3 d,菌落呈圆形,边缘整齐,淡黄色,表面光滑(图3)。该菌株在葡萄糖、甘露醇、甘露糖、甘油、乙酰氨基葡糖、麦芽糖中均可生长,其基因组DNA的G+C含量为41.1mol%,与弧菌属中相关菌种的生理生化特性的比较见表1。M0101中主要脂肪酸成分及含量为C12:0(9.38%), C14:0(4.24%), C16:0(15.61%), C18:1ω7c(14.69%), C18:1ω9c(1.95%), C14:0 3OH/C16:1 iso 1(9.09%), C16:1 w7c/C16:1 w6c(45.06%),与弧菌属相关菌种的脂肪酸成分比较如表2所示,经中国科学院微生物研究所检测鉴定,菌株M0101为弧菌Vibrio sp.。
表 1 菌株的生理生化特征Table 1. Physiological and biochemical characteristics of strains典型菌株 1 2 3 4 5 栖息环境 马尾藻 奶酪 排水池 潮汐滩地 猪圈垫层木屑 4 ℃生长情况 ND + + + − 42 ℃生长情况 + − − + + 12%(w/v)NaCl + − − + + 碳源利用 D-葡萄糖 + + + + + D-甘露糖 + + + + + D-半乳糖 − ND − + + 丙三醇 + ND − + + D- 甘露醇 + + − + + N-乙酰-D-葡糖胺 + + − + + D-麦芽糖 + + + + + 蔗糖 − ND + − + G+C 含量(mol%) 41.4 41.8 43.2 ND 43.0 注:1. Vibrio gangliei M0101; 2. Vibrio casei WS 4539T[16]; 3. Vibrio rumoiensis S-1T / Vibrio rumoiensis FERM P-14531T[17-18]; 4. Vibrio litoralis MANO22DT[17]; 5. Vibrio gangliei SZDIS-1T[19]; +:阳性反应;−:阴性反应;ND:未确定或未报道。 表 2 不同菌株脂肪酸组成(%)Table 2. Fatty acids composition of different strains (%)脂肪酸 1 2 3 4 5 C12:0 9.38 3.8 4.0 3.8 − C14:0 4.24 1.5 1.8 4.0 − C16:0 15.61 22.0 23.9 23.2 − C18:1ω7c 14.69 15.6 12 14.0 − C18:1ω9c 1.95 − − − − C14:0 3OH/C16:1 iso 1 9.09 3.2 7.7 7.0 − C16:1 w7c/C16:1 w6c 45.06 41.0 − 38.62 − 注:1. Vibrio gangliei M0101; 2. Vibrio casei WS 4539T[16]; 3. Vibrio rumoiensis S-1T / Vibrio rumoiensis FERM P-14531T[17-18]; 4. Vibrio litoralis MANO22DT[17]; 5, Vibrio gangliei SZDIS-1T[19];-:未确定或未报道。 对菌株的16S rDNA进行PCR扩增,产物经测序后,经同源性比对发现菌株M0101与Vibrio gangliei SZDIS-1序列的相似性达100%,该菌株与其近源菌株16S rDNA序列构建的系统发育树见图4,结合形态与生理生化特征分析,该菌株与弧菌Vibrio gangliei SZDIS-1相似性很高,将其命名为Vibrio gangliei M0101。
![]() 图 4 菌株M0101的16S rDNA序列系统进化树Figure 4. The phylogenetic tree of the 16S rRNA sequence of strain M0101
图 4 菌株M0101的16S rDNA序列系统进化树Figure 4. The phylogenetic tree of the 16S rRNA sequence of strain M01012.2 产酶条件优化
2.2.1 碳源、氮源浓度优化
培养基成分中海藻酸钠(C源)、硫酸铵(N源)对菌株M0101生长和相对酶活力影响的结果如图5所示。在18~50 g/L海藻酸钠浓度范围内,随着海藻酸钠浓度的升高,菌株的海藻酸钠裂解酶的相对酶活力和生物量呈上升的趋势(图5A)。尽管海藻酸钠添加量为50 g/L的培养基在接入M0101种子液前呈现极为浓稠的胶状,但接种发酵20 h后,浓稠胶状的培养基被水解成有较大流动性的溶液状态,细菌在培养基中分布均匀,可直接稀释一定倍数测定OD600反映其生物量。结果表明菌株M0101具备水解高浓度海藻酸钠的能力,其生物量和相对酶活力均在实验的浓度范围内达到最大值(图5A)。在更高的海藻酸钠浓度条件下,菌株M0101可能具有更高的水解酶活力,这有待进一步作深入研究,本文没有继续探讨其水解海藻酸钠的能力极限与发酵时间的关系,故选择海藻酸钠添加量为50 g/L为基准作后续优化。
![]() 图 5 不同碳源、氮源浓度对菌株M0101产酶和生长的影响Figure 5. The effect of different concentration of carbon source and nitrogen source on enzyme production and growth of strain M0101
图 5 不同碳源、氮源浓度对菌株M0101产酶和生长的影响Figure 5. The effect of different concentration of carbon source and nitrogen source on enzyme production and growth of strain M0101在0.5~8 g/L的浓度范围内,(NH4)2SO4对M0101生物量和相对酶活力的影响见图5B。当(NH4)2SO4质量浓度在0.5~2 g/L时,随着氮源浓度增加,菌株生物量和相对酶活力均随之增加,当(NH4)2SO4质量浓度为2 g/L时,菌株的相对酶活力达到最大值,之后随着氮源浓度的继续增加,生物量处于稳定的状态,而相对酶活力有所波动,综合考虑生物量、相对酶活力因素及用料成本,选择(NH4)2SO4的添加量为2 g/L,以此为基准作后续优化。
2.2.2 培养条件优化
不同温度、转速和初始pH发酵条件对菌株M0101产酶和生长影响结果如图6所示。随着发酵温度升高,菌株的生长和相对酶活力增加,相对酶活30 ℃时达到最大值,35 ℃时相对酶活力持平,随后随着温度增加而逐步下降。生物量在35 ℃后下降明显,相对酶活力受温度影响相对较低。当45 ℃时,菌株仍能生长,但生物量只有高峰值的三分之一,相对酶活力达到高峰值的80%左右(图6A);故本研究以较温和的温度30 ℃为基准作后续优化。
![]() 图 6 不同培养条件对菌株M0101产酶和生长的影响Figure 6. The effect of different culture conditions on enzyme production and growth of strain M0101
图 6 不同培养条件对菌株M0101产酶和生长的影响Figure 6. The effect of different culture conditions on enzyme production and growth of strain M0101转速对菌株相对酶活力影响不大,但对菌株的生长具有一定的影响(图6B),随着转速的增加,相对酶活力呈较平稳状态,而菌株的生长呈上升的趋势,转速低于200 r/min时,菌株的生长受抑制,可能与转速影响培养基中的容氧量相关。转速在200~300 r/min的范围内,相对酶活力有所波动,但转速分别为200 r/min和300 r/min时,相对酶活力没有显著性差异(P=0.065>0.05),故后续实验选择的转速为200 r/min,以此为基准作进一步优化。
初始pH对菌株产酶和生长影响结果如图6C所示,在pH5.5~9.5范围内,其相对酶活力和菌株生长随pH升高而呈上升趋势,pH为9.5时,相对酶活力达到最大值,且与pH为9.0时比较,相对酶活力相差不大,没有显著性差异(P=0.605>0.05);当pH大于9.5时,菌株的相对酶活力和生长能力急剧下降。综合考虑生物量、相对酶活力因素及未来可能的实际应用,后续实验选择的最适宜的酸碱度为pH9.0,以此为基准进一步优化NaCl浓度。
2.2.3 NaCl浓度优化
菌株M0101分离自腐烂的马尾藻,其生长和产酶需要一定的盐度,在培养基中添加不同浓度的NaCl对菌株M0101的生长和产酶影响很大。由图7可知,M0101的生长和产酶能力与NaCl的浓度密切相关,当NaCl浓度为100 g/L时,菌株的生物量和相对酶活力均达到最高值,尤其是生物量明显高于其他条件,说明其能适应高浓度NaCl环境,与其同源性很高的菌株Vibrio gangliei SZDIS-1能在NaCl浓度高达14%(140 g/L)的环境中生长[19],表明两者同为耐盐菌;M0101具有耐盐性推测与菌株生长的海洋环境相关;当NaCl浓度继续增加,菌株的生长和相对酶活力急剧下降,NaCl浓度大于140 g/L时,不利于菌株的生长和产酶。综上所述,选择NaCl的浓度为100 g/L。
![]() 图 7 不同NaCl浓度对菌株M0101产酶和生长的影响Figure 7. The effect of different concentration of NaCl on enzyme production and growth of strain M0101
图 7 不同NaCl浓度对菌株M0101产酶和生长的影响Figure 7. The effect of different concentration of NaCl on enzyme production and growth of strain M01012.2.4 优化前后菌株M0101生长和产酶能力
根据上文优化结果,产酶优化方案确定为:海藻酸钠50 g/L,(NH4)2SO4 2 g/L,K2HPO4 1 g/L,NaCl 100 g/L,MgSO4·7H2O 1 g/L,FeSO4·7H2O 0.01 g/L,初始pH9.0,30 ℃,200 r/min。菌株M0101发酵条件优化前后的生长及产酶能力如图8所示,优化前,菌株M0101发酵过程中延滞期较短,24 h以前已达到稳定期(见图8A),酶活力也处于平稳状态(图8B),酶活力较低。优化后,菌株M0101对海藻酸钠有较强的利用能力,培养基中碳源浓度高,粘稠,菌株发酵过程中延滞期延后,24 h以前处于延滞期,经24 h发酵后,浓稠状态的培养基(含50 g/L海藻酸钠的培养基呈浓稠状态)才被水解为流动的溶液状态,菌体在培养基中均匀分布,此时可利用紫外可见光分光光度计测定600 nm处的吸光度值,以此反映发酵菌株的生物量。32 h后菌株进入对数生长期(图8A),是海藻酸钠裂解酶迅速积累期,发酵32 h酶活力达到最大值221.90 U/mL(图8B),48 h后菌株的生长到达稳定期,此时酶活力已开始逐渐降低。相较于发酵条件优化前最大酶活36.32 U/mL,优化后酶活力是优化前的6.11倍。
![]() 图 8 菌株M010发酵条件优化前后生长及产酶曲线Figure 8. Growth and enzyme production curve of strain M0101 before and after optimization of fermentation conditions
图 8 菌株M010发酵条件优化前后生长及产酶曲线Figure 8. Growth and enzyme production curve of strain M0101 before and after optimization of fermentation conditions3. 讨论与结论
褐藻胶裂解酶酶活的高低是微生物吸收利用褐藻胶的关键,其作为一种用途广泛的工具酶,在制备低分子质量的褐藻胶寡糖时,因反应条件温和、专一性强、得率高而逐渐成为制备褐藻胶寡糖的主要方式[20-21]。近年来,从细菌中分离出了许多褐藻胶裂解酶,产酶菌属主要包括Vibrio[22-23]、Flavobacterium[24-25]、Pseudomonas[26-27]、Microbulbifer[28-29]等。本研究中产酶菌株是从北海涠洲岛采集的马尾藻中分离出来的一株产褐藻胶裂解酶菌株,根据菌株形态、生理生化特征和16S rRNA基因序列分析,鉴定为弧菌属Vibrio,命名为Vibrio gangliei M0101。弧菌属内不同菌株的褐藻胶裂解酶活力存在较大差异,李云涛等[30]对Vibrio sp.SS-1通过响应面法优化后,酶活力最大可达191.8 U/mL,傅晓妍等[31]筛选到的弧菌Vibrio sp.QY102优化培养120 h后,产酶达10.2 U/mL,与本文中Vibrio gangliei M0101发酵优化后,产酶活性达221.90 U/mL比较,均存在差异,推测与褐藻胶裂解酶相关基因的差异有关。
培养基的成分在微生物发酵过程中对细菌的产酶活力具有决定性作用,合适的碳、氮、金属离子能够促进菌株的生长和产酶[32]。菌株M0101能够高效利用高浓度海藻酸钠,随着海藻酸钠浓度的升高,菌株的褐藻胶裂解酶活力和生物量呈上升的趋势,具备高效水解高浓度海藻酸钠(50 g/L)的能力。NaCl通常起着维持菌体内外渗透压、维持细胞内外电势差、参与物质运输的作用,在以褐藻胶为碳源的微生物培养基中,还起着运送褐藻胶大分子进入细胞膜的作用,当培养基中NaCl质量浓度高达100 g/L时,菌株
M0101仍能正常生长,符合海洋细菌嗜盐的特点,与其他同类型产海藻酸钠裂解酶的嗜盐菌株相比,该菌株的盐耐受性较强[33];同时,NaCl还能影响海藻酸钠裂解酶的活性,Xu等[34]报道的PL7家族海藻酸钠裂解酶AlyC3是一种新型的耐低温、盐活化酶,NaCl浓度为0.5 mol/L(29.25 g/L)时酶活力最强,且浓度为3 mol/L(175.5 g/L)时酶活力仍高于没有NaCl存在时。Huang等[35]发现 V. weizhoudaoensis M0101(Vibrio gangliei M0101)中的algM4基因编码的AlgM4酶是一种新型的盐活化的双功能酶,经克隆、表达、纯化该双功能酶,发现其最适的NaCl浓度为1 mol/L(58.5 g/L),NaCl不仅改变了该酶的二级结构元素的组成,同时也提高了该酶的热稳定性;该酶的最适NaCl浓度与其菌体产酶的最适浓度(100 g/L)差异大,对M0101基因组数据进行分析,发现M0101具有5种海藻酸钠裂解酶编码基因,推测这种差异与M0101能同时表达多种褐藻酸钠裂解酶有关。 对培养基成分及培养条件优化后,确定菌株M0101最佳产酶条件为海藻酸钠50 g/L,(NH4)2SO4 2 g/L,K2HPO4 1 g/L,NaCl 100 g/L,MgSO4·7H2O 1 g/L,FeSO4·7H2O 0.01 g/L,初始pH9.0,培养温度30 ℃,摇床转速200 r/min,在此条件下可获得最大酶活为221.90 U/mL,是优化前的6.11倍。这为高活性褐藻胶裂解酶工业制备和褐藻胶寡糖生产提供了理论依据。
-
![]()
图 1 不同产海藻酸钠裂解酶菌株CPC(chlorhexadecyl pyridine)染色结果
注:A~F:407-3, XW6102, 209, XW119, M0101, R202。
Figure 1. CPC straining of different alginate lyases-producing bacterial strains
![]()
图 2 不同产海藻酸钠裂解酶菌株相对酶活力
Figure 2. Relative enzyme activity of different alginate lyases-producing bacterial strains
![]()
图 4 菌株M0101的16S rDNA序列系统进化树
Figure 4. The phylogenetic tree of the 16S rRNA sequence of strain M0101
![]()
图 5 不同碳源、氮源浓度对菌株M0101产酶和生长的影响
Figure 5. The effect of different concentration of carbon source and nitrogen source on enzyme production and growth of strain M0101
![]()
图 6 不同培养条件对菌株M0101产酶和生长的影响
Figure 6. The effect of different culture conditions on enzyme production and growth of strain M0101
![]()
图 7 不同NaCl浓度对菌株M0101产酶和生长的影响
Figure 7. The effect of different concentration of NaCl on enzyme production and growth of strain M0101
![]()
图 8 菌株M010发酵条件优化前后生长及产酶曲线
Figure 8. Growth and enzyme production curve of strain M0101 before and after optimization of fermentation conditions
表 1 菌株的生理生化特征
Table 1 Physiological and biochemical characteristics of strains
典型菌株 1 2 3 4 5 栖息环境 马尾藻 奶酪 排水池 潮汐滩地 猪圈垫层木屑 4 ℃生长情况 ND + + + − 42 ℃生长情况 + − − + + 12%(w/v)NaCl + − − + + 碳源利用 D-葡萄糖 + + + + + D-甘露糖 + + + + + D-半乳糖 − ND − + + 丙三醇 + ND − + + D- 甘露醇 + + − + + N-乙酰-D-葡糖胺 + + − + + D-麦芽糖 + + + + + 蔗糖 − ND + − + G+C 含量(mol%) 41.4 41.8 43.2 ND 43.0 注:1. Vibrio gangliei M0101; 2. Vibrio casei WS 4539T[16]; 3. Vibrio rumoiensis S-1T / Vibrio rumoiensis FERM P-14531T[17-18]; 4. Vibrio litoralis MANO22DT[17]; 5. Vibrio gangliei SZDIS-1T[19]; +:阳性反应;−:阴性反应;ND:未确定或未报道。  下载: 导出CSV
下载: 导出CSV
表 2 不同菌株脂肪酸组成(%)
Table 2 Fatty acids composition of different strains (%)
脂肪酸 1 2 3 4 5 C12:0 9.38 3.8 4.0 3.8 − C14:0 4.24 1.5 1.8 4.0 − C16:0 15.61 22.0 23.9 23.2 − C18:1ω7c 14.69 15.6 12 14.0 − C18:1ω9c 1.95 − − − − C14:0 3OH/C16:1 iso 1 9.09 3.2 7.7 7.0 − C16:1 w7c/C16:1 w6c 45.06 41.0 − 38.62 − 注:1. Vibrio gangliei M0101; 2. Vibrio casei WS 4539T[16]; 3. Vibrio rumoiensis S-1T / Vibrio rumoiensis FERM P-14531T[17-18]; 4. Vibrio litoralis MANO22DT[17]; 5, Vibrio gangliei SZDIS-1T[19];-:未确定或未报道。
下载: 导出CSV
-
[1] Zhu B W, Ni F, Ning L M, et al. Cloning and characterization of a new pH-stable alginate lyase with high salt tolerance from marine Vibrio sp. NJ-04[J]. International Journal of Biological Macromolecules,2018,115:1063−1070. doi: 10.1016/j.ijbiomac.2018.04.108
[2] Chen J Y, Hu Y, Zhang L R, et al. Alginate oligosaccharide DP5 exhibits antitumor effects in osteosarcoma patients following surgery[J]. Front Pharmacol,2017,8:623. doi: 10.3389/fphar.2017.00623
[3] Falkeborg M, Cheong L Z, Gianfico C, et al. Alginate oligosaccharides: Enzymatic preparation and antioxidant property evaluation[J]. Food Chem,2014,164:185−194. doi: 10.1016/j.foodchem.2014.05.053
[4] Khan S, Tøndervik A, Sletta H, et al. Overcoming drug resistance with alginate oligosaccharides able to potentiate the action of selected antibiotics[J]. Antimicrob Agents Chemother,2012,56(10):5134−5141. doi: 10.1128/AAC.00525-12
[5] Liu H, Zhang Y H, Yin H, et al. Alginate oligosaccharides enhanced Triticum aestivum L. tolerance to drought stress[J]. Plant Physiol Biochem,2013,62:33−40. doi: 10.1016/j.plaphy.2012.10.012
[6] 邰宏博, 唐丽薇, 陈带娣, 等. 褐藻胶寡糖制备的研究进展[J]. 生命科学研究,2015,19(1):75−79. [Tai H B, Tang L W, Chen D D, et al. Progresses on preparation of alginate oligosaccharide[J]. Life Science Research,2015,19(1):75−79. [7] Osawa T, Matsubara Y, Muramatsu T, et al. Crystal structure of the alginate (poly alpha-l-guluronate) lyase from Corynebacterium sp. at 1.2 A resolution[J]. Journal of Molecular Biology,2005,345(5):1111−1118. doi: 10.1016/j.jmb.2004.10.081
[8] Tomoo S, Takahashi H, Ezura Y, et al. Cloning, sequence analysis and expression of Pseudoalteromonas elyakoTii IAM 14594 gene (alyPEEC) encoding the extracellular alginate lyase[J]. Carbohydrate Research,2001,335(1):11−21. doi: 10.1016/S0008-6215(01)00198-7
[9] 刘海超, 张健孙, 王共明, 等. 褐藻胶的降解方法及其产物生物活性研究进展[J]. 食品工业科技,2020,41(13):350−363. [Liu H C, Zhang J S, Wang G M, et al. Advances in degradation methods of alginate and bioactivity of its degradation products[J]. Science and Technology of Food Industry,2020,41(13):350−363. [10] 刘岩, 江晓路, 管华诗. 褐藻胶裂解酶研究进展[J]. 中国水产科学,2000,7(4):99−103. [Liu Y, Jiang X L, Guan H S. Advance in alginate lyases[J]. Journal of Fishery Sciences of China,2000,7(4):99−103. doi: 10.3321/j.issn:1005-8737.2000.04.023 [11] Cheng W, Yan X, Xiao J, et al. Isolation, identification, and whole genome sequence analysis of the alginate-degrading bacterium Cobetia sp. cqz5-12[J]. Sci Rep,2020,10(1):10920. doi: 10.1038/s41598-020-67921-7
[12] 赵婉琳, 叶静, 张娜, 等. 褐藻胶降解菌的筛选、鉴定及产酶条件优化[J]. 微生物学报,2019,59(1):169−180. [Zhao W L, Ye J, Zhang N, et al. Screening, identification and fermentation optimization of an alginate-degrading strain[J]. Acta Microbiologica Sinica,2019,59(1):169−180. [13] 郑志国, 黄惠琴, 朱军, 等. 响应面法优化类芽胞杆菌HB172198产褐藻胶裂解酶发酵培养基[J]. 基因组学与应用生物学,2020,32(1):151−158. [Zheng Z G, Huang H Q, Zhu J, et al. Optimization of culture medium for alginate lyase produced by Paenibacillus HB172198 using response surface methodology[J]. Genomics and Applied Biology,2020,32(1):151−158. [14] 东秀珠, 蔡妙英.常见细菌系统鉴定手册[M]. 北京: 科学出版社, 2001. Dong X Z, Cai M Y. Manual for system identification of common bacteria [M]. Beijing: Science Press, 2001.
[15] Zhu Y, Wu L, Chen Y, et al. Characterization of an extracellular biofunctional alginate lyase from marine Microbulbifer sp. ALW1 and antioxidant activity of enzymatic hydrolysates[J]. Microbiol Res,2016,182:49−58. doi: 10.1016/j.micres.2015.09.004
[16] Bleicher A, Neuhaus K, Scherer S. Vibrio casei sp. nov., isolated from the surfaces of two French red smear soft cheeses[J]. Int J Syst Evol Microbiol,2010,60(Pt 8):1745−1749.
[17] Nam Y D, Chang H W, Park J R, et al. Vibrio litoralis sp. nov., isolated from a Yellow Sea tidal flat in Korea[J]. Int J Syst Evol Microbiol,2007,57(Pt 3):562−565.
[18] Yumoto I I, Iwata H, Sawabe T, et al. Characterization of a facultatively psychrophilic bacterium, Vibrio rumoiensis sp. nov., that exhibits high catalase activity[J]. Appl Environ Microbiol,1999,65(1):67−72. doi: 10.1128/AEM.65.1.67-72.1999
[19] Meng Y C, Liu H C, Zhou Y G, et al. Vibrio gangliei sp. nov., a novel member of Vibrionaceae isolated from sawdust in a pigpen[J]. International Journal of Systemation and Evolutionary Microbiology,2018,68(6):1969−1974. doi: 10.1099/ijsem.0.002779
[20] Liu J, Yang S Q, Li X T, et al. Alginate oligosaccharides: Production, biological activities, and potential applications[J]. Comprehensive Reviews in Food Science and Food Safety,2019,18(6):1859−1881. doi: 10.1111/1541-4337.12494
[21] 陈淑琼, 李晓月, 吴晨烁, 等. 酶解制备褐藻寡糖工艺优化及活性研究[J]. 食品研究与开发,2019,40(13):51−56. [Chen S Q, Li X Y, Wu C S, et al. Optimization of process conditions and activity for enzymatic hydrolysis of sodium alginate[J]. Food Research and Development,2019,40(13):51−56. [22] Li S S, Wang L N, Hao J H, et al. Purification and characterization of a new alginate lyase from marine bacterium Vibrio sp. SY08[J]. Mar Drugs,2016,15(1):1−11. doi: 10.3390/md15010001
[23] 李云涛, 张齐, 汪立平, 等. 海洋弧菌中褐藻胶裂解酶Alg的克隆表达及酶学性质[J]. 山东农业大学学报(自然科学版),2018,49(4):659−666. [Li Y T, Zhang Q, Wang L P, et al. Cloning expression and characters of alginate lyase gene alg from marine Vibrio[J]. Journal of Shandong Agricultural University (Natural Science Edition),2018,49(4):659−666. [24] Lavín P, Atala C, Gallardo-Cerda J, et al. Isolation and characterization of an Antarctic Flavobacterium strain with agarase and alginate lyase activities[J]. Polish Polar Research,2016,37(3):403−419. doi: 10.1515/popore-2016-0021
[25] Zhou H X, Xu S S, Yin X J, et al. Characterization of a new bifunctional and cold-adapted polysaccharide lyase (PL) family 7 alginate lyase from Flavobacterium sp.[J]. Mar Drugs,2020,18(8):388−406. doi: 10.3390/md18080388
[26] Li L Y, Jiang X L, Guan H S, et al. Three alginate lyases from marine bacterium Pseudomonas fluorescens HZJ216: Purification and characterization[J]. Appl Biochem Biotechnol,2011,164(3):305−317. doi: 10.1007/s12010-010-9136-4
[27] Lundqvist L C E, Jam M, Barbeyron T, et al. Substrate specificity of the recombinant alginate lyase from the marine bacteria Pseudomonas alginovora[J]. Carbohydrate Research,2012,352:44−50. doi: 10.1016/j.carres.2012.02.014
[28] Yang J, Cui D D, Chen D W, et al. Purification and characterization of a novel endolytic alginate lyase from Microbulbifer sp. SH-1 and its agricultural application[J]. Mar Drugs,2020,18(4):1−19.
[29] 杨锦, 沈宏. 海藻酸裂解酶高产菌株 Microbulbifer sp. SH-1的分离、鉴定及其产酶条件优化[J]. 微生物学报,2019,360(4):85. [Yang J, Shen H. Isolation and identification of an alginate lyase-producing strain Microbulbifer sp. SH-1 and optimization of its enzyme-producing conditions[J]. Acta Microbiologica Sinica,2019,360(4):85. [30] 李云涛, 汪立平, 张孟, 等. 鲍鱼来源褐藻胶裂解酶菌株的筛选及发酵条件优化[J]. 大连海洋大学学报,2017,32(5):574−583. [Li Y T, Wang L P, Zhang M, et al. Screening of alginate lyase-producing microorganisms and optimization of fermentation condition in disc abalone[J]. Journal of Dalian University,2017,32(5):574−583. [31] 傅晓妍, 李京宝, 韩峰, 等. 褐藻胶裂解酶产生菌Vibro sp. QY102的发酵条件优化[J]. 中国海洋大学学报(自然科学版),2007,37(3):432−436. [Fu X X, Li J B, Han F, et al. Studies on Vibro sp. QY102 fermentation processes for alginate lyase production[J]. Periodical of Ocean University of China,2007,37(3):432−436. [32] 侯士昌, 温少红, 唐志红, 等. 一株高效褐藻酸降解菌的筛选、鉴定及其发酵条件的优化[J]. 海洋科学,2014,38(7):20−26. [Hou S C, Wen S H, Tang Z H, et al. The screening, identification of alginate degrading bacteria and optimization of fermentation conditions[J]. Marine Sciences,2014,38(7):20−26. doi: 10.11759/hykx20130722001 [33] 许超, 熊亚茹, 卢明倩, 等. 一株具有褐藻胶降解能力的海洋细菌的筛选鉴定及其多糖利用能力研究[J]. 生物技术通报,2017,33(4):198−204. [Xu C, Xiong Y R, Lu M Q, et al. Screening and identification of marine alginate-degrading bacterium and the utilization capacity of polysaccharide[J]. Biotechnology Bulletin,2017,33(4):198−204. [34] Xu F, Chen X L, Sun X H, et al. Structural and molecular basis for the substrate positioning mechanism of a new PL7 subfamily alginate lyase from the arctic[J]. J Biol Chem,2020,295(48):16380−16392. doi: 10.1074/jbc.RA120.015106
[35] Huang G Y, Wang Q Z, Lu M Q, et al. AlgM4: A new salt-activated alginate lyase of the PL7 family with endolytic activity[J]. Marine Drugs,2018,16(4):1−13.
-
期刊类型引用(5)
1. 高含,刘锐,姜铖,王飞飞. 超高效液相色谱—串联质谱法测定植物油中甲胺磷残留量的不确定度评定. 食品与机械. 2024(10): 62-67 .  百度学术
百度学术
2. 董孝元,周玉,余义,刘莎,汪明迪,孙莉,唐艳荣. LC-ESI-MS/MS测定白酒中氨基甲酸乙酯的方法研究. 酿酒科技. 2023(11): 105-111 . 百度学术
3. 聂叶,焦富,罗汝叶,李巧玉,牟明月. 酱香型白酒酒醅中乙醇测定方法的不确定度评定. 酿酒科技. 2022(08): 125-130 . 百度学术
4. 陈同强,梁锋,吴海智,向俊,宋阳,荆辉华,袁列江,李灿,李凯龙. 气相色谱-质谱法测定发酵乳中氨基甲酸乙酯含量. 乳业科学与技术. 2022(06): 18-21 . 百度学术
5. 李正刚,马彧,王丹彧,李本淳,赵艳. HPLC法测定制何首乌饮片中2, 3, 5, 4'-四羟基二苯乙烯-2-O-β-D-葡萄糖苷含量的不确定度评估. 中国医药导刊. 2021(07): 522-525 . 百度学术
其他类型引用(0)
 下载:
下载:















计量
- 文章访问数: 226
- HTML全文浏览量: 161
- PDF下载量: 39
- 被引次数: 5



